Virus

Un virus (do latín virus 'toxina' ou 'veleno') é un axente infeccioso microscópico non celular que só pode replicarse penetrando no interior dunha célula. Os virus afectan todo tipo de organismos, desde animais e plantas a bacterias e arqueas[1]. Unha partícula de virus completa e infectiva fóra (xeralmente) da célula denomínase virión[2]. O descubrimento dos virus empezou cando Dmitri Ivanovsky en 1892 atopou un axente infeccioso das plantas do tabaco tan pequeno que atravesaba os poros dos filtros que retiñan bacterias, o que levou ao descubrimento do virus do mosaico do tabaco por Martinus Beijerinck en 1898.[3] Desde entón, describíronse en detalle arredor duns 5.000 virus[4], aínda que se estima que existen millóns de tipos diferentes de virus[5]. Os virus atópanse en case calquera ecosistema da Terra e estas diminutas estruturas son o tipo de entidade biolóxica numericamente máis abundante[6][7]. O estudo dos virus coñécese como viroloxía, unha subespecialidade da microbioloxía.
Os virus non teñen estrutura celular, senón que simplemente están formados por unha cápside (ou cápsida) de proteínas que envolve o seu ácido nucleico, que pode ser ADN ou ARN. Esta estrutura pode, á súa vez, estar rodeada pola envoltura vírica, unha capa lipídica con diferentes proteínas dependendo do virus. O ciclo vital dun virus sempre necesita da maquinaria metabólica dunha célula para poder replicar o seu material xenético e cápside, producindo moitas copias do virus orixinal. No devandito proceso reside a capacidade destrutora dos virus, xa que poden prexudicar a célula ata destruíla. Poden infectar tanto células eucarióticas como procarióticas (neste último caso denomínanse bacteriófagos, ou simplemente fagos).
As partículas víricas, chamadas virións poden ser de forma helicoidal ou icosaédrica ou ter formas máis complexas. O tamaño medio dun virus é cen veces menor có das bacterias, polo que miden poucas decenas de nanómetros e só se poden ver con microscopio electrónico, aínda que hai uns poucos exemplos de virus que comparativamente son excepcionalmente grandes.
A orixe dos virus na historia evolutiva da vida non está clara: algúns puideron ter evolucionado a partir de plásmidos, que son pedazos de ADN que poden pasar dunha célula a outra, mentres que outros puideron evolucionar regresivamente a partir de bacterias. Na evolución os virus son un medio importante para que teña lugar a transferencia horizontal de xenes entre especies, que incrementa a diversidade xenética.[8] Algúns consideran que os virus son formas de vida, porque levan material xenético, reprodúcense e evolucionan por selección natural. Con todo, carecen de características fundamentais como a estrutura celular, que son xeralmente consideradas necesarias para falar de vida. Como posúen algunhas pero non todas as calidades da vida, os virus foron descritos como "organismos no límite da vida".[9]
Os virus transmítense de diversos xeitos. Os virus de plantas xeralmente transmítense de planta a planta por medio de insectos que se alimentan dos zumes das plantas, como os áfidos; os virus de animais poden ser transportados por insectos hematófagos. Estes organismos que transmiten enfermidades inoculando o axente infeccioso denomínanse vectores. Outros virus como o da gripe espállanse polos espirros e toses. Os norovirus e rotavirus, que son unha causa común de gastroenterite viral, transmítense pola ruta fecal-oral e pasan por contacto de persoa a persoa, entrando no corpo coa comida e auga. Virus como o VIH son exemplo de virus transmitidos por vía sexual e sanguínea. A gama de células hóspede que un virus pode infectar é o seu "rango de hóspedes". Este rango pode ser estreito ou máis amplo, se o virus pode infectar moitas especies.[10]
As infeccións virais en animais causan unha resposta inmunitaria que xeralmente elimina o virus infeccioso. As respostas inmunes poden tamén producirse pola administración de vacinas, as cales confiren unha inmunidade adquirida artificialmente contra unha determinada especie ou cepa de virus. Porén, algúns virus, entre os que están os que causan a SIDA e a hepatite viral, evaden estas respostas inmunes e dan lugar a infeccións crónicas. Os antibióticos non teñen efecto sobre os virus, pero existen diversos medicamentos antivirais.
Etimoloxía
[editar | editar a fonte]A palabra virus vén do latín virus, que significa veleno, toxina. A palabra en plural tamén se di virus. De virus derivan os adxectivos vírico, viral, virulento e o nome virión. En principio, a palabra virus tiña un significado moi xeral e aplicábase a calquera axente infeccioso que causase enfermidades (incluídas as enfermidades bacterianas) e rexístrase por primeira vez no século XVIII[11], pero desde o final do século XIX, a palabra foi adquirindo o seu significado actual máis específico. Un virión é unha partícula vírica estable e infectiva que é liberada da célula e ten capacidade de infectar outras células do mesmo tipo.[2]
Historia
[editar | editar a fonte]
Ao longo do século XIX foise aceptando que moitas enfermidades se debían a axentes infecciosos, que se cría eran bacterias e chegaron a identificarse varios destes axentes bacterianos. Pero non sempre era fácil identificar o axente causante de certas doenzas infecciosas. Louis Pasteur non puido atopar cal era o axente causante da rabia e especulou sobre que podía ser un patóxeno demasiado pequeno para ser detectado utilizando un microscopio.[12] En 1884, o microbiólogo francés Charles Chamberland inventou un filtro que tiña poros máis pequenos ca unha bacteria. Facendo pasar unha solución que contiña bacterias a través dese filtro, as bacterias quedaban retidas nel e a solución filtrada obtida era estéril.[13] En 1892, o biólogo ruso Dmitri Ivanovsky utilizou estes filtros para estudar un extracto de plantas do tabaco afectadas pola enfermidade do mosaico (que hoxe sabemos é producida polo virus do mosaico do tabaco). Ditos extractos filtrados por Ivanovsky seguían sendo infectivos. Ivanovsky suxeriu que a infección podería estar causada por unha toxina producida pola bacteria, pero non desenvolveu a idea.[14] Naquel tempo pensábase que todos os axentes infecciosos podían quedar retidos nos filtros e cultivados nun medio nutritivo, o que formaba parte da teoría do xerme das enfermidades.[3] En 1898, o microbiólogo holandés Martinus Beijerinck repetiu os experimentos e quedou convencido de que a solución filtrada contiña unha nova forma de axente infeccioso moi pequeno.[15] Observou que dito axente se multiplicaba só en células que estaban dividíndose, pero como os seus experimentos non mostraron que estaba feito de partículas, chamouno contagium vivum fluidum (xerme vivo soluble) e reintroduciu a palabra virus.[14] Beijerinck sostiña que os virus eran de natureza líquida, unha teoría que máis tarde foi desacreditada por Wendell Stanley, que probou que eran particulados.[14] Nese mesmo ano Friedrich Loeffler e Paul Frosch fixeron pasar por un filtro similar o primeiro virus animal, o axente da febre aftosa (Aphthovirus).[16]
A comezos do século XX, o bacteriólogo inglés Frederick Twort descubriu un grupo de virus que infectaban bacterias, hoxe chamados bacteriófagos[17] (ou comunmente fagos), o microbiólogo francocanadense Félix d'Herelle describiu virus que, cando se engadían a colonias de bacterias en placa de ágar, producían áreas de bacterias mortas. Diluíu con precisión unha suspensión deses virus e descubriu que canto maior era a dilución (é dicir, menor era a concentración viral), en vez de mataren todas as bacterias os virus producían áreas discretas de bacterias mortas. Contando estas áreas e multiplicando polo factor de dilución podía calcular o número de virus na suspensión orixinal.[18] Os fagos consideráronse como un tratamento potencial de enfermidades como as febres tifoides e o cólera, pero esa esperanza foi esquecida coa chegada da penicilina. O estudo dos fagos proporcionou unha nova visión sobre a activación e inactivación de xenes e do modo de introducir xenes alleos dentro de bacterias.
No final do século XIX, os virus eran definidos en termos da súa infectividade, a súa capacidade de seren filtrados e os seus requirimentos de hóspedes vivos. Ata entón os virus cultiváranse só en plantas e animais. En 1906, Ross Granville Harrison inventou un método para cultivar tecidos en linfa, e, en 1913, E. Steinhardt, C. Israeli e R. A. Lambert utilizaron ese método para cultivar o virus vaccinia en fragmentos de tecido corneal de cobaia.[19] En 1928, H. B. Maitland e M. C. Maitland cultivaron o virus vaccinia en suspensións de riles de galiña esmiuzados. O seu método non se xeneralizou ata 1950, cando se cultivou o poliovirus para a produción de vacinas en grande escala.[20]
En 1931 conseguiuse outro grande avance cando o patólogo norteamericano Ernest William Goodpasture cultivou o virus da gripe e outros varios virus en ovos fertilizados de galiña.[21] En 1949, John Franklin Enders, Thomas Weller e Frederick Robbins cultivaron o virus da polio en células humanas embrionarias, que foi o primeiro virus que se cultivou sen usar un tecido animal sólido ou ovos. Este traballo permitiulle a Jonas Salk preparar unha vacina efectiva contra a polio.[22]
As primeiras imaxes de virus obtivéronse cando en 1931 os enxeñeiros alemáns Ernst Ruska e Max Knoll inventaron o microscopio electrónico .[23] En 1935, o bioquímico e virólogo norteamericano Wendell Meredith Stanley examinou o virus do mosaico do tabaco e comprobou que estaba feito principalmente de proteína.[24] Pouco tempo despois, viuse que este virus estaba formado por proteínas e ARN.[25] O virus do mosaico do tabaco foi o primeiro en ser cristalizado e puido así determinarse a súa estrutura en detalle. As primeiras imaxes de difracción de raios X do virus cristalizado obtivéronas Bernal e Fankuchen en 1941. Baseándose nas súas imaxes, Rosalind Franklin descubriu a estrutura completa deste virus en 1955.[26] Ese mesmo ano, Heinz Fraenkel-Conrat e Robley Williams demostraron que o ARN e a proteína da cuberta purificadas do virus do mosaico do tabaco poden autoensamblarse formando virus funcionais, o que suxería que este simple mecanismo era probablemente o modo polo que se orixinaban os virus nas células vivas.[27]
A segunda metade do século XX foi a idade dourada de descubrimento de virus e recoñecéronse máis de 2.000 novas especies de virus de animais, plantas e bacterias.[28] En 1957, descubríronse o arterivirus equino e o virus da diarrea bovina (un pestivirus). En 1963, Baruch Blumberg descubriu o virus da hepatite B,[29] e en 1965, Howard Temin describiu o primeiro retrovirus. A reversotranscriptase, que é o encima fundamental dos retrovirus, que utilizan para copiar o seu ARN a ADN, describiuse en 1970, independentemente por Howard Martin Temin e David Baltimore.[30] En 1983 o equipo de Luc Montagnier no Instituto Pasteur en Francia, illou por primeira vez o retrovirus que hoxe coñecemos como VIH.[31]
Orixe
[editar | editar a fonte]Os virus encóntranse en todos os lugares onde haxa seres vivos, e levan existindo probablemente desde que evolucionaron as primeiras células.[32] A orixe dos virus non está clara porque non forman fósiles, polo que houbo que usar técnicas moleculares para comparar o seu ADN ou ARN e investigar como xurdiron.[33] Porén, o material xenético viral pode integrarse ocasionalmente na liña xerminal do organismo hóspede e pasar así para a súa descendencia durante moitas xeracións, o que proporciona unha valiosa fonte de información aos paleovirólogos para que sigan o rastro de antigos virus que existiron hai millóns de anos. Existen tres hipóteses principais que intentan explicar a orixe dos virus:[34][35]
- Hipótese regresiva
- Os virus puideron ser inicialmente pequenas células que parasitaban células meirandes. Co paso do tempo, foron perdendo os xenes que non eran imprescindibles para ese parasitismo. As bacterias rickettsias e clamidias son células vivas que, igual cós virus, só poden reproducirse dentro doutras células e serven para apoiar esta hipótese, xa que a súa dependencia do parasitismo é probable que causase a perda dos xenes que lles permitían orixinalmente sobrevivir fóra da célula. Tamén se chama hipótese da dexeneración,[36][37] ou hipótese da redución.[38]
- Hipótese da orixe celular
- Algúns virus pode que evolucionasen a partir de fragmentos de ADN ou ARN que "escaparon" dos xenes dun organismo máis grande. O ADN que escapou puido proceder de plásmidos (anacos de ADN que poden moverse entre distintas células) ou transposóns (moléculas de ADN que se replican e se moven a unha posición diferente dentro dos xenes da célula).[39] Os transposóns, antes chamados "xenes saltadores", son exemplos de elementos xenéticos móbiles e poderían ser a orixe de moitos virus. Foron descubertos no millo por Barbara McClintock en 1950.[40] Esta hipótese chámase ás veces hipótese da lacazanería (vagrancy hypothesis),[36][41] ou hipótese do escape.[38]
- Hipótese da coevolución
- A hipótese da coevolución tamén se chama hipótese dos virus primeiro.[38] Propón que os virus puideron ter evolucionado a partir de moléculas complexas de proteínas e ácidos nucleicos ao mesmo tempo que apareceron as primeiras células na Terra e terían sido dependentes da vida celular durante miles de millóns de anos. Os viroides son moléculas de ARN que non se clasifican como virus porque carecen de cuberta proteica. Porén, teñen características que son comúns a varios virus e a miúdo denomínanse axentes subvirais.[42] Os viroides son patóxenos importantes das plantas.[43] Non codifican proteínas pero interactúan coa célula hóspede e utilizan a maquinaria do hóspede para a súa replicación.[44] O virus da hepatite delta ou D dos humanos ten un xenoma de ARN similar ao dos viroides pero ten unha cuberta proteica derivada do virus da hepatite B e non pode producir a súa propia cuberta. É, por tanto, un virus defectivo e non pode propagar sen a axuda do virus da hepatite B.[45] De modo similar, o virófago Sputnik depende do mimivirus, o cal infecta ao protozoo Acanthamoeba castellanii.[46] Estes virus, que dependen da presenza doutra especie de virus na célula hóspede, chámanse virus satélites e poden representar intermediarios evolutivos entre os viroides e os virus.[47][48]
No pasado, todas estas hipóteses presentaban problemas. A hipótese regresiva non explica por que mesmo os parasitos celulares máis pequenos non se parecen aos virus en ningún modo. A hipótese do escape non explica a orixe das cápsides e outras estruturas tan complexas que presentan as partículas víricas. A hipótese dos virus primeiro contravén a definición de virus, xa que estes requiren células hóspedes e a hipótese non.[38] Os virus recoñécense agora como moi antigos e con orixes que precederon ao momento da diverxencia da vida en tres dominios.[49] Este descubrimento levou aos modernos virólogos a reconsiderar e reavaliar estas tres hipóteses clásicas.[49]
As evidencias que apoian a existencia dun mundo ancestral de ARN[50] e as análises computacionais de secuencias de ADN viral e de hóspedes están permitindo comprender mellor as relacións evolutivas entre diferentes virus e poden axudar a identificar os devanceiros dos virus modernos. Ata agora, ditas análises non probaron cal destas hipóteses é correcta.[50] Porén, parece improbable que todos os virus coñecidos actualmente teñan un antepasado común, e os virus probablemente se orixinaron numerosas veces no pasado por un ou varios mecanismos.[51]
Algunhas proteínas virais determinadas están presentes en virus que infectan a seres vivos dos tres dominios da vida,[52] pero a maioría das proteínas virais non teñen homólogas nos organismos celulares actuais, todo o cal parece indicar que son moi antigos. Algúns tipos de virións seguramente se orixinaron independentemente antes que LUCA, o último antepasado común universal da vida celular, como indican os estudos das proteínas da cápside. Os estudos moleculares indican tamén que os virus, ao seren parasitos intracelulares obrigados, coevolucionaron cos seus hóspedes.[53] Os virus tenden a adaptarse ao seu hóspede natural e, se levan longo tempo evolucionando nel, causan enfermidades leves ou limitadas, propagándose sen afectaren demasiado ao organismo hóspede, pero non así cando infectan a un hóspede que non é o seu hóspede habitual ou levan pouco tempo nunha especie hóspede, xa que nese caso adoitan ser máis virulentos.
Os prións son moléculas de proteínas infecciosas que non conteñen ADN ou ARN.[54] Poden causar infeccións como o mal das ovellas tolas ou scrapie, a encefalopatía esponxiforme bovina ("enfermidade das vacas tolas") nos vacúns e a enfermidade debilitante crónica nos cervos; nos humanos entre as enfermidades priónicas están o kuru, enfermidade de Creutzfeldt-Jakob e a síndrome de Gerstmann-Sträussler-Scheinker.[55] Aínda que os prións son esencialmente diferentes dos virus e viroides, o seu descubrimento deulle creto á teoría de que os virus poderían ter evolucionado a partir de moléculas autorreplicantes.[56]
Microbioloxía
[editar | editar a fonte]Propiedades vitais
[editar | editar a fonte]Aínda que a opinión máis común é que os virus non deben considerarse seres vivos, hai distintos pareceres sobre se efectivamente deben considerarse formas de vida ou só estruturas orgánicas que interaccionan cos organismos vivos, e o debate incrementouse co descubrimento do Mimivirus, un virus cun xenoma de gran tamaño, e outros virus similares. Os virus foron descritos como "organismos no limiar da vida",[9] xa que se parecen aos organismos en que posúen xenes e evolucionan por selección natural,[57] e reprodúcense creando múltiples copias de si mesmos por autoensamblaxe dos seus compoñentes. Aínda que teñen xenes, non teñen estrutura celular, a cal se considera comunmente como a unidade básica da vida. Os virus non teñen o seu propio metabolismo, e requiren unha célula hóspede para fabricar novos produtos. Por tanto, non poden reproducirse naturalmente fóra da súa célula hóspede.[58] (aínda que especies bacterianas como as rickettsias e clamidias son consideradas organismos vivos a pesar de teren esa mesma limitación).[59][60] As formas de vida aceptadas como tales utilizan a división celular para reproducirse, mentres que os virus o que fan é ensamblarse espontaneamente dentro de células. Isto é tamén distinto do crecemento autónomo dun cristal, xa que herdan mutacións xenéticas ao estaren suxeitos á selección natural. A autoensamblaxe dos virus dentro das células ten implicacións para o estudo da orixe da vida, xa que dan maior creto á hipótese de que a vida puido comezar como unha autoensamblaxe de moléculas orgánicas.[1]
Estrutura
[editar | editar a fonte]



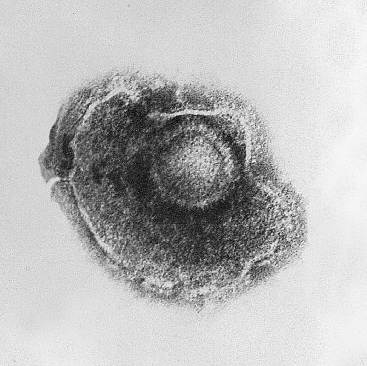

Os virus presentan unha gran diversidade de formas e tamaños na súa morfoloxía. En xeral, os virus son moito máis pequenos cás bacterias. A maioría dos virus que foron estudados teñen un diámetro entre 20 e 300 nanómetros. Algúns filovirus teñen unha lonxitude total de ata 1.400 nm, pero os seus diámetros son só duns 80 nm.[61] A maioría dos virus non poden verse co microscopio óptico, polo que se debe usar o microscopio electrónico para visualizar os virións.[62] Para incrementar o contraste entre os virus e o fondo onde se encontran, utilízanse "tinguiduras" electrodensas, que son solucións de sales de metais pesados, como o volframio, que dispersan os electróns das rexións cubertas coa tinguidura. Cando os virións están cubertos coa tinguidura (tinguidura positiva), os detalles máis finos quedan escurecidos. A tinguidura negativa soluciona ese problema ao tinguir só o fondo.[63]
Unha partícula vírica completa, coñecida como virión, consta dun ácido nucleico rodeado dunha cuberta protectora de proteínas chamada cápside. As cápsides están formadas por subunidades proteicas idénticas chamadas capsómeros.[64] Os virus poden ter unha "envoltura" lipídica derivada das membranas celulares das células hóspede. As proteínas da cápside están codificadas polo xenoma viral. A forma da cápside serve para distinguir os virus morfoloxicamente.[65][66] As subunidades proteicas codificadas polo virus autoensámblanse para formar a cápside, e en xeral requiren a presenza do xenoma do virus. Os virus complexos codifican proteínas que colaboran na construción da cápside. As proteínas asociadas co ácido nucleico coñécense como nucleoproteínas, e a asociación das proteínas da cápside viral co ácido nucleico viral denomínase nucleocápside. A cápside e a estrutura enteira do virus poden ser sondadas fisicamente con microscopio de forza atómica.[67][68]
En xeral, hai catro tipos morfolóxicos de virus, que son:
- Helicoidal
- Estes virus están compostos por un só tipo de capsómero disposto arredor dun eixe central para formar unha estrutura helicoidal, a cal pode ter unha cavidade central ou un tubo oco. Esta disposición orixina virións coa forma de bastón ou filamentosos: estes poden ser curtos e moi ríxidos, ou longos e moi flexibles. O material xenético, que, en xeral, é ARN monocatenario, pero nalgúns casos ADN monocatenario, está ligado ás proteínas por interaccións entre o ácido nucleico cargado negativamente e as cargas positivas das proteínas. En conxunto, a lonxitude dunha cápside helicoidal está relacionada coa lonxitude do ácido nucleico que contén e o diámetro depende do tamaño e disposición dos capsómeros. O virus do mosaico do tabaco é un exemplo de virus helicoidal.[69]
- Icosaédrico
- A maioría dos virus animais son icosaédricos ou case esféricos con simetría icosaédrica. Adoptar a forma dun icosaedro regular é a maneira óptima de formar unha cuberta pechada con subunidades idénticas. O mínimo número de capsómeros idénticos que se require é doce, cada un composto por cinco subunidades idénticas. Moitos virus, como os rotavirus, teñen máis de doce capsómeros e parecen esféricos pero conservan esta simetría. Os capsómeros dos ápices están rodeados doutros cinco capsómeros e denomínanse pentóns. Os capsómeros das caras triangulares están rodeados doutros seis e chámanse hexóns.[70] Os hexóns son esencialmente planos. Os pentóns están curvados e forman os 12 vértices do icosaedro. A mesma proteína pode actuar como subunidade tanto nos pentámeros coma nos hexámeros ou poden estar compostos por proteínas diferentes.
- Prolato
- É unha variante do icosaédrico, xa que se pode considerar como un icosaedro alongado ao longo dun eixe e é unha disposición común nas cabezas dos bacteriófagos. Esta estrutura está composta por un cilindro cunha tapa en ambos os extremos.[71]
- Con envoltura
- Algunhas especies de virus están envoltos nunha forma modificada dalgunha membrana celular, xeralmente a membrana externa que rodea a célula hóspede infectada (membrana plasmática) ou membranas internas como a nuclear ou a do retículo endoplasmático. Deste modo o virus adquire unha bicapa lipídica externa chamada envoltura viral. Esta membrana está inzada de proteínas codificadas polos xenomas viral e do hóspede; a membrana lipídica e os carbohidratos que presente son orixinarios enteiramente do hóspede. O virus da gripe e o VIH utilizan esta estratexia. A maioría dos virus con envoltura dependen dela para a súa infectividade.[72]
- Complexa
- Estes virus posúen unha cápside que non é nin puramente helicoidal nin puramente icosaédrica, e que pode posuír estruturas extra como colas de composición proteica ou unha parede externa complexa. Algúns bacteriófagos, como o fago T4 de Enterobacterias, ten unha estrutura complexa que consiste nunha cabeza icosaédrica unida a unha cola helicoidal, a cal pode ter unha placa hexagonal na base con fibras da cola proteicas que sobresaen. Esta estrutura da cola actúa como unha xiringa molecular, que serve para unirse á superficie do hospede bacteriano e inxectar o xenoma viral dentro da célula.[73]
Os poxvirus son grandes virus complexos que teñen unha morfoloxía pouco común. O xenoma viral está asociado con proteínas dentro dunha estrutura discoidal central chamada nucleoide. O nucleoide está rodeado por unha membrana e dous corpos laterais de función descoñecida. O virus ten unha envoltura externa cunha grosa capa de proteínas chatoladas sobre a súa superficie. O virión completo é lixeiramente pleomórfico, xa que a súa forma vai desde ovoide a coa forma de ladrillo.[74] O mimivirus era ata hai pouco o virus máis longo coñecido, cun diámetro de cápside de 400 nm. Desde a súa superficie proxéctanse filamentos proteicos que miden 100 nm. A cápside aparece hexagonal vista ao microscopio electrónico, e, por tanto, dita cápside é probablemente do tipo icosaédrico.[75] En 2011, descubriuse un virus aínda maior no fondo oceánico da costa do Chile, que foi chamado provisionalmente Megavirus chilensis, e pode verse mesmo con microscopio óptico. [76]
Algúns virus que infectan arqueas teñen estruturas complexas que non están relacionadas con ningunha outra forma presente nos virus, cunha ampla variedade de morfoloxías, que van desde estruturas con forma de fuso, a virus que lembran bastóns con gancho, bágoas ou mesmo botellas. Outros virus de arqueas lembran aos bacteriófagos con cola, e poden ter diversas estruturas de cola.[77]
Xenoma
[editar | editar a fonte]| Propiedade | Parámetros |
|---|---|
| Ácido nucleico |
|
| Forma |
|
| Número de cadeas |
|
| Sentido |
|
As distintas especies de virus presentan unha enorme variedade de estruturas xenómicas, e, como grupo, os virus conteñen máis diversidade xenómica estrutural cás plantas e animais, arqueas, ou bacterias. Hai millóns de tipos de virus,[5] aínda que só 5.000 foron descritos en detalle.[4] O ácido nucleico que leva os xenes dos virus é ou ben ADN ou ARN, polo que se denominan virus de ADN ou virus de ARN, respectivamente. A gran maioría dos virus teñen xenomas de ARN. Os virus das plantas adoitan a ter xenomas de ARN monocatenario e os bacteriófagos de ADN bicatenario.[78]
Os xenomas virais poden ser circulares, como nos poliomavirus, ou lineares, como nos adenovirus. O tipo de ácido nucleico é irrelevante con respecto á forma do xenoma. Entre os virus de ARN e certos virus de ADN, o xenoma está a miúdo dividido en partes separadas, polo que se chama segmentado. Nos virus de ARN, cada segmento xeralmente codifica unha soa proteína e o máis común é que os segmentos se encontren xuntos dentro dunha mesma cápside. Porén, non todos os segmentos teñen que estar no mesmo virión para que o virus sexa infeccioso, como se demostra no virus do mosaico do Bromus (afecta a planta Bromus inermis) e noutros virus de plantas.[61]
Un xenoma viral, independentemente do seu tipo de ácido nucleico, é case sempre monocatenario ou bicatenario, xa que os mixtos son raros. Os xenomas monocatenarios constan dun ácido nucleico coas bases non apareadas. Os xenomas bicatenarios constan de dúas cadeas complementarias con bases apareadas. As partículas víricas dalgunhas familias de virus, como as que pertencen aos Hepadnaviridae, conteñen un xenoma que é parcialmente de dobre cadea e parcialmente monocatenario.[78]
Na maioría dos xenomas víricos de ARN e algúns con xenomas de ADN monocatenario, as cadeas simples poden ser de sentido positivo (chamadas cadeas máis ou +) ou ben de sentido negativo (chamadas cadeas menos ou -), segundo sexan ou non complementarias ao ARN mensaxeiro viral. O ARN viral de sentido positivo ten o mesmo sentido có ARNm viral e así polo menos unha parte del pode ser traducido inmediatamente pola célula hóspede. O ARN viral de sentido negativo é complementario do ARNm e debe ser convertido nun ARN de sentido positivo por unha ARN polimerase ARN dependente antes da súa tradución. A nomenclatura do ADN para os virus de ADN monocatenario xenómico dun só sentido é similar á nomenclatura do ARN en que a cadea codificante para o ARNm viral é complementaria dela (−), e a cadea non codificante é unha copia dela (+).[78] Con todo, varios tipos de virus de ADN monocatenario e ARN monocatenario teñen xenomas que son ambisentido nos que se forma un intermediato replicativo de dobre cadea. Exemplos de virus con xenoma ambisentido son os xeminivirus, que son virus de ADN monocatenario de plantas, e os arenavirus, que son virus de ARN monocatenario de animais.[79]
O tamaño dos xenomas varía moito entre as especies. Os xenomas virais máis pequenos (os dos circovirus de ADN monocatenario da familia Circoviridae) codifican só dúas proteínas e teñen un tamaño de xenoma de só 2 quilobases, e os máis grandes (o dos mimivirus) teñen tamaños xenómicos dunhas 1,2 megabases e codifican unhas mil proteínas.[80] Os xenomas dos virus de ADN grande nucleocitoplasmáticos, grupo ao que pertence o mimivirus, son excepcionais, xa que son dun tamaño moi grande para un virus, codifican moitas proteínas e son comparables aos dalgunhas bacterias. En xeral, os virus de ARN teñen tamaños xenómicos menores cós virus de ADN debido a unha maior taxa de erro durante a replicación, e teñen un tamaño superior máximo límite.[33] Alén deste límite, os erros no xenoma que se producen durante a replicación fan que o virus sexa inoperante ou non competitivo. Para compensar isto, os virus de ARN con frecuencia teñen xenomas segmentados (o xenoma divídese en moléculas máis pequenas) reducindo así a posibilidade de que un erro nun só compoñente do xenoma incapacite a totalidade do xenoma. Polo contrario, os virus de ADN xeralmente teñen xenomas máis longos a causa da maior fidelidade de copia que posúen os seus encimas replicativos.[81] Os virus de ADN monocatenario son unha excepción a esta regra, xa que as taxas de mutación nestes xenomas poden aproximarse ao caso extremo dos virus de ARN monocatenario.[82]

Os virus poden sufrir cambios xenéticos por medio de varios mecanismos. Estes inclúen un proceso chamado deriva antixénica na que as bases do ADN ou ARN mutan a outras bases. A maioría destas mutacións puntuais son "silenciosas", é dicir, non causan cambios na proteína codificada polo xene, pero outras poden proporcionar vantaxes evolutivas como a resistencia a fármacos antivirais.[83][84] O cambio antixénico (antigenic shift) ten lugar cando hai un cambio importante no xenoma do virus. Isto pode ser o resultado da recombinación xenética ou de redistribución (reassortment). Cando isto ocorre co virus da gripe poden orixinarse pandemias.[85] Os virus de ARN ás veces existen como quasiespecies ou enxames de virus da mesma especie pero con secuencias xenómicas lixeiramente distintas. Estas quasiespecies son unha das dianas principais da selección natural.[86]
Os xenomas segmentados teñen tamén vantaxes evolutivas. Distintas cepas dun virus cun xenoma segmentado poden intercambiar e combinar xenes e producir unha proxenie de virus que ten características únicas. Isto denomínase redistribución ou sexo viral.[87]
A recombinación xenética é o proceso polo cal unha fibra de ADN rompe e despois volve a unirse ao extremo dunha molécula diferente da orixinal. Isto pode ocorrer cando varios virus infectan a unha célula simultaneamente, e os estudos da evolución viral indican que a recombinación é un proceso moi estendido nas especies estudadas.[88] A recombinación é común tanto nos virus de ADN coma nos de ARN.[89][90]
Ciclo de replicación
[editar | editar a fonte]As poboacións virais non crecen por división celular, porque os virus son acelulares. O que fan é utilizar a maquinaria e metabolismo da célula hóspede para producir múltiples copias de si mesmos, fabricando os seus compoñentes e ensamblándoos na célula.


O ciclo de vida dos virus difire moito entre especies pero pódese dicir que hai seis estadios básicos nun ciclo de vida viral típico, que son:[91]
- Adhesión á superficie celular (ou adsorción). É a unión específica entre as proteínas da cápside viral ou da envoltura con receptores específicos da superficie da célula hóspede. Estes receptores teñen outras misións na célula, pero o virus utilízaos para a súa adhesión. Esta especificidade determina o rango de hóspedes (e mesmo os tipos celulares) aos que pode infectar o virus. Por exemplo, o VIH infecta só a un determinado tipo de células humanas, principalmente leucocitos. Isto é así porque a súa proteína de superficie, gp120, interacciona especificamente co receptor celular de quimiocinas CD4, que se encontra principalmente na superficie das células T CD4+. Este mecanismo evolucionou favorecendo os virus que infectan só a células nas cales son capaces de replicarse. A adhesión ao receptor pode inducir que a proteína viral da envoltura sufra cambios que orixinen a fusión das bicapas lipídicas da envoltura viral e da membrana plasmática celular, ou cambios nas proteínas das cápsides dos virus sen envoltura que faciliten a entrada do virus na célula.
- Penetración. Os virións entran na célula hóspede por medio dunha endocitose mediada por receptor ou fusión de membranas. Isto chámase xeralmente entrada viral. A infección de células de plantas e fungos é forzosamente diferente á de células animais, porque ditas células teñen paredes celulares ríxidas de celulosa ou quitina, polo que moitos virus só poden penetrar na célula se houbo un trauma que produciu danos na parede celular.[92] Con todo, unha vez dentro, case todos os virus de plantas (como o virus do mosaico do tabaco) poden tamén moverse directamente de célula a célula, en forma de complexos nucleoproteínicos monocatenarios, que atravesan os poros que comunican as células chamados plasmodesmos.[93] As bacterias, igual cás plantas, teñen fortes paredes celulares que o virus debe romper para infectar a célula, pero como as paredes bacterianas son moito menos grosas cás das plantas debido ao pequeno tamaño destes microorganismos, nalgúns virus evolucionaron mecanismos que inxectan o seu xenoma na célula bacteriana a través da parede celular, deixando as cápsides (baleiras) fóra da célula.[94]
- Decapsidación. É o proceso polo cal se elimina a cápside viral. Pode tratarse dunha degradación feita por encimas virais ou por encimas do hóspede ou ser unha simple disociación dos seus compoñentes. O resultado final é a liberación do ácido nucleico vírico que estaba dentro.
- Replicación. A replicación do virus implica principalmente a multiplicación do seu xenoma. A replicación depende da síntese de ARN mensaxeiro viral dos xenes "temperáns", que son os primeiros en expresarse, (con excepcións nos virus de ARN de sentido positivo), a síntese de proteínas virais, a posible ensamblaxe das proteínas virais e despois a replicación do xenoma mediado pola expresión de proteínas temperás ou regulatorias. Isto pode ir seguido, en virus complexos de xenoma grande, dunha ou máis roldas de síntese de ARNm: a expresión de xenes "tardíos" é, en xeral, de proteínas estruturais ou do virión.
- Autoensamblaxe. Despois da autoensamblaxe mediada pola estrutura das partículas víricas, teñen lugar algunhas modificacións nas proteínas do virus, o que ás veces se chama maduración do virus. En virus como o VIH, esta modificación ocorre despois de que o virus foi liberado da célula, e noutros virus ten lugar dentro da célula.[95]
- Liberación. Os virus poden liberarse da célula pola lise desta, un proceso que mata a célula ao rebentar a súa membrana e, no seu caso, a parede celular. Isto é característico de moitos virus de bacterias e dalgúns virus animais. Algúns virus sofren un ciclo lisoxénico no cal o xenoma viral se incorpora e integra por recombinación xenética nun cromosoma da célula, xeralmente nun lugar específico. O xenoma viral integrado recibe o nome de "provirus" ou, no caso dos bacteriófagos, de "profago".[96] Nesta situación, sempre que a célula se divida, o xenoma viral tamén se replica, polo que todas as células fillas levan unha copia do provirus. O xenoma viral permanece principalmente latente, inactivo, dentro da célula hóspede; porén, en certo momento, o provirus ou profago pode dar lugar a virus activos, os cales poden lisar a célula hóspede (ciclo lítico).[97] Os virus con envoltura (caso do VIH) libéranse da célula por evaxinación na membrana. Durante este proceso o virus adquire a súa envoltura, que é unha parte da membrana celular (da plasmática ou de membranas de orgánulos), modificada engadindo proteínas do virus.[98]
O material xenético dentro das partículas víricas, e o método polo cal se replica o material, varía considerablemente entre diferentes tipos de virus.[99]
- Virus de ADN
- A replicación do xenoma da maioría dos virus de ADN ten lugar no núcleo celular. Se a célula ten o receptor apropiado na súa superficie, estes virus entran na célula ás veces por fusión directa coa membrana da célula (por exemplo, os herpesvirus) ou, máis comunmente, por unha endocitose mediada por receptor. A maioría dos virus de ADN son totalmente dependentes da maquinaria celular de síntese de ARN e ADN, e de procesamento do ARN, pero os virus con xenomas máis grandes poden codificar parte desta maquinaria no seu propio xenoma. Nos eucariotas o xenoma viral debe cruzar os límites da membrana nuclear para acceder a esta maquinaria, mentres que nas bacterias só cómpre que entre na célula.[100]
- Virus de ARN
- A replicación xeralmente ten lugar no citoplasma. Os virus de ARN poden clasificarse en catro grupos distintos segundo o seu modo de replicación. A polaridade ou sentido dos virus de ARN monocatenarios (é dicir, se usan, ou non, directamente o seu ARN para sintetizar proteínas nos ribosomas) determina en gran medida o mecanismo de replicación; outro criterio importante é se o material xenético é bicatenario ou monocatenario. Todos os virus de ARN utilizan os seus propios encimas ARN replicases para crearen copias dos seus xenomas.[101]
- Virus con reversotranscrición
- Son virus de ARN monocatenario (Retroviridae, Metaviridae, Pseudoviridae) ou de ADN bicatenario (Caulimoviridae e Hepadnaviridae). Os virus que fan a reversotranscrición dos seus xenomas de ARN (retrovirus), usan unha molécula intermediaria de ADN para replicarse, mentres que os que teñen xenomas de ADN (pararretrovirus) utilizan un intermediario de ARN durante a replicación do xenoma. Ambos os tipos usan o encima reversotranscriptase, ou ADN polimerase-ARN-dependente, para levar a cabo a conversión do ácido nucleico de ARN a ADN. Os retrovirus integran o ADN producido por reversotranscrición no xenoma do seu hóspede, orixinando así un provirus como parte do seu proceso de replicación. Os pararretrovirus non fan isto, aínda que as copias do xenoma integradas de pararretrovirus de plantas poden dar lugar a virus infecciosos.[102] Estes virus son susceptibles a fármacos antivirais que inhiban a reversotranscriptase, por exemplo, a zidovudina e a lamivudina. Un exemplo do primeiro tipo é o VIH, que é un retrovirus. Exemplos do segundo tipo son os Hepadnaviridae, entre os que está o virus da hepatite B.[103]
Efectos sobre a célula hóspede
[editar | editar a fonte]A gama de efectos estruturais e bioquímicos da infección vírica sobre a célula hóspede é ampla.[104] Denomínanse efectos citopáticos.[105] A maioría das infeccións víricas orixinan finalmente a morte da célula hóspede. As causas da morte poden ser a lise celular, alteracións na superficie da membrana da célula e apoptose.[106] Con frecuencia a morte celular está causada polo cesamento das súas actividades normais debido á supresión exercida por proteínas víricas específicas, non todas as cales son compoñentes da partícula vírica.[107]
Algúns virus non causan cambios aparentes na célula infectada. As células nas que o virus está latente e inactivo mostran poucos sinais de infección e a miúdo funcionan normalmente.[108] Isto causa infeccións persistentes e o virus acostuma estar en estado dormente durante moitos meses ou anos. Isto é común nos virus do herpes.[109][110] Algúns virus, como o de Epstein-Barr, poden causar unha proliferación celular sen orixinar un tumor maligno,[111] e outros, como os papilomavirus, poden causar cancro.[112]
Rango de hóspedes
[editar | editar a fonte]Os virus son, con gran diferenza, as entidades biolóxicas máis abundantes da Terra e o seu número supera o de todos os seres vivos xuntos.[113] Infectan a todo tipo de vida celular, tanto de animais coma de plantas, fungos, protistas ou bacterias.[4] Porén, cada tipo de virus pode infectar só un limitado rango de hóspedes e moitos son específicos de especie. Algúns, como o virus da varíola por exemplo, poden infectar só unha especie, a humana neste caso,[114] e dise que teñen un rango de hóspedes estreito. Outros virus, como o virus da rabia, poden infectar diferentes especies de animais e dise que teñen un rango amplo.[115] Os virus que infectan plantas son inofensivos para os animais, e a maioría dos virus que infectan outros animais son inofensivos para os humanos.[116] O rango de hóspedes dalgúns bacteriófagos está limitado a unha soa cepa de bacterias e poden utilizarse para rastrear a fonte de estalidos infecciosos por un método chamado tipificación de fagos.[117]
Clasificación
[editar | editar a fonte]- Artigo principal: Clasificación dos virus.
A clasificación trata de describir a diversidade dos virus dándolles nome e agrupándoos segundo as súas semellanzas. En 1962, André Lwoff, Robert Horne e Paul Tournier foron os primeiros que elaboraron un método de clasificación dos virus, chamado LHT, baseado no sistema xerárquico da taxonomía de Linneo.[118] Este sistema utilizaba as categorías de filo, clase, orde, familia, xénero e especie. Os virus agrupábanse de acordo coas propiedades que compartían (e non coas dos seus hóspedes) e o tipo de ácido nucleico que formaba os seus xenomas.[119] Máis tarde formouse o Comité Internacional de Taxonomía de Virus (ICTV), que elaborou unha clasificación na que os virus non están clasificados en filos ou clases, xa que o pequeno tamaño do seu xenoma e alta taxa de mutación fai difícil determinar os seus antepasados alén da categoría de orde. Como complemento a esta clasificación utilízase a clasificación de Baltimore.
Clasificación do ICTV
[editar | editar a fonte]O Comité Internacional de Taxonomía de Virus (International Committee on Taxonomy of Viruses, ICTV) elaborou o sistema de clasificación actual dos virus e publicou directrices que facían grande énfase en certas propiedades dos virus para manter a uniformidade das familias víricas. Estableceuse así unha taxonomía unificada (un sistema universal para clasificar os virus). O 9º informe do ICTV[120] define o concepto de especie vírica como o taxon máis baixo da xerarquía dos taxons virais.[121] Con todo, só se estudou unha pequena parte da diversidade total de virus, e as análises de mostras de procedencia humana encontraron que arredor do 20% das secuencias de virus recollidas non foran vistas nunca antes, e as mostras recollidas do ambiente, como as procedentes de auga mariña e sedimentos oceánicos, atoparon que a gran maioría das secuencias eran completamente novas.[122]
A estrutura taxonómica xeral é a seguinte:
- Orde (-virales)
- Familia (-viridae)
- Subfamilia (-virinae)
- Familia (-viridae)
Na taxonomía do ICTV publicada no 2011, distínguense seis ordes, os Caudovirales, Herpesvirales, Mononegavirales, Nidovirales, Picornavirales e Tymovirales. Propúxose ademais unha sétima orde, os Ligamenvirales. O Comité non distingue formalmente entre subespecies, cepas e illamentos. No total hai 6 ordes, 87 familias, 19 subfamilias, 349 xéneros, arredor de 2.284 especies e uns 3.000 tipos aínda non clasificados.[120][123][124]
Clasificación de Baltimore
[editar | editar a fonte]- Artigo principal: Clasificación de Baltimore.

O biólogo e premio Nobel David Baltimore ideou o sistema de clasificación dos virus coñecido como clasificación de Baltimore.[30][125] O sistema de clasificación do ICTV utilízase conxuntamente con estoutro nas clasificacións dos virus modernas.[126][127][128]
A clasificación de Baltimore dos virus está baseada no mecanismo utilizado para a produción do ARNm. Os virus deben xerar os seus ARNm a partir dos seus xenomas para poder producir proteínas e replicarse, pero utilizan diferentes mecanismos para facelo en cada familia vírica. Os xenomas virais poden ser monocatenarios ou bicatenarios, de ARN ou de ADN, e poden ou non utilizar a reversotranscriptase (RT). Ademais, os virus de ARN monocatenario poden ser de sentido positivo (+) ou antisentido (−). Esta clasificación establece sete grupos:
- I: Virus de ADN bicatenarios (por exemplo, Adenovirus, Herpesvirus, Poxvirus).
- II: Virus de ADN monocatenarios (de fibra + ou sentido) (por exemplo, Parvovirus).
- III: Virus de ARN bicatenarios (por exemplo, Reovirus).
- IV: Virus de ARN monocatenarios (+) (de fibra + ou sentido) (por exemplo, Picornavirus, Togavirus).
- V: Virus de ARN monocatenarios (−) (de fibra − ou antisentido) (por exemplo, Orthomyxovirus, Rhabdovirus).
- VI: Virus de ARN monocatenario-RT (de fibra + ou sentido) con retrotranscrición e cun intermediario de ADN no seu ciclo vital (por exemplo, Retrovirus).
- VII: Virus de ADN bicatenario-RT con retrotranscrición (por exemplo, Hepadnavirus).
Como exemplo da clasificación viral, o virus da varicela, chamado virus da varicela zóster (VZV), pertence á orde Herpesvirales, familia Herpesviridae, subfamilia Alphaherpesvirinae e xénero Varicellovirus (na clasificación do ICTV). Ademais está encadrado no grupo I da clasificación de Baltimore porque é un virus de ADN bicatenario que non utiliza reversotranscriptase.
Axentes subvirais e prións
[editar | editar a fonte]Outros axentes similares a virus, que tamén son estudados pola viroloxía, pero que non presentan todas as propiedades da definición tradicional de virus (non teñen cápside proteica, non se replican na falta doutros virus), e non están incluídos nas clasificacións dos virus normais, son os chamados axentes subvirais. Entre eles están:
- Viroides. Formados por ARN circular monocatenario sen cápside. Parasitan plantas. Non codifican proteínas e replícanse utilizando encimas celulares. Non necesitan a presenza doutro virus para a replicación.[129]
- Satélites. Formados só por ácido nucleico, hainos de ADN e de ARN[130] (estes denomínanse ácidos nucleicos satélites, e os de ARN circular monocatenario chámanse virusoides)[131], ou por ácido nucleico e cápside proteica (estes serían virus satélites coa morfoloxía típica dos virus). Os satélites non se replican autonomamente aproveitándose da maquinaria da célula, senón que deben infectar a célula conxuntamente con outro virus dunha especie determinada, do cal dependen para a súa replicación. Algúns satélites afectan negativamente ou impiden a replicación do virus axudante co que coinfectan e denomínanse virófagos.[132]
Outros axentes estudados pola viroloxía son os prións. Os prións non son virus, xa que se trata de proteínas sen ácido nucleico, pero con capacidade infecciosa. Segundo a interpretación común, os prións adoptan unha conformación tridimensional anormal que causa unha enfermidade e que pode propagarse a outras proteínas iguais que tiñan a conformación normal. Poden transmitirse dun individuo a outro, polo que pode dicirse que teñen carácter infeccioso. Exemplos de doenzas causada por prións son a síndrome de Creutzfeldt-Jakobs [133] e a encefalopatía esponxiforme bovina (mal das vacas tolas).
En 2024 descubriuse un novo axente subviral similar a un viroide con xenoma de ARN e secuencias distintas ás de todas as formas de vida coñecidas, aos que se lles chamou obeliscos.[134]
Os virus nas enfermidades humanas
[editar | editar a fonte]
Exemplos de enfermidades humanas comúns causadas por virus son o arrefriado común, a gripe, a varicela e o herpes labial. Moitas enfermidades serias como o ébola, a SIDA, a gripe aviaria e a SARS son tamén causadas por virus. A capacidade relativa dos virus de causar doenzas denomínase virulencia. Estanse a investigar outras doenzas para comprobar se teñen un virus como axente causante, como a posible conexión entre o herpesvirus humano 6 (HHV6) e enfermidades neurolóxicas como a esclerose múltiple e a síndrome de fatiga crónica.[137] Hai controversia sobre se o bornavirus, que previamente se pensaba que causaba enfermidades neurolóxicas en cabalos, podería ser responsable de enfermidades psiquiátricas en humanos.[138]
Os virus utilizan diferentes mecanismos para producir enfermidades nun organismo, o cal depende en gran medida da especie viral de que se trate. Os mecanismos a nivel celular inclúen principalmente a lise celular, isto é, a saída da célula rompendo a súa membrana coa conseguinte morte da célula. En organismos multicelulares, se morre un número suficiente de células, todo o organismo empezará a sufrir os efectos. Aínda que os virus distorsionan a homeostase normal saudable, orixinando doenzas, poden existir dentro dun organismo de forma relativamente inofensiva. Un exemplo é a capacidade do virus herpes simplex, que causa o herpes labial, de permanecer en estado dormente no corpo humano, o que se denomina latencia,[139] e que é unha característica dos virus herpes, como o virus de Epstein-Barr, que causa a febre glandular, e o virus varicela zóster, que causa a varicela e o herpes zóster. A maioría da xente foi infectada por algún tipo de herpes.[140] Porén, estes virus latentes poderían en certas circunstancias ser beneficiosos, xa que ás veces a presenza do virus pode incrementar a inmunidade contra patóxenos bacterianos como Yersinia pestis.[141]
Algúns virus causan infeccións crónicas que duran toda a vida, nas que os virus continúan replicándose no corpo a pesar dos esforzos dos mecanismos de defensa do hóspede.[142] Isto é común nas infeccións polos virus das hepatites B e C. As persoas infectadas cronicamente denomínanse portadores, xa que serven de reservorios de virus infecciosos.[143] En poboacións cunha alta proporción de portadores, dise que a enfermidade é endémica.[144]
Epidemioloxía
[editar | editar a fonte]A epidemioloxía viral é a rama da medicina que trata da transmisión e control das infeccións víricas en humanos. A transmisión dos virus pode ser vertical, o que significa que se transmite de nai a fillo, ou horizontal, que significa que se transmite dunha persoa a outra. Exemplos de transmisión vertical son a do virus da hepatite B e do VIH, na que o meniño xa nace infectado co virus.[145] Outro exemplo máis raro é o do virus da varicela zóster, o cal, a pesar de que causa infeccións relativamente leves nos humanos, pode ser letal para o feto ou o neonato.[146]
A transmisión horizontal é o mecanismo máis común de transmisión das infeccións víricas nas poboacións. A transmisión pode ocorrer nos seguintes casos:
- intercambios de fluídos corporais durante a actividade sexual, por exemplo, no caso do VIH;
- intercambio de sangue por transfusións contaminadas ou por compartir agullas ou xiringas, por exemplo, a hepatite C ou o VIH;
- intercambio de saliva pola boca, por exemplo, o virus de Epstein-Barr;
- inxestión de alimentos e auga contaminada, por exemplo, o norovirus;
- inhalación de aerosois que conteñen virións, por exemplo, o virus da gripe;
- inoculación por insectos vectores, como mosquitos que perforan a pel do hóspede, por exemplo, o virus do dengue.
A taxa ou velocidade de transmisión das infeccións virais depende de factores como a densidade de poboación, o número de individuos susceptibles (por exemplo, os que non son inmunes),[147] a calidade do servizo de recollida de lixo e o tempo climatolóxico.[148]
A epidemioloxía pode utilizarse para romper a cadea de infeccións en poboacións durante os estalidos de enfermidades virais.[149] Utilízanse medidas de control baseándose no coñecemento de como se transmite o virus. É importante atopar a fonte ou fontes do estalido e identificar o virus. Unha vez que o virus foi identificado, a cadea de transmisión pode interromperse ás veces polas vacinas. Cando non se dispón de vacinas, poden ser efectivas a hixiene e a desinfección. A miúdo, as persoas infectadas son illadas do resto da comunidade, e os que estiveron expostos ao virus deben pasar un período de corentena.[150] En epidemias en animais ás veces hai que sacrificar moitos animais.[151] A maioría das infeccións virais de humanos e doutros animais teñen un período de incubación durante o cal a infección non causa síntomas evidentes.[152] Os períodos de incubación das enfermidades virais varían desde uns poucos días a semanas.[153] Despois do período de incubación ou solapándose un pouco con el, hai un período en que o individuo ou animal infectado é contaxioso e pode infectar outras persoas ou animais.[153] Coñécense os períodos de incubación e de contaxio de moitas infeccións virais, o que é importante para o control das epidemias.[154] Cando o estalido infeccioso causa unha proporción anormalmente alta de casos na poboación, localidade ou rexión, denomínase epidémico. Se a infección se estende por todo o mundo, denomínase pandémica.[155]
Epidemias e pandemias
[editar | editar a fonte]
As poboacións indíxenas de América foron decimadas por enfermidades contaxiosas como a varíola, traídas a América polos colonizadores europeos. Non se sabe exactamente cantos indíxenas americanos morreron por estas enfermidades alleas que entraron no continente despois da chegada de Colón, pero estímase que foi preto do 70% da poboación. O dano feito por estas doenzas axudou significativamente os intentos europeos de desprazar e conquistar a poboación nativa.[156]
Unha pandemia é unha epidemia estendida por todo o mundo. A pandemia de gripe de 1918, que se prolongou ata 1919, tivo unha severidade de categoría 5 e foi causada por unha cepa do virus da gripe A infrecuentemente virulenta e mortal. As vítimas eran con frecuencia adultos mozos e saudables, ao contrario da maioría das epidemias de gripe, que afectan principalmente nenos, vellos, ou persoas debilitadas.[157] As estimacións vellas calculaban que matara a 40–50 millóns de persoas,[158] pero as máis recentes estiman que puido matar a 100 millóns, ou o 5% da poboación mundial en 1918.[159]
A maioría dos investigadores cre que o VIH se orixinou na África subsahariana durante o século XX,[160] e converteuse agora nunha pandemia. Calcúlase que hai hoxe no mundo 38,6 millóns de persoas que viven coa enfermidade.[161] O programa das Nacións Unidas contra a SIDA (UNAIDS) e a Organización Mundial da Saúde (OMS) estiman que a SIDA leva matado xa a máis de 25 millóns de persoas desde que se coñeceu en 1981, o que fai que se considere unha das epidemias máis destrutivas da historia.[162] En 2007 houbo 2,7 millóns de novas infeccións polo VIH e 2 millóns de mortes relacionadas co VIH.[163]

Varios virus patóxenos moi letais son membros dos Filoviridae. Os filovirus son virus do tipo filamentoso, que causan febre hemorráxica viral, e entre eles están o virus Ébola e o virus Marburg. O virus Marburg causou en 2004-2005 un estalido infeccioso en Angola, que foi a peor epidemia coñecida dunha febre hemorráxica viral.[164]
Cancro
[editar | editar a fonte]- Véxase tamén: Oncovirus.
Certos virus son causa ben establecida de cancro en humanos e outras especies. Os cancros virais ocorren só nunha minoría das persoas ou animais infectados. Os virus procancerosos pertencen a varias familias víricas, tanto de virus de ARN coma de ADN, polo que non hai un só tipo de "oncovirus" (un termo usado orixinalmente para os retrovirus agudamente transformantes). O desenvolvemento do cancro está determinado por unha variedade de factores como a inmunidade do hóspede[165] e as mutacións no hóspede.[166] Entre os virus que se aceptan como causa de cancros humanos están algúns xenotipos de papilomavirus humano, virus da hepatite B, virus da hepatite C, virus de Epstein-Barr, herpesvirus asociado ao sarcoma de Kaposi e virus T-linfotrópico humano. O virus do cancro humano descuberto máis recentemente é un poliomavirus (poliomavirus da célula de Merkel), que causa a maioría dos casos dunha rara forma de cancro de pel chamado carcinoma de célula de Merkel.[167] Os virus da hepatite poden orixinar infeccións crónicas que dean lugar a cancro de fígado.[168][169] A infección polo virus T-linfotrópico humano pode causar mielopatía asociada ao virus HTLV-1 e leucemia de célula T adulta.[170] Os papilomavirus humanos orixinan cancros de cérvix uterino, pel, ano e pene.[171] Entre os Herpesviridae, os herpesvirus asociados ao sarcoma de Kaposi causan sarcoma de Kaposi e certos linfomas, e o virus de Epstein-Barr causa o linfoma de Burkitt, linfoma de Hodgkin, trastornos linfoproliferativos de células B e carcinoma nasofarínxeo.[172] O poliomavirus de células de Merkel está moi relacionado co SV40 e poliomavirus de ratos, que foron utilizados como modelos animais de virus do cancro durante 50 anos.[173]
Mecanismos de defensa do hóspede
[editar | editar a fonte]- Véxase tamén: Sistema inmunitario.
A primeira liña de defensa do corpo contra os virus é o sistema inmunitario innato. Este consta de células e outros mecanismos que defenden o hóspede das infeccións dun modo non específico. Isto significa que as células do sistema innato recoñecen e responden aos patóxenos dun modo xenérico, pero, ao contrario do que ocorre coa acción do sistema inmunitario adaptativo, isto non confire ao hóspede unha inmunidade protectora de longa duración.[174]
A interferencia de ARN é unha importante defensa innata contra os virus.[175] Moitos virus teñen unha estratexia de replicación que implica a formación de ARN bicatenario. Cando eses virus infectan á célula, liberan a súa molécula ou moléculas de ARN, as cales se unen inmediatamente a un complexo proteico chamado Dicer que corta o ARN en pequenos anacos. Actívase unha vía bioquímica chamada complexo RISC, a cal degrada o ARNm viral e a célula sobrevive á infección. Os rotavirus evitan este mecanismo de defensa non decapsidándose por completo dentro da célula e liberando o novo ARNm producido a través de poros da cápside interna da partícula. O ARN de dobre cadea xenómico permanece protexido dentro da parte central (core) do virión.[176][177]
Cando o sistema inmunitario adaptativo dos vertebrados encontra un virus, produce anticorpos específicos, que se unen ao virus e a miúdo o converten en non infeccioso (neutralización do virus). Esta é a inmunidade humoral. Hai dous tipos de anticorpos que son os máis importantes. O primeiro son as IgM, que son moi efectivas neutralizando virus, pero que as células inmunitarias producen só durante unhas poucas semanas. O segundo, chamado IgG, prodúcese por tempo prolongado. A presenza ou non de IgM no sangue do hóspede utilízase para comprobar que hai unha infección aguda, mentres que a IgG indica que se produciu a infección nalgún momento pasado.[178] Os anticorpos IgG mídense cando se están realizando probas de inmunidade.[179]
Os anticorpos poden seguir sendo un mecanismo de defensa efectivo mesmo despois de que os virus conseguisen entrar na célula hóspede. Unha proteína celular chamada TRIM21 pode unirse aos anticorpos que permaneceron unidos á superficie da partícula do virus unha vez que este entrou na célula. Isto prepara a subseguinte destrución dos virus polos encimas do sistema do proteosoma da célula.[180]

Unha segunda defensa que teñen os vertebrados contra os virus denomínase inmunidade mediada por células e implica ás chamadas células T. As células do corpo despregan constantemente curtos fragmentos das súas proteínas na superficie celular, e, se unha célula T recoñece un fragmento viral sospeitoso alí, a célula hóspede é destruída polas células T citotóxicas e proliferan as células T específicas contra ese virus. Células como os macrófagos e células dendríticas son especialistas nesta presentación de antíxenos.[181]
A produción de interferón é outro mecanismo importante de defensa do hóspede. O interferón é unha proteína producida polas células do corpo cando están infectadas por virus. O seu papel na inmunidade é complexo; desencadea a produción de encimas antivíricos que finalmente impiden a reprodución do virus. O interferón gamma activa a actividade de macrófagos e células asasinas naturais.[182]
Non todas as infeccións víricas producen unha resposta inmune protectora por esta vía. O VIH evita o sistema inmunitario ao cambiar continuamente a secuencia das proteínas da superficie do virión e por permanecer un tempo latente integrado no cromosoma[183]. Estes virus persistentes evitan o control do sistema inmunitario secuestrando ou bloqueando a presentación de antíxenos, con resistencia ás citocinas, con evasión das actividades das células asasinas naturais, evitando a apoptose e con cambios antixénicos.[184] Outros virus chamados virus neurotrópicos, espállanse por propagación neural no sistema nervioso, onde o sistema inmune non pode acadalos.
Prevención e tratamento
[editar | editar a fonte]Como os virus utilizan vías metabólicas vitais da célula hóspede para replicarse, son difíciles de eliminar sen usar fármacos que causen efectos tóxicos ao hóspede en xeral. As estratexias médicas máis efectivas contra as enfermidades de orixe viral son a vacinación para adquirir inmunidade contra a infección, e as fármaco antivirais que interfiren selectivamente coa replicación viral.
Vacinas
[editar | editar a fonte]- Véxase tamén: Vacina.
A vacinación é un modo barato e efectivo de previr as infeccións víricas. As vacinas xa se usaban para previr as infeccións virais moito antes de que se descubrise a existencia dos virus. A primeira vacina obtida por Edward Jenner no século XVIII inmunizaba contra o virus da varíola. O seu uso supuxo un declive drástico na morbilidade (enfermidade) e mortalidade asociadas con infeccións virais como a polio, sarampelo, papeiras e rubéola.[185] As infeccións polo virus do sarampelo foron erradicadas nos países onde se fixo unha correcta campaña continuada de vacinación.[186] Hai vacinas efectivas para previr unhas trece infeccións virais nos humanos,[187] e moitas outras utilízanse para previr infeccións virais en animais.[188] As vacinas poden consistir en virus atenuados ou inactivados, ou en proteínas virais (antíxenos).[189] As vacinas de virus atenuados conteñen formas debilitadas do virus, que non poden causar a enfermidade, pero conteñen antíxenos que provocarán a inmunización. As vacinas de virus atenuados poden ser perigosas cando se administran a persoas cun sistema inmunitario debilitado (persoas inmunocomprometidas), porque nestas persoas, o virus debilitado pode causar a enfermidade.[190] Utilizando diversas técnicas biotecnolóxicas e a enxeñaría xenética poden producirse vacinas de subunidades. Nestas vacinas úsanse só as proteínas da cápside do virus. A vacina da hepatite B é un exemplo deste tipo de vacinas.[191] As vacinas de subunidades son seguras nos pacientes inmunocomprometidos porque non poden causar a enfermidade.[192] A vacina do virus da febre amarela, constituída por unha cepa atenuada chamada 17D, é probablemente a vacina máis segura e efectiva que se xerou ata agora.[193]
Fármacos antivirais
[editar | editar a fonte]

Os medicamentos ou fármacos antivirais son con frecuencia análogos de nucleósidos, (as unidades que forman o ADN unidas a fosfatos, que non son as auténticas no caso dos análogos), que os virus incorporan aos seus xenomas durante a replicación en lugar dos verdadeiros nucleósidos. O ciclo de vida do virus queda detido porque o novo ADN sintetizado é inactivo. Isto débese a que estes análogos carecen de grupos hidroxilo, os cales son os que se enlazan cos grupos fosfato para formar o esqueleto da molécula de ADN.[194] Exemplos de análogos de nucleósidos son o aciclovir utilizado contra o virus Herpes simplex e a lamivudina usada contra o VIH e o virus da hepatite B. O aciclovir é un dos fármacos antivirais máis antigos e máis usados.[195] Outros fármacos antivirais atacan outros estadios do ciclo de vida do virus. O VIH depende dun encima proteolítico chamado protease de VIH-1 para madurar e facerse plenamente infeccioso. Hai un tipo de fármacos chamados inhibidores da protease que inactivan este encima.
A hepatite C orixínase pola infección por un virus de ARN. No 80% das persoas infectadas, a enfermidade é crónica, e, se non reciben tratamento, quedan infectados para o resto da súa vida. Porén, hai agora un tratamento efectivo que utiliza o análogo de nucleósido ribavirina combinada con interferón.[196] Tamén se desenvolveu un tratamento para portadores asintomáticos crónicos do virus da hepatite B que utiliza unha estratexia similar usando lamivudina.[197]
Infeccións víricas noutras especies
[editar | editar a fonte]Os virus infectan a todos os tipos de vida celular e, aínda que os virus están presentes universalmente, cada especie celular é sensible á súa propia gama específica de virus, que con frecuencia infectan só a esas especies.[198] Algúns virus, chamados satélites, poden replicarse só en células que foron á vez infectadas por outro virus determinado.[46]
Virus de animais
[editar | editar a fonte]Os virus son patóxenos importantes do gando vacún. Enfermidades como a febre aftosa e a lingua azul son causadas por virus.[199] Os animais de compañía como gatos, cans e cabalos, se non están vacinados, son susceptibles de padecer infeccións virais serias. O parvovirus canino é un pequeno virus de ADN e as súas infeccións son con frecuencia mortais nas crías.[200] Como todos os invertebrados, a abella melífera é susceptible a moitas infeccións virais.[201] Porén, a maioría dos virus coexisten sen causar danos nos seus hóspedes e non causan síntomas de enfermidade.[3]
Virus de plantas
[editar | editar a fonte]
Existen moitos tipos de virus de plantas, pero a miúdo causan só unha perda do rendemento da colleita, e non é economicamente viable tratar de controlalas. Os virus de plantas son xeralmente transmitidos de planta a planta pola intermediación doutros organismos, denominados vectores. Estes son normalmente insectos, pero algúns fungos, vermes nematodos e os protozoos poden ser tamén vectores. Cando se considera economicamente viable controlar as infeccións víricas das plantas, os esforzos concéntranse en exterminar nesa área aos vectores e eliminar hóspedes alternativos, como malas herbas.[202] Os virus de plantas non poden infectar a humanos e a outros animais porque só poden reproducirse en células de plantas.[203]
As plantas teñen mecanismos de defensa elaborados e efectivos contra os virus. Un dos máis efectivos é a presenza dos chamados xenes de resistencia (R). Cada xene R confire resistencia a un virus determinado ao activar a morte celular localizada de certas partes da planta arredor da célula infectada, o cal pode a miúdo verse a primeira vista en forma de grandes manchas. Isto frea o espallamento da infección.[204] A interferencia de ARN é tamén unha defensa efectiva nas plantas.[205] Cando están infectadas, as plantas adoitan producir desinfectantes naturais que destrúen aos virus, como o ácido salicílico, óxido nítrico e moléculas reactivas do osíxeno.[206]
As partículas dos virus de plantas ou partículas similares a virus (VLPs) teñen aplicacións en biotecnoloxía e nanotecnoloxía. As cápsides da maioría dos virus de plantas son estruturas simples e robustas e poden producirse en grandes cantidades ou ben por infección das plantas ou pola expresión en diversos sistemas heterólogos. As partículas de virus de plantas poden modificarse xenética e quimicamente para encapsular material alleo e poden ser incorporadas en estruturas supramoleculares para o seu uso en biotecnoloxía.[207]
Virus de bacterias
[editar | editar a fonte]- Artigo principal: Bacteriófago.

Os bacteriófagos son un grupo de virus común e diverso e son a forma máis abundante de entidade biolóxica en medios acuáticos: hai ata dez veces máis deste tipo de virus nos océanos do que bacterias,[208] atinxindo niveis de 250.000.000 de bacteriófagos por mililitro de auga do mar.[209] Estes virus infectan a bacterias específicas uníndose a moléculas receptoras de superficie e despois entrando na célula. Nun breve período de tempo, nalgúns casos só minutos, empeza a traducirse o ARNm viral a proteínas. Estas proteínas orixinan novos virións dentro da célula, proteínas auxiliares, que axudan a ensamblar novos virións, ou proteínas implicadas na lise da célula. Os encimas virais axudan na degradación da membrana da célula, e, no caso do fago T4, en só uns vinte minutos despois da inxección libéranse uns 300 fagos.[210]
O principal modo que utilizan as bacterias para defenderse dos bacteriófagos é producir encimas que destrúen o ADN alleo. Estes encimas, chamados endonucleases de restrición, cortan en anacos o ADN viral que os bacteriófagos inxectan nas bacterias.[211] As bacterias tamén dispoñen dun sistema que utiliza as secuencias CRISPR para reter fragmentos do xenoma dos virus con que as bacterias estiveron en contacto no pasado, o que lles permite bloquear a replicación dos virus por medio dun mecanismo similar á interferencia de ARN.[212][213] Este sistema xenético proporciona ás bacterias inmunidade adquirida ás infeccións.
Virus de arqueas
[editar | editar a fonte]Algúns virus replícanse dentro de arqueas: estes son virus de ADN de dobre cadea con formas raras e ás veces únicas.[6][77] Estes virus foron estudados en maior detalle en arqueas termófilas, particularmente as ordes dos Sulfolobales e Thermoproteales.[214] A defensa contra estes virus poden facela por medio de interferencia de ARN a partir de secuencias de ADN repetitivo dos xenomas das arqueas que están relacionados cos xenes dos virus.[215][216]
Papel dos virus en ecosistemas acuáticos
[editar | editar a fonte]Unha culleradiña de auga de mar contén arredor dun millón de virus.[217] Son esenciais na regulación dos ecosistemas de auga salgada e doce.[218] A maioría destes virus son bacteriófagos, polo que son inofensivos para plantas e animais. Estes virus infectan e destrúen bacterias nas comunidades microbianas acuáticas, as cales interveñen no mecanismo máis importante de reciclaxe do carbono nos medios mariños. As moléculas orgánicas liberadas polas células bacterianas por acción dos virus estimulan o crecemento de novas bacterias e algas.[219]
Os microorganismos constitúen máis do 90% da biomasa do mar. Estímase que os virus matan aproximadamente o 20% desta biomasa cada día e que hai 15 veces máis virus nos océanos ca bacterias e arqueas. Os virus son os principais axentes responsables da rápida destrución das nocivas floracións de algas (entre elas as mareas vermellas),[220] que poden matar ou intoxicar a outras formas de vida mariñas.[221] O número de virus nos océanos decrece canto máis nos afastamos da costa e canto maior é a profundidade, onde hai menos organismos hóspedes.[222]
Os efectos dos virus mariños son de grande alcance. Ao incrementaren a cantidade de fotosíntese nos océanos cambiando a composición das comunidades de seres vivos mariñas, os virus son indirectamente responsables da redución do dióxido de carbono atmosférico en aproximadamente 3 xigatoneladas de carbono por ano.[222]
Como calquera outros organismos, os mamíferos mariños son susceptibles ás infeccións virais por virus de animais. En 1988 e 2002, morreron en Europa miles de focas comúns (Phoca vitulina) a causa do virus do moquillo das focas (un paramixovirus do xénero Morbilivirus).[223] Moitos outros virus, como calicivirus, herpesvirus, adenovirus e parvovirus, circulan entre as poboacións de mamíferos mariños.[222]
Papel dos virus na evolución
[editar | editar a fonte]- Artigo principal: Transferencia horizontal de xenes.
Os virus son un medio natural importante de transferencia de xenes entre diferentes especies, o cal incrementa a diversidade xenética e impulsa a evolución.[224] Pénsase que os virus tiveron un papel central nos inicios da evolución na Terra, antes da diversificación de bacterias, arqueas e eucariotas e no tempo en que viviu o último antepasado universal común dos seres vivos da Terra.[225] Os virus son aínda hoxe un dos maiores reservorios de diversidade xenética inexplorada na Terra.[222]
Aplicacións
[editar | editar a fonte]Ciencias da vida e medicina
[editar | editar a fonte]
Os virus son importantes nas investigacións de bioloxía molecular e bioloxía celular, xa que proporcionan sistemas simples que poden usarse para manipular e estudar as funcións da célula.[226][227] Por exemplo, os virus foron moi útiles no estudo da xenética e axudaron a comprender os mecanismos básicos de xenética molecular, como a replicación do ADN, transcrición, procesamento do ARN, tradución, transporte de proteínas e inmunoloxía.
Os xenetistas a miúdo utilizan os virus como vectores para introducir xenes nas células que están a estudar. Isto é útil para facer que a célula produza unha substancia allea, ou para estudar o efecto de introducir un novo xene no xenoma. De modo similar, a viroterapia utiliza os virus como vectores para tratar varias doenzas, xa que poden unirse especificamente a células e ao ADN. É moi prometedor o seu uso no tratamento do cancro e na terapia xénica. Científicos da Europa do leste utilizaron contra as infeccións bacterianas a terapia de fagos como unha alternativa aos antibióticos durante algún tempo, e o interese por esta estratexia está incrementándose, debido ao crecente nivel de resistencia a antibióticos que están presentando algunhas bacterias patóxenas.[228] A expresión de proteínas heterólogas polos virus é a base de varios procesos de produción actuais de proteínas como as dos antíxenos de vacinas e anticorpos. Desenvolvéronse recentemente procesos industriais que usan vectores virais, e varias proteínas farmacolóxicas están en proceso de ensaio preclínico ou clínico.[229]
Ciencias dos materiais e nanotecnoloxía
[editar | editar a fonte]As tendencias actuais en nanotecnoloxía pretenden facer un uso máis versátil dos virus. Desde o punto de vista dun científico dos materiais, os virus poden verse como nanopartículas orgánicas. A súa superficie leva instrumentos específicos deseñados para cruzar as barreiras das súas células hóspede. O tamaño e forma dos virus, e o número e natureza dos grupos funcionais da súa superficie, está definido con precisión. Os virus utilízanse comunmente en ciencias dos materiais como armazóns para modificacións de superficies unidas covalentemente. Unha calidade especial dos virus é que poden ser adaptados por evolución dirixida. As poderosas técnicas desenvolvidas polas ciencias da vida están converténdose na base de estratexias de enxeñaría para obter nanomateriais, o que abre un amplo campo de aplicacións alén da bioloxía e medicina.[230]
Debido ao seu tamaño, forma e estrutura química ben definida, os virus utilizáronse como moldes para organizar materiais a nanoescala. Exemplos recentes inclúen traballos no Laboratorio de Investigación Naval dos Estados Unidos, nos que se utilizaron partículas do virus do mosaico do feixón caupí (Vigna unguiculata), ou CPMV, para amplificar os sinais de sensores baseados en micromatrices de ADN. Nesta aplicación, as partículas do virus separan a tinguidura fluorescente utilizada para a sinalización para impedir a formación de dímeros non fluorescentes que actúen como atenuadores da fluorescencia.[231] Outro exemplo é o uso de CPMV como taboleiros de circuítos a nanoescala na electrónica molecular.[232]
Virus sintéticos
[editar | editar a fonte]Moitos virus poden sintetizarse de novo e o primeiro virus sintético creouse en 2002.[233] Na realidade, non é o verdadeiro virus o que se sintetiza, senón só o seu xenoma de ADN (no caso dos virus de ADN), ou unha copia de ADNc do seu xenoma (no caso dos virus de ARN). Para moitas familias de virus o ADN ou ARN sintético espido (unha vez que foi convertido encimaticamente en ADNc sintético) é infeccioso se o introducimos nunha célula. É dicir, o virus contén toda a información precisa para producir novos virus. Esta tecnoloxía está agora usándose para investigar novas estratexias de produción de vacinas.[234] A capacidade de sintetizar virus ten consecuencias de enorme alcance, xa que un virus xa non se pode considerar extinguido, con tal que se coñeza e preserve a información da secuencia do seu xenoma e se dispoña de células permisivas para ese virus. Actualmente, pode dispoñerse publicamente en bases de datos en liña dos Institutos Nacionais da Saúde dos EEUU das secuencias xenómicas completas de 2.408 virus distintos (incluíndo o da varíola).[235]
Armas biolóxicas
[editar | editar a fonte]A capacidade dos virus de causar epidemias devastadoras nas sociedades humanas trouxo a preocupación de que os virus se utilizasen como armas na guerra biolóxica. A preocupación aumentou cando se conseguiu volver a producir con éxito no laboratorio o virus da gripe de 1918 de sinistra sona.[236] O virus da varíola foi historicamente unha praga en moitas áreas antes da súa erradicación. Hai polo menos dous centros coñecidos no mundo que gardan mostras do virus da varíola: o Instituto Vector en Rusia e os Centros para a Prevención e Control de Enfermidades nos EEUU.[237] Os temores de que poidan usarse como armas microbiolóxicas non están totalmente infundados[237] e o virus da varíola podería ser unha arma temible. Téñase en conta que a vacina da varíola tiña unha porcentaxe de casos con efectos secundarios graves (durante os últimos anos da vacinación antes da erradicación, cando xa había poucos casos de varíola, houbo máis casos de xente seriamente enferma como resultado da vacinación ca de mortes pola varíola),[238] e a vacinación contra a varíola xa non se practica universalmente.[239] En consecuencia, gran parte da poboación humana actual case non adquiriu ningunha resistencia á varíola.[237]
Notas
[editar | editar a fonte]- ↑ 1,0 1,1 Koonin EV, Senkevich TG, Dolja VV (2006). "The ancient Virus World and evolution of cells". Biol. Direct 1: 29. PMC 1594570. doi:10.1186/1745-6150-1-29. 16984643. Arquivado dende o orixinal o 04 de decembro de 2012. Consultado o 1 de outubro de 2013.
- ↑ 2,0 2,1 Casjens S (2010). Mahy BWJ and Van Regenmortel MHV, ed. Desk Encyclopedia of General Virology. Boston: Academic Press. p. 167. ISBN 0-12-375146-2.
- ↑ 3,0 3,1 3,2 (Dimmock, Easton & Leppard 2007, p. 4)
- ↑ 4,0 4,1 4,2 (Dimmock, Easton & Leppard 2007, p. 49)
- ↑ 5,0 5,1 Breitbart M, Rohwer F (2005). "Here a virus, there a virus, everywhere the same virus?". Trends Microbiol 13 (6): 278–84. doi:10.1016/j.tim.2005.04.003. 15936660. Arquivado dende o orixinal o 04 de outubro de 2013. Consultado o 1 de outubro de 2013.
- ↑ 6,0 6,1 Lawrence CM, Menon S, Eilers BJ; et al. (2009). "Structural and functional studies of archaeal viruses". J. Biol. Chem. 284 (19): 12599–603. PMC 2675988. PMID 19158076. doi:10.1074/jbc.R800078200.
- ↑ Edwards RA, Rohwer F (2005). "Viral metagenomics". Nat. Rev. Microbiol. 3 (6): 504–10. doi:10.1038/nrmicro1163. 15886693. Consultado o 1 de outubro de 2013.
- ↑ Canchaya C, Fournous G, Chibani-Chennoufi S, Dillmann ML, Brüssow H. Phage as agents of lateral gene transfer. Curr. Opin. Microbiol.. 2003;6(4):417–24. doi:10.1016/S1369-5274(03)00086-9. PMID 12941415.
- ↑ 9,0 9,1 Rybicki, EP (1990). "The classification of organisms at the edge of life, or problems with virus systematics". S Afr J Sci 86: 182–186.
- ↑ (Shors 2008, pp. 49–50)
- ↑ Harper D (2011). "virus". The Online Etymology Dictionary. Consultado o 14 maio 2013.
- ↑ Bordenave G (2003). "Louis Pasteur (1822–1895)". Microbes and Infection / Institut Pasteur 5 (6): 553–60. PMID 12758285. doi:10.1016/S1286-4579(03)00075-3.
- ↑ (Shors 2008, pp. 76–77)
- ↑ 14,0 14,1 14,2 (Collier & Mahy 1998, p. 3)
- ↑ (Dimmock, Easton & Leppard 2007, pp. 4–5)
- ↑ Fenner F. (2009). Mahy B. W. J. and Van Regenmortal M. H. V., ed. Desk Encyclopedia of General Virology (1 ed.). Oxford, UK: Academic Press. pp. 15. ISBN 0-12-375146-2.
- ↑ (Shors 2008, p. 589)
- ↑ D'Herelle F (2007). "On an invisible microbe antagonistic toward dysenteric bacilli: brief note by Mr. F. D'Herelle, presented by Mr. Roux". Research in Microbiology 158 (7): 553–4. PMID 17855060. doi:10.1016/j.resmic.2007.07.005.
- ↑ Steinhardt E, Israeli C, Lambert R.A. (1913). "Studies on the cultivation of the virus of vaccinia". J. Inf Dis. 13 (2): 294–300. doi:10.1093/infdis/13.2.294.
- ↑ (Collier & Mahy 1998, p. 4)
- ↑ Goodpasture EW, Woodruff AM, Buddingh GJ (1931). "The cultivation of vaccine and other viruses in the chorioallantoic membrane of chick embryos". Science 74 (1919): 371–372. PMID 17810781. doi:10.1126/science.74.1919.371.
- ↑ Rosen, FS (2004). "Isolation of poliovirus—John Enders and the Nobel Prize". New England Journal of Medicine 351 (15): 1481–83. PMID 15470207. doi:10.1056/NEJMp048202.
- ↑ From Nobel Lectures, Physics 1981–1990, (1993) Editor-in-Charge Tore Frängsmyr, Editor Gösta Ekspång, World Scientific Publishing Co., Singapore.
- En 1887, Buist visualizou un dos virus máis grandes, o virus vaccinia, por microscopía óptica despois de tinguilo. Daquela non se sabía que a vaccinia era un virus. (Buist J.B. Vaccinia and Variola: a study of their life history Churchill, London)
- ↑ Stanley WM, Loring HS (1936). "The isolation of crystalline tobacco mosaic virus protein from diseased tomato plants". Science 83 (2143): 85. PMID 17756690. doi:10.1126/science.83.2143.85.
- ↑ Stanley WM, Lauffer MA (1939). "Disintegration of tobacco mosaic virus in urea solutions". Science 89 (2311): 345–347. PMID 17788438. doi:10.1126/science.89.2311.345.
- ↑ Creager AN, Morgan GJ (2008). "After the double helix: Rosalind Franklin's research on Tobacco mosaic virus". Isis 99 (2): 239–72. PMID 18702397. doi:10.1086/588626.
- ↑ (Dimmock, Easton & Leppard 2007, p. 12)
- ↑ Norrby E (2008). "Nobel Prizes and the emerging virus concept". Arch. Virol. 153 (6): 1109–23. PMID 18446425. doi:10.1007/s00705-008-0088-8.
- ↑ (Collier & Mahy 1998, p. 745)
- ↑ 30,0 30,1 Temin HM, Baltimore D (1972). "RNA-directed DNA synthesis and RNA tumor viruses". Adv. Virus Res. 17: 129–86. PMID 4348509. doi:10.1016/S0065-3527(08)60749-6.
- ↑ Barré-Sinoussi, F.; et al. (1983). "Isolation of a T-lymphotropic retrovirus from a patient at risk for acquired immune deficiency syndrome (AIDS)". Science 220 (4599): 868–871. PMID 6189183. doi:10.1126/science.6189183.
- ↑ Iyer LM, Balaji S, Koonin EV, Aravind L (2006). "Evolutionary genomics of nucleo-cytoplasmic large DNA viruses". Virus Res. 117 (1): 156–84. PMID 16494962. doi:10.1016/j.virusres.2006.01.009.
- ↑ 33,0 33,1 Sanjuán R, Nebot MR, Chirico N, Mansky LM, Belshaw R (2010). "Viral mutation rates". Journal of Virology 84 (19): 9733–48. PMC 2937809. PMID 20660197. doi:10.1128/JVI.00694-10.
- ↑ (Shors 2008, pp. 14–16)
- ↑ (Collier & Mahy 1998, pp. 11–21)
- ↑ 36,0 36,1 (Dimmock, Easton & Leppard 2007, p. 16)
- ↑ (Collier & Mahy 1998, p. 11)
- ↑ 38,0 38,1 38,2 38,3 Mahy WJ & Van Regenmortel MHV (eds) (2009). "Desk Encyclopedia of General Virology". Oxford: Academic Press: 24. ISBN 0-12-375146-2.
- ↑ (Shors 2008, p. 574)
- ↑ McClintock, B. (1950). "The origin and behavior of mutable loci in maize". Proc Natl Acad Sci U S A. 36 (6): 344–55. PMC 1063197. PMID 15430309. doi:10.1073/pnas.36.6.344.
- ↑ (Collier & Mahy 1998, pp. 11–12)
- ↑ (Dimmock, Easton & Leppard 2007, p. 55)
- ↑ (Shors 2008, pp. 551–553)
- ↑ Tsagris EM, de Alba AE, Gozmanova M, Kalantidis K (2008). "Viroids". Cell. Microbiol. 10 (11): 2168. PMID 18764915. doi:10.1111/j.1462-5822.2008.01231.x.
- ↑ (Shors 2008, pp. 492–493)
- ↑ 46,0 46,1 La Scola B, Desnues C, Pagnier I, Robert C, Barrassi L, Fournous G, Merchat M, Suzan-Monti M, Forterre P, Koonin E, Raoult D (2008). "The virophage as a unique parasite of the giant mimivirus". Nature 455 (7209): 100–4. PMID 18690211. doi:10.1038/nature07218.
- ↑ (Collier & Mahy 1998, p. 777)
- ↑ (Dimmock, Easton & Leppard 2007, pp. 55–57)
- ↑ 49,0 49,1 Mahy WJ & Van Regenmortel MHV (eds) (2009). "Desk Encyclopedia of General Virology". Oxford: Academic Press: 28. ISBN 0-12-375146-2.
- ↑ 50,0 50,1 Mahy WJ & Van Regenmortel MHV (eds) (2009). "Desk Encyclopedia of General Virology". Oxford: Academic Press: 26. ISBN 0-12-375146-2.
- ↑ (Dimmock, Easton & Leppard 2007, pp. 15–16)
- ↑ Institut de Génétique et Microbiologie (2006). "The origin of viruses and their possible roles in major evolutionary transitions.". Virus Res. 5 (117): 5–16. PMID 16476498. Consultado o 24 maio 2013.
- ↑ Koonin EV, Senkevich TG, Dolja VV. (2006). "The ancient Virus World and evolution of cells.". PUBMED I: 29. PMID 16984643. Consultado o 24 maio 2013.
- ↑ Liberski PP (2008). "Prion diseases: a riddle wrapped in a mystery inside an enigma". Folia Neuropathol 46 (2): 93–116. PMID 18587704.
- ↑ Belay ED and Schonberger LB (2009). "Desk Encyclopedia of Human and Medical Virology". Boston: Academic Press: 497–504. ISBN 0-12-375147-0.
- ↑ Lupi O, Dadalti P, Cruz E, Goodheart C (2007). "Did the first virus self-assemble from self-replicating prion proteins and RNA?". Med. Hypotheses 69 (4): 724–30. PMID 17512677. doi:10.1016/j.mehy.2007.03.031.
- ↑ Holmes EC (2007). "Viral evolution in the genomic age". PLoS Biol. 5 (10): e278. PMC 1994994. PMID 17914905. doi:10.1371/journal.pbio.0050278.
- ↑ Wimmer E, Mueller S, Tumpey TM, Taubenberger JK (2009). "Synthetic viruses: a new opportunity to understand and prevent viral disease". Nature Biotechnology 27 (12): 1163–72. PMC 2819212. PMID 20010599. doi:10.1038/nbt.1593.
- ↑ Horn M (2008). "Chlamydiae as symbionts in eukaryotes". Annual Review of Microbiology 62: 113–31. PMID 18473699. doi:10.1146/annurev.micro.62.081307.162818.
- ↑ Ammerman NC, Beier-Sexton M, Azad AF (2008). "Laboratory maintenance of Rickettsia rickettsii". Current Protocols in Microbiology. Chapter 3: Unit 3A.5. PMC 2725428. PMID 19016440. doi:10.1002/9780471729259.mc03a05s11.
- ↑ 61,0 61,1 (Collier & Mahy 1998, pp. 33–55)
- ↑ (Collier & Mahy 1998, p. 33–37)
- ↑ Kiselev NA, Sherman MB, Tsuprun VL (1990). "Negative staining of proteins". Electron Microsc. Rev. 3 (1): 43–72. PMID 1715774. doi:10.1016/0892-0354(90)90013-I.
- ↑ (Collier & Mahy 1998, p. 40)
- ↑ Caspar DL, Klug A (1962). "Physical principles in the construction of regular viruses". Cold Spring Harb. Symp. Quant. Biol. 27: 1–24. PMID 14019094.
- ↑ Crick FH, Watson JD (1956). "Structure of small viruses". Nature 177 (4506): 473–5. PMID 13309339. doi:10.1038/177473a0.
- ↑ Falvo, M.R.; Washburn, S.; Superfine, R.; Finch, M.; Brooks Jr, F.P.; Chi, V.; Taylor 2nd, R.M. (1997). "Manipulation of individual viruses: friction and mechanical properties". Biophysical Journal 72 (3): 1396–1403. PMC 1184522. PMID 9138585. doi:10.1016/S0006-3495(97)78786-1.
- ↑ Kuznetsov, Yu. G.; Malkin, A. J.; Lucas,, R. W.; Plomp, M.; McPherson, A. (2001). "Imaging of viruses by atomic force microscopy". J Gen Virol 82 (9): 2025–2034. PMID 11514711.
- ↑ (Collier & Mahy 1998, p. 37)
- ↑ (Collier & Mahy 1998, pp. 40, 42)
- ↑ Casens, S. (2009). Desk Encyclopedia of General Virology. Boston: Academic Press. pp. 167–174. ISBN 0-12-375146-2.
- ↑ (Collier & Mahy 1998, p. 42–43)
- ↑ Rossmann MG, Mesyanzhinov VV, Arisaka F, Leiman PG (2004). "The bacteriophage T4 DNA injection machine". Curr. Opin. Struct. Biol. 14 (2): 171–80. PMID 15093831. doi:10.1016/j.sbi.2004.02.001.
- ↑ Long GW, Nobel J, Murphy FA, Herrmann KL, Lourie B (1970). "Experience with electron microscopy in the differential diagnosis of smallpox". Appl Microbiol 20 (3): 497–504. PMC 376966. PMID 4322005.
- ↑ Suzan-Monti M, La Scola B, Raoult D (2006). "Genomic and evolutionary aspects of Mimivirus". Virus Research 117 (1): 145–155. PMID 16181700. doi:10.1016/j.virusres.2005.07.011.
- ↑ "World’s biggest virus discovered in ocean depths near Chile". Consultado o 07 maio 2013.
- ↑ 77,0 77,1 Prangishvili D, Forterre P, Garrett RA (2006). "Viruses of the Archaea: a unifying view". Nat. Rev. Microbiol. 4 (11): 837–48. PMID 17041631. doi:10.1038/nrmicro1527.
- ↑ 78,0 78,1 78,2 (Collier & Mahy 1998, pp. 96–99)
- ↑ Saunders, Venetia A.; Carter, John (2007). Virology: principles and applications. Chichester: John Wiley & Sons. pp. 72. ISBN 0-470-02387-2.
- ↑ Van Etten JL, Lane LC, Dunigan DD (2010). "DNA viruses: the really big ones (giruses)". Annual Review of Microbiology 64: 83–99. PMC 2936810. PMID 20690825. doi:10.1146/annurev.micro.112408.134338.
- ↑ Pressing J, Reanney DC (1984). "Divided genomes and intrinsic noise". J Mol Evol 20 (2): 135–46. PMID 6433032. doi:10.1007/BF02257374.
- ↑ Duffy S, Holmes EC (2009). "Validation of high rates of nucleotide substitution in geminiviruses: phylogenetic evidence from East African cassava mosaic viruses". The Journal of General Virology 90 (Pt 6): 1539–47. PMID 19264617. doi:10.1099/vir.0.009266-0.
- ↑ Sandbulte MR, Westgeest KB, Gao J, Xu X, Klimov AI, Russell CA, Burke DF, Smith DJ, Fouchier RA, Eichelberger MC. Discordant antigenic drift of neuraminidase and hemagglutinin in H1N1 and H3N2 influenza viruses. Proceedings of the National Academy of Sciences of the United States of America. 2011;108(51):20748–53. doi:10.1073/pnas.1113801108. PMID 22143798.
- ↑ Moss RB, Davey RT, Steigbigel RT, Fang F. Targeting pandemic influenza: a primer on influenza antivirals and drug resistance. The Journal of Antimicrobial Chemotherapy. 2010;65(6):1086–93. doi:10.1093/jac/dkq100. PMID 20375034.
- ↑ Hampson AW, Mackenzie JS (2006). "The influenza viruses". Med. J. Aust. 185 (10 Suppl): S39–43. PMID 17115950.
- ↑ Metzner KJ (2006). "Detection and significance of minority quasispecies of drug-resistant HIV-1". J HIV Ther 11 (4): 74–81. PMID 17578210.
- ↑ Goudsmit, Jaap. Viral Sex. Oxford Univ Press, 1998.ISBN 978-0-19-512496-5 ISBN 0-19-512496-0
- ↑ Worobey M, Holmes EC (1999). "Evolutionary aspects of recombination in RNA viruses". J. Gen. Virol. 80 ( Pt 10): 2535–43. PMID 10573145.
- ↑ Lukashev AN (2005). "Role of recombination in evolution of enteroviruses". Rev. Med. Virol. 15 (3): 157–67. PMID 15578739. doi:10.1002/rmv.457.
- ↑ Umene K (1999). "Mechanism and application of genetic recombination in herpesviruses". Rev. Med. Virol. 9 (3): 171–82. PMID 10479778. doi:10.1002/(SICI)1099-1654(199907/09)9:3<171::AID-RMV243>3.0.CO;2-A.
- ↑ (Collier & Mahy 1998, pp. 75–91)
- ↑ (Dimmock, Easton & Leppard 2007, p. 70)
- ↑ Boevink P, Oparka KJ (2005). "Virus-host interactions during movement processes". Plant Physiol. 138 (4): 1815–21. PMC 1183373. PMID 16172094. doi:10.1104/pp.105.066761.[Ligazón morta]
- ↑ (Dimmock, Easton & Leppard 2007, p. 71)
- ↑ Barman S, Ali A, Hui EK, Adhikary L, Nayak DP (2001). "Transport of viral proteins to the apical membranes and interaction of matrix protein with glycoproteins in the assembly of influenza viruses". Virus Res. 77 (1): 61–9. PMID 11451488. doi:10.1016/S0168-1702(01)00266-0.
- ↑ (Shors 2008, pp. 60, 597)
- ↑ Dimmock, Chapter 15, Mechanisms in virus latentcy, pp.243–259
- ↑ (Dimmock, Easton & Leppard 2007, pp. 185–187)
- ↑ Rico Garófalo, Xavier (2019). "Los tipos de virus: cómo funcionan y enfermedades asociadas". Med Salud. Arquivado dende o orixinal o 24 de xuño de 2019. Consultado o 24 de xuño de 2019.
- ↑ (Shors 2008, p. 54); (Collier & Mahy 1998, p. 78)
- ↑ (Collier & Mahy 1998, p. 79)
- ↑ Staginnus C, Richert-Pöggeler KR (2006). "Endogenous pararetroviruses: two-faced travelers in the plant genome". Trends in Plant Science 11 (10): 485–91. PMID 16949329. doi:10.1016/j.tplants.2006.08.008.
- ↑ (Collier & Mahy 1998, pp. 88–89)
- ↑ (Collier & Mahy 1998, pp. 115–146)
- ↑ (Collier & Mahy 1998, p. 115)
- ↑ Roulston A, Marcellus RC, Branton PE (1999). "Viruses and apoptosis". Annu. Rev. Microbiol. 53: 577–628. PMID 10547702. doi:10.1146/annurev.micro.53.1.577.
- ↑ Alwine JC (2008). "Modulation of host cell stress responses by human cytomegalovirus". Curr. Top. Microbiol. Immunol. 325: 263–79. PMID 18637511. doi:10.1007/978-3-540-77349-8_15.
- ↑ Sinclair J (2008). "Human cytomegalovirus: Latency and reactivation in the myeloid lineage". J. Clin. Virol. 41 (3): 180–5. PMID 18164651. doi:10.1016/j.jcv.2007.11.014.
- ↑ Jordan MC, Jordan GW, Stevens JG, Miller G (1984). "Latent herpesviruses of humans". Ann. Intern. Med. 100 (6): 866–80. PMID 6326635.
- ↑ Sissons JG, Bain M, Wills MR (2002). "Latency and reactivation of human cytomegalovirus". J. Infect. 44 (2): 73–7. PMID 12076064. doi:10.1053/jinf.2001.0948.
- ↑ Barozzi P, Potenza L, Riva G, Vallerini D, Quadrelli C, Bosco R, Forghieri F, Torelli G, Luppi M (2007). "B cells and herpesviruses: a model of lymphoproliferation". Autoimmun Rev 7 (2): 132–6. PMID 18035323. doi:10.1016/j.autrev.2007.02.018.
- ↑ Subramanya D, Grivas PD (2008). "HPV and cervical cancer: updates on an established relationship". Postgrad Med 120 (4): 7–13. PMID 19020360. doi:10.3810/pgm.2008.11.1928.
- ↑ Crawford, Dorothy H. (2011). Viruses: A Very Short Introduction. Oxford University Press, USA. p. 16. ISBN 0-19-957485-5.
- ↑ (Shors 2008, p. 388)
- ↑ (Shors 2008, p. 353)
- ↑ (Dimmock, Easton & Leppard 2007, p. 272)
- ↑ Baggesen DL, Sørensen G, Nielsen EM, Wegener HC (2010). "Phage typing of Salmonella Typhimurium – is it still a useful tool for surveillance and outbreak investigation?". Eurosurveillance 15 (4): 19471. PMID 20122382.
- ↑ Lwoff A, Horne RW, Tournier P (1962). "A virus system". C. R. Hebd. Seances Acad. Sci. (en French) 254: 4225–7. PMID 14467544.
- ↑ Lwoff A, Horne R, Tournier P (1962). "A system of viruses". Cold Spring Harb. Symp. Quant. Biol. 27: 51–5. PMID 13931895.
- ↑ 120,0 120,1 King AMQ, Lefkowitz E, Adams MJ, Carstens EB (2011). Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses. Elsevier. p. 6. ISBN 0-12-384684-6.
- ↑ Adams, MJ.; Lefkowitz, EJ.; King, AM.; Carstens, EB. (2013). "Recently agreed changes to the International Code of Virus Classification and Nomenclature.". Archives of Virology 158 (12): 2633–9. PMID 23836393. doi:10.1007/s00705-013-1749-9.
- "Unha especie é un grupo monofilético de virus cuxas propiedades poden ser distinguidas das doutras especies por múltiples criterios."
- ↑ Delwart EL (2007). "Viral metagenomics". Rev. Med. Virol. 17 (2): 115–31. PMID 17295196. doi:10.1002/rmv.532.
- ↑ Virus Taxonomy 2009. Arquivado 23 de xullo de 2011 en Wayback Machine. International Committee on Taxonomy of Viruses. Retrieved on December 24, 2011.
- ↑ ICTV Master Species List 2009 Arquivado 05 de marzo de 2016 en Wayback Machine.
- Este ficheiro contén a lista oficial de Especies Mestras do ICTV de 2009, e inclúe unha relación de todos os taxons de virus aprobados. Esta é a versión 10 do MSL publcada o 24 de agosto de 2011.
- ↑ Baltimore D (1974). "The strategy of RNA viruses". Harvey Lect. 70 Series: 57–74. PMID 4377923.
- ↑ van Regenmortel MH, Mahy BW (2004). "Emerging issues in virus taxonomy". Emerging Infect. Dis. 10 (1): 8–13. PMID 15078590.
- ↑ Mayo MA (1999). "Developments in plant virus taxonomy since the publication of the 6th ICTV Report. International Committee on Taxonomy of Viruses". Arch. Virol. 144 (8): 1659–66. PMID 10486120. doi:10.1007/s007050050620.
- ↑ de Villiers EM, Fauquet C, Broker TR, Bernard HU, zur Hausen H (2004). "Classification of papillomaviruses". Virology 324 (1): 17–27. PMID 15183049. doi:10.1016/j.virol.2004.03.033.
- ↑ Góra-Sochacka A. (2004). "Viroids: unusual small pathogenic RNAs.". Acta Biochim Pol. (51(3)): 587–607. PMID 15448723.
- ↑ Roossinck MJ, Sleat D, Palukaitis P. (1992). "Satellite RNAs of plant viruses: structures and biological effects.". Microbiol Rev. (xuño;56(2)): 265–279. PMID 1620065.
- ↑ Symons RH. (1991). "The intriguing viroids and virusoids: what is their information content and how did they evolve?". Mol Plant Microbe Interact. (Mar-Apr;4(2)): 111–121. PMID 1932808.
- ↑ Bernard La Scola, Christelle Desnues, Isabelle Pagnier, Catherine Robert, Lina Barrassi, Ghislain Fournous, Michèle Merchat, Marie Suzan-Monti, Patrick Forterre, Eugene Koonin and Didier Raoult (2008). "The virophage as a unique parasite of the giant mimivirus". Nature 455 (7205): 100. PMID 18690211. doi:10.1038/nature07218.
- ↑ Head MW1, Ironside JW. (2012). "Review: Creutzfeldt-Jakob disease: prion protein type, disease phenotype and agent strain.". Neuropathol Appl Neurobiol. (xuño;38(4)): 296–310. PMID 22394291.
- ↑ Ivan N. Zheludev; Robert C. Edgar; Maria Jose Lopez-Galiano; Marcos de la Peña; Artem Babaian; Ami S. Bhatt; Andrew Z. Fire (21 de xaneiro de 2024). "Viroid-like colonists of human microbiomes".
- ↑ Mainly Chapter 33 (Disease summaries), pages 367–392 in:Fisher, Bruce; Harvey, Richard P.; Champe, Pamela C. (2007). Lippincott's Illustrated Reviews: Microbiology (Lippincott's Illustrated Reviews Series). Hagerstwon, MD: Lippincott Williams & Wilkins. pp. pages 367–392. ISBN 0-7817-8215-5.
- ↑ Para o arrefriado común: National Institute of Allergy and Infectious Diseases (NIAID) - Common Cold. posta ao día por última vez en decembro de 2007.
- ↑ Komaroff AL (2006). "Is human herpesvirus-6 a trigger for chronic fatigue syndrome?". J. Clin. Virol. 37 Suppl 1: S39–46. PMID 17276367. doi:10.1016/S1386-6532(06)70010-5.
- ↑ Chen C, Chiu Y, Wei F, Koong F, Liu H, Shaw C, Hwu H, Hsiao K (1999). "High seroprevalence of Borna virus infection in schizophrenic patients, family members and mental health workers in Taiwan". Mol Psychiatry 4 (1): 33–8. PMID 10089006. doi:10.1038/sj.mp.4000484.
- ↑ Margolis TP, Elfman FL, Leib D; et al. (2007). "Spontaneous reactivation of herpes simplex virus type 1 in latently infected murine sensory ganglia". J. Virol. 81 (20): 11069–74. PMC 2045564. PMID 17686862. doi:10.1128/JVI.00243-07.
- ↑ Whitley RJ, Roizman B (2001). "Herpes simplex virus infections". Lancet 357 (9267): 1513–8. PMID 11377626. doi:10.1016/S0140-6736(00)04638-9.
- ↑ Barton ES, White DW, Cathelyn JS; et al. (2007). "Herpesvirus latency confers symbiotic protection from bacterial infection". Nature 447 (7142): 326–9. PMID 17507983. doi:10.1038/nature05762.
- ↑ Bertoletti A, Gehring A (2007). "Immune response and tolerance during chronic hepatitis B virus infection". Hepatol. Res. 37 Suppl 3: S331–8. PMID 17931183. doi:10.1111/j.1872-034X.2007.00221.x.
- ↑ Rodrigues C, Deshmukh M, Jacob T, Nukala R, Menon S, Mehta A (2001). "Significance of HBV DNA by PCR over serological markers of HBV in acute and chronic patients". Indian journal of medical microbiology 19 (3): 141–4. PMID 17664817.
- ↑ Nguyen VT, McLaws ML, Dore GJ (2007). "Highly endemic hepatitis B infection in rural Vietnam". Journal of Gastroenterology and Hepatology 22 (12): 2093–100. PMID 17645465. doi:10.1111/j.1440-1746.2007.05010.x.
- ↑ Fowler MG, Lampe MA, Jamieson DJ, Kourtis AP, Rogers MF (2007). "Reducing the risk of mother-to-child human immunodeficiency virus transmission: past successes, current progress and challenges, and future directions". Am. J. Obstet. Gynecol. 197 (3 Suppl): S3–9. PMID 17825648. doi:10.1016/j.ajog.2007.06.048.
- ↑ Sauerbrei A, Wutzler P (2000). "The congenital varicella syndrome". Journal of perinatology : official journal of the California Perinatal Association 20 (8 Pt 1): 548–54. PMID 11190597.
- ↑ Garnett GP (2005). "Role of herd immunity in determining the effect of vaccines against sexually transmitted disease". J. Infect. Dis. 191 Suppl 1: S97–106. PMID 15627236. doi:10.1086/425271.
- ↑ Platonov AE (2006). "(The influence of weather conditions on the epidemiology of vector-borne diseases by the example of West Nile fever in Russia)". Vestn. Akad. Med. Nauk SSSR (en Russian) (2): 25–9. PMID 16544901.
- ↑ (Shors 2008, p. 198)
- ↑ (Shors 2008, p. 199, 209)
- ↑ (Shors 2008, p. 19)
- ↑ (Shors 2008, p. 126)
- ↑ 153,0 153,1 (Shors 2008, pp. 193–194)
- ↑ (Shors 2008, p. 194)
- ↑ (Shors 2008, pp. 192–193)
- ↑
- Ranlet P (2000). "The British, the Indians, and smallpox: what actually happened at Fort Pitt in 1763?". Pa Hist 67 (3): 427–41. PMID 17216901.
- Van Rijn K (2006). ""Lo! The poor Indian!" colonial responses to the 1862–63 smallpox epidemic in British Columbia and Vancouver Island". Can Bull Med Hist 23 (2): 541–60. PMID 17214129.
- Patterson KB, Runge T (2002). "Smallpox and the Native American". Am. J. Med. Sci. 323 (4): 216–22. PMID 12003378. doi:10.1097/00000441-200204000-00009.
- Sessa R, Palagiano C, Scifoni MG, di Pietro M, Del Piano M (1999). "The major epidemic infections: a gift from the Old World to the New?". Panminerva Med 41 (1): 78–84. PMID 10230264.
- Bianchine PJ, Russo TA (1992). "The role of epidemic infectious diseases in the discovery of America". Allergy Proc 13 (5): 225–32. PMID 1483570. doi:10.2500/108854192778817040. Consultado o 16 September 2008.
- Hauptman LM (1979). "Smallpox and American Indian; Depopulation in Colonial New York". N Y State J Med 79 (12): 1945–9. PMID 390434.
- Fortuine R (1988). "Smallpox decimates the Tlingit (1787)". Alaska Med 30 (3): 109. PMID 3041871.
- ↑ (Collier & Mahy 1998, pp. 409–415)
- ↑ Patterson KD, Pyle GF (1991). "The geography and mortality of the 1918 influenza pandemic". Bull Hist Med. 65 (1): 4–21. PMID 2021692.
- ↑ Johnson NP, Mueller J (2002). "Updating the accounts: global mortality of the 1918–1920 "Spanish" influenza pandemic". Bull Hist Med 76 (1): 105–15. PMID 11875246. doi:10.1353/bhm.2002.0022.
- ↑ Gao F, Bailes E, Robertson DL; et al. (1999). "Origin of HIV-1 in the Chimpanzee Pan troglodytes troglodytes". Nature 397 (6718): 436–441. PMID 9989410. doi:10.1038/17130.
- ↑ (Shors 2008, p. 447)
- ↑ Mawar N, Saha S, Pandit A, Mahajan U (2005). "The third phase of HIV pandemic: social consequences of HIV/AIDS stigma & discrimination & future needs" (PDF). Indian J. Med. Res. 122 (6): 471–84. PMID 16517997. Arquivado dende o orixinal (PDF) o 04 de marzo de 2016. Consultado o 13 September 2008.
- ↑ "Status of the global HIV epidemic" (PDF). UNAIDS. 2008. Arquivado dende o orixinal (PDF) o 22 de novembro de 2015. Consultado o 15 September 2008.
- ↑ Towner JS, Khristova ML, Sealy TK; et al. (2006). "Marburgvirus genomics and association with a large hemorrhagic fever outbreak in Angola". J. Virol. 80 (13): 6497–516. PMC 1488971. PMID 16775337. doi:10.1128/JVI.00069-06.
- ↑ Einstein MH, Schiller JT, Viscidi RP, Strickler HD, Coursaget P, Tan T, Halsey N, Jenkins D (2009). "Clinician's guide to human papillomavirus immunology: knowns and unknowns". The Lancet Infectious Diseases 9 (6): 347–56. PMID 19467474. doi:10.1016/S1473-3099(09)70108-2.
- ↑ Shuda M, Feng H, Kwun HJ, Rosen ST, Gjoerup O, Moore PS, Chang Y (2008). "T antigen mutations are a human tumor-specific signature for Merkel cell polyomavirus". Proceedings of the National Academy of Sciences of the United States of America 105 (42): 16272–7. PMC 2551627. PMID 18812503. doi:10.1073/pnas.0806526105.
- ↑ Pulitzer MP, Amin BD, Busam KJ (2009). "Merkel cell carcinoma: review". Advances in Anatomic Pathology 16 (3): 135–44. PMID 19395876. doi:10.1097/PAP.0b013e3181a12f5a.
- ↑ Koike K (2007). "Hepatitis C virus contributes to hepatocarcinogenesis by modulating metabolic and intracellular signalling pathways". J. Gastroenterol. Hepatol. 22 Suppl 1: S108–11. PMID 17567457. doi:10.1111/j.1440-1746.2006.04669.x.
- ↑ Hu J, Ludgate L (2007). "HIV-HBV and HIV-HCV coinfection and liver cancer development". Cancer Treat. Res. 133: 241–52. doi:10.1007/978-0-387-46816-7_9.
- ↑ Bellon M, Nicot C (2007). "Telomerase: a crucial player in HTLV-I-induced human T-cell leukemia". Cancer genomics & proteomics 4 (1): 21–5. PMID 17726237.
- ↑ Schiffman M, Castle PE, Jeronimo J, Rodriguez AC, Wacholder S (2007). "Human papillomavirus and cervical cancer". Lancet 370 (9590): 890–907. PMID 17826171. doi:10.1016/S0140-6736(07)61416-0.
- ↑ Klein E, Kis LL, Klein G (2007). "Epstein-Barr virus infection in humans: from harmless to life endangering virus-lymphocyte interactions". Oncogene 26 (9): 1297–305. PMID 17322915. doi:10.1038/sj.onc.1210240.
- ↑ zur Hausen H (2008). "Novel human polyomaviruses—re-emergence of a well known virus family as possible human carcinogens". International Journal of Cancer. Journal International Du Cancer 123 (2): 247–50. PMID 18449881. doi:10.1002/ijc.23620.
- ↑ Alberts, Bruce; Johnson, Alexander; Lewis, Julian; Raff, Martin; Roberts, Keith; Walters, Peter (2002). Molecular Biology of the Cell; Fourth Edition. New York and London: Garland Science. ISBN 0-8153-3218-1. Consultado o 15 September 2008.
- ↑ Ding SW, Voinnet O (2007). "Antiviral immunity directed by small RNAs". Cell 130 (3): 413–26. PMC 2703654. PMID 17693253. doi:10.1016/j.cell.2007.07.039.
- ↑ Patton JT, Vasquez-Del Carpio R, Spencer E (2004). "Replication and transcription of the rotavirus genome". Curr. Pharm. Des. 10 (30): 3769–77. PMID 15579070. doi:10.2174/1381612043382620.
- ↑ Jayaram H, Estes MK, Prasad BV (2004). "Emerging themes in rotavirus cell entry, genome organization, transcription and replication". Virus Res. 101 (1): 67–81. PMID 15010218. doi:10.1016/j.virusres.2003.12.007.
- ↑ Greer S, Alexander GJ (1995). "Viral serology and detection". Baillieres Clin. Gastroenterol. 9 (4): 689–721. PMID 8903801. doi:10.1016/0950-3528(95)90057-8.
- ↑ Matter L, Kogelschatz K, Germann D (1997). "Serum levels of rubella virus antibodies indicating immunity: response to vaccination of subjects with low or undetectable antibody concentrations". J. Infect. Dis. 175 (4): 749–55. PMID 9086126. doi:10.1086/513967.
- ↑ Mallery DL, McEwan WA, Bidgood SR, Towers GJ, Johnson CM, James LC (2010). "Antibodies mediate intracellular immunity through tripartite motif-containing 21 (TRIM21)". Proceedings of the National Academy of Sciences of the United States of America 107 (46): 19985–90. PMC 2993423. PMID 21045130. doi:10.1073/pnas.1014074107.
- ↑ Cascalho M, Platt JL (2007). "Novel functions of B cells". Crit. Rev. Immunol. 27 (2): 141–51. PMID 17725500.
- ↑ Le Page C, Génin P, Baines MG, Hiscott J (2000). "Interferon activation and innate immunity". Rev Immunogenet 2 (3): 374–86. PMID 11256746.
- ↑ Robert F. Siliciano ,Warner C. Greene. HIV Latency Cold Spring Harb Perspect Med September 2011;1:a007096doi:10.1101/cshperspect.a007096. [1] Cita: "En estado latente, o virus persiste simplemente como información (en forma de ADN de VIH-1 integrado de 10 kb), e así non é afectado polos fármacos antirretrovirais ou as respostas inmunes" ("In the latent state, the virus persists simply as information (in the form of 10 kb of integrated HIV-1 DNA), and it is thus unaffected by antiretroviral drugs or immune responses.")
- ↑ Hilleman MR (2004). "Strategies and mechanisms for host and pathogen survival in acute and persistent viral infections". Proc. Natl. Acad. Sci. U.S.A. 101 Suppl 2: 14560–6. PMC 521982. PMID 15297608. doi:10.1073/pnas.0404758101.
- ↑ Asaria P, MacMahon E (2006). "Measles in the United Kingdom: can we eradicate it by 2010?". BMJ 333 (7574): 890–5. PMC 1626346. PMID 17068034. doi:10.1136/bmj.38989.445845.7C.
- ↑ Lane JM (2006). "Mass vaccination and surveillance/containment in the eradication of smallpox". Curr. Top. Microbiol. Immunol. 304: 17–29. PMID 16989262. doi:10.1007/3-540-36583-4_2.
- ↑ Arvin AM, Greenberg HB (2006). "New viral vaccines". Virology 344 (1): 240–9. PMID 16364754. doi:10.1016/j.virol.2005.09.057.
- ↑ Pastoret PP, Schudel AA, Lombard M (2007). "Conclusions—future trends in veterinary vaccinology". Rev. – Off. Int. Epizoot. 26 (2): 489–94, 495–501, 503–9. PMID 17892169.
- ↑ Palese P (2006). "Making better influenza virus vaccines?". Emerging Infect. Dis. 12 (1): 61–5. PMID 16494719. doi:10.3201/eid1201.051043.
- ↑ Thomssen R (1975). "Live attenuated versus killed virus vaccines". Monographs in allergy 9: 155–76. PMID 1090805.
- ↑ McLean AA (1986). "Development of vaccines against hepatitis A and hepatitis B". Rev. Infect. Dis. 8 (4): 591–8. PMID 3018891.
- ↑ Casswall TH, Fischler B (2005). "Vaccination of the immunocompromised child". Expert review of vaccines 4 (5): 725–38. PMID 16221073. doi:10.1586/14760584.4.5.725.
- ↑ Barnett ED, Wilder-Smith A, Wilson ME (2008). "Yellow fever vaccines and international travelers". Expert Rev Vaccines 7 (5): 579–87. PMID 18564013. doi:10.1586/14760584.7.5.579.
- ↑ Magden J, Kääriäinen L, Ahola T (2005). "Inhibitors of virus replication: recent developments and prospects". Appl. Microbiol. Biotechnol. 66 (6): 612–21. PMID 15592828. doi:10.1007/s00253-004-1783-3.
- ↑ Mindel A, Sutherland S (1983). "Genital herpes — the disease and its treatment including intravenous acyclovir". J. Antimicrob. Chemother. 12 Suppl B: 51–9. PMID 6355051.
- ↑ Witthöft T, Möller B, Wiedmann KH; et al. (2007). "Safety, tolerability and efficacy of peginterferon alpha-2a and ribavirin in chronic hepatitis C in clinical practice: The German Open Safety Trial". J. Viral Hepat. 14 (11): 788–96. PMC 2156112. PMID 17927615. doi:10.1111/j.1365-2893.2007.00871.x.
- ↑ Rudin D, Shah SM, Kiss A, Wetz RV, Sottile VM (2007). "Interferon and lamivudine vs. interferon for hepatitis B e antigen-positive hepatitis B treatment: meta-analysis of randomized controlled trials". Liver Int. 27 (9): 1185–93. PMC 2156150. PMID 17919229. doi:10.1111/j.1478-3231.2007.01580.x.
- ↑ (Dimmock, Easton & Leppard 2007, p. 3)
- ↑ Goris N, Vandenbussche F, De Clercq K (2008). "Potential of antiviral therapy and prophylaxis for controlling RNA viral infections of livestock". Antiviral Res. 78 (1): 170–8. PMID 18035428. doi:10.1016/j.antiviral.2007.10.003.
- ↑ Carmichael L (2005). "An annotated historical account of canine parvovirus". J. Vet. Med. B Infect. Dis. Vet. Public Health 52 (7–8): 303–11. PMID 16316389. doi:10.1111/j.1439-0450.2005.00868.x.
- ↑ Chen YP, Zhao Y, Hammond J, Hsu H, Evans JD, Feldlaufer MF (October–November 2004). "Multiple virus infections in the honey bee and genome divergence of honey bee viruses". Journal of Invertebrate Pathology 87 (2–3): 84–93. PMID 15579317. doi:10.1016/j.jip.2004.07.005.
- ↑ (Shors 2008, p. 584)
- ↑ (Shors 2008, p. 562–587)
- ↑ Dinesh-Kumar SP, Tham Wai-Hong, Baker BJ (2000). "Structure—function analysis of the tobacco mosaic virus resistance gene N". PNAS 97 (26): 14789–94. PMC 18997. PMID 11121079. doi:10.1073/pnas.97.26.14789.
- ↑ (Shors 2008, p. 573–576)
- ↑ Soosaar JL, Burch-Smith TM, Dinesh-Kumar SP (2005). "Mechanisms of plant resistance to viruses". Nat. Rev. Microbiol 3 (10): 789–98. PMID 16132037. doi:10.1038/nrmicro1239.
- ↑ Lomonossoff, GP (2011). "Virus Particles and the Uses of Such Particles in Bio- and Nanotechnology". Recent Advances in Plant Virology. Caister Academic Press. ISBN 978-1-904455-75-2.
- ↑ Wommack KE, Colwell RR (2000). "Virioplankton: viruses in aquatic ecosystems". Microbiol. Mol. Biol. Rev. 64 (1): 69–114. PMC 98987. PMID 10704475. doi:10.1128/MMBR.64.1.69-114.2000.
- ↑ Bergh O, Børsheim KY, Bratbak G, Heldal M (1989). "High abundance of viruses found in aquatic environments". Nature 340 (6233): 467–8. PMID 2755508. doi:10.1038/340467a0.
- ↑ (Shors 2008, pp. 595–97)
- ↑ Bickle TA, Krüger DH (1 June 1993). "Biology of DNA restriction". Microbiol. Rev. 57 (2): 434–50. PMC 372918. PMID 8336674.
- ↑ Barrangou R, Fremaux C, Deveau H; et al. (2007). "CRISPR provides acquired resistance against viruses in prokaryotes". Science 315 (5819): 1709–12. PMID 17379808. doi:10.1126/science.1138140.
- ↑ Brouns SJ, Jore MM, Lundgren M; et al. (2008). "Small CRISPR RNAs guide antiviral defense in prokaryotes". Science 321 (5891): 960–4. PMID 18703739. doi:10.1126/science.1159689.
- ↑ Prangishvili D, Garrett RA (2004). "Exceptionally diverse morphotypes and genomes of crenarchaeal hyperthermophilic viruses". Biochem. Soc. Trans. 32 (Pt 2): 204–8. PMID 15046572. doi:10.1042/BST0320204.
- ↑ Mojica FJ, Díez-Villaseñor C, García-Martínez J, Soria E (2005). "Intervening sequences of regularly spaced prokaryotic repeats derive from foreign genetic elements". J. Mol. Evol. 60 (2): 174–82. PMID 15791728. doi:10.1007/s00239-004-0046-3.
- ↑ Makarova KS, Grishin NV, Shabalina SA, Wolf YI, Koonin EV (2006). "A putative RNA-interference-based immune system in prokaryotes: computational analysis of the predicted enzymatic machinery, functional analogies with eukaryotic RNAi, and hypothetical mechanisms of action". Biol. Direct 1: 7. PMC 1462988. PMID 16545108. doi:10.1186/1745-6150-1-7. Arquivado dende o orixinal o 10 de maio de 2013. Consultado o 05 de maio de 2013.
- ↑ (Shors 2008, p. 4)
- ↑ (Shors 2008, p. 5)
- ↑ (Shors 2008, p. 593)
- ↑ Suttle CA (2005). "Viruses in the sea". Nature 437 (7057): 356–61. PMID 16163346. doi:10.1038/nature04160.
- ↑ "Harmful Algal Blooms: Red Tide: Home". www.cdc.gov. CDC HSB. Consultado o 23 October 2009.
- ↑ 222,0 222,1 222,2 222,3 Suttle CA (2007). "Marine viruses—major players in the global ecosystem". Nature Reviews. Microbiology 5 (10): 801–12. PMID 17853907. doi:10.1038/nrmicro1750.
- ↑ Hall, A. J., Jepson, P. D., Goodman, S. J. & Harkonen, T. "Phocine distemper virus in the North and European Seas — data and models, nature and nurture". Biol. Conserv. 131, 221–229 (2006)
- ↑ Canchaya C, Fournous G, Chibani-Chennoufi S, Dillmann ML, Brüssow H (2003). "Phage as agents of lateral gene transfer". Curr. Opin. Microbiol. 6 (4): 417–24. PMID 12941415. doi:10.1016/S1369-5274(03)00086-9.
- ↑ Forterre P, Philippe H (1999). "The last universal common ancestor (LUCA), simple or complex?". The Biological Bulletin 196 (3): 373–5; discussion 375–7. PMID 11536914. doi:10.2307/1542973.
- ↑ (Collier & Mahy 1998, p. 8)
- ↑ Lodish, Harvey; Berk, Arnold; Zipursky, S. Lawrence; Matsudaira, Paul; Baltimore, David; Darnell, James.Viruses:Structure, Function, and Uses Retrieved on September 16, 2008
- ↑ Matsuzaki S, Rashel M, Uchiyama J, Sakurai S, Ujihara T, Kuroda M, Ikeuchi M, Tani T, Fujieda M, Wakiguchi H, Imai S (2005). "Bacteriophage therapy: a revitalized therapy against bacterial infectious diseases". Journal of Infection and Chemotherapy : Official Journal of the Japan Society of Chemotherapy 11 (5): 211–9. PMID 16258815. doi:10.1007/s10156-005-0408-9.
- ↑ Gleba, YY; Giritch, A (2011). "Plant Viral Vectors for Protein Expression". Recent Advances in Plant Virology. Caister Academic Press. ISBN 978-1-904455-75-2.
- ↑ Fischlechner M, Donath E (2007). "Viruses as Building Blocks for Materials and Devices". Angewandte Chemie International Edition 46 (18): 3184–93. PMID 17348058. doi:10.1002/anie.200603445.
- ↑ Soto CM, Blum AS, Vora GJ; et al. (2006). "Fluorescent signal amplification of carbocyanine dyes using engineered viral nanoparticles". J. Am. Chem. Soc. 128 (15): 5184–9. PMID 16608355. doi:10.1021/ja058574x.
- ↑ Blum AS, Soto CM, Wilson CD; et al. (2005). "An Engineered Virus as a Scaffold for Three-Dimensional Self-Assembly on the Nanoscale". Small 7: 702. PMID 17193509. doi:10.1002/smll.200500021.
- ↑ Cello J, Paul AV, Wimmer E (2002). "Chemical synthesis of poliovirus cDNA: generation of infectious virus in the absence of natural template". Science 297 (5583): 1016–8. PMID 12114528. doi:10.1126/science.1072266.
- ↑ Coleman JR, Papamichail D, Skiena S, Futcher B, Wimmer E, Mueller S (2008). "Virus attenuation by genome-scale changes in codon pair bias". Science 320 (5884): 1784–7. PMC 2754401. PMID 18583614. doi:10.1126/science.1155761.
- ↑ Genomes. "NIH viral genome database". Ncbi.nlm.nih.gov. Consultado o 2012-05-07.
- ↑ (Shors 2008, p. 331)
- ↑ 237,0 237,1 237,2 Artenstein AW, Grabenstein JD (2008). "Smallpox vaccines for biodefense: need and feasibility". Expert Review of Vaccines 7 (8): 1225–37. PMID 18844596. doi:10.1586/14760584.7.8.1225.
- ↑ Aragón TJ, Ulrich S, Fernyak S, Rutherford GW (2003). "Risks of serious complications and death from smallpox vaccination: a systematic review of the United States experience, 1963–1968". BMC public health 3: 26. PMC 194634. PMID 12911836. doi:10.1186/1471-2458-3-26.
- ↑ Weiss MM, Weiss PD, Mathisen G, Guze P (2004). "Rethinking smallpox". Clin. Infect. Dis. 39 (11): 1668–73. PMID 15578369. doi:10.1086/425745.
Véxase tamén
[editar | editar a fonte]| Wikimedia Commons ten máis contidos multimedia na categoría: Virus |
| Vexa a entrada do Galizionario acerca de Virus |
| Wikispecies posúe unha páxina sobre: Virus |

Bibliografía
[editar | editar a fonte]- Collier, Leslie; Mahy, Brian W. J. (1998). Balows, Albert; Sussman, Max, eds. Topley and Wilson's Microbiology and Microbial Infections: Volume 1: Virology (en inglés) (9ª ed.). Arnold. ISBN 978-0340663165.
- Dimmock, N.J; Easton, Andrew J; Leppard, Keith (2007). Introduction to Modern Virology (en inglés) (6ª ed.). Blackwell Publishing. ISBN 978-1405136457.
- Knipe, David M; Howley, Peter M; Griffin, Diane E; Lamb, Robert A; Martin, Malcolm A; Roizman, Bernard; Straus Stephen E. (2007). Fields Virology (en inglés). Lippincott Williams & Wilkins. ISBN 0-7817-6060-7.
- Mahy WJ; Van Regenmortel MHV, eds. (2009). "Desk Encyclopedia of General Virology" (en inglés). Oxford: Academic Press. ISBN 0-12-375146-2.
- Shors, Teri (2008). Understanding Viruses (en inglés). Jones and Bartlett Publishers. ISBN 0-7637-2932-9.
Outros artigos
[editar | editar a fonte]Ligazóns externas
[editar | editar a fonte]- "ViralZone" (en inglés). Recursos de todas as familias virais, con información molecular e epidemiolóxica.
- Baltimore, David. "Introduction to Viruses and HIV" [Introdución aos virus e ao VIH]. iBiology (en inglés). Arquivado dende o orixinal o 29 de outubro de 2013. Consultado o 27 de xuño de 2015.
- Helenius, Ari. "Virus Entry". iBiology (en inglés). Arquivado dende o orixinal o 14 de outubro de 2011. Consultado o 05 de maio de 2013.
- "Viruses — Open Access Virology Journal" (en inglés).
- "Virus en 3 dimensións". The European Bioinformatics Institute (en inglés).