Arĥeoj
 | ||||
|---|---|---|---|---|
| Troveblo de fosilioj: Arĥaiko - Nuntempe | ||||
 | ||||
| Biologia klasado | ||||
| ||||
Aliaj Vikimediaj projektoj
| ||||
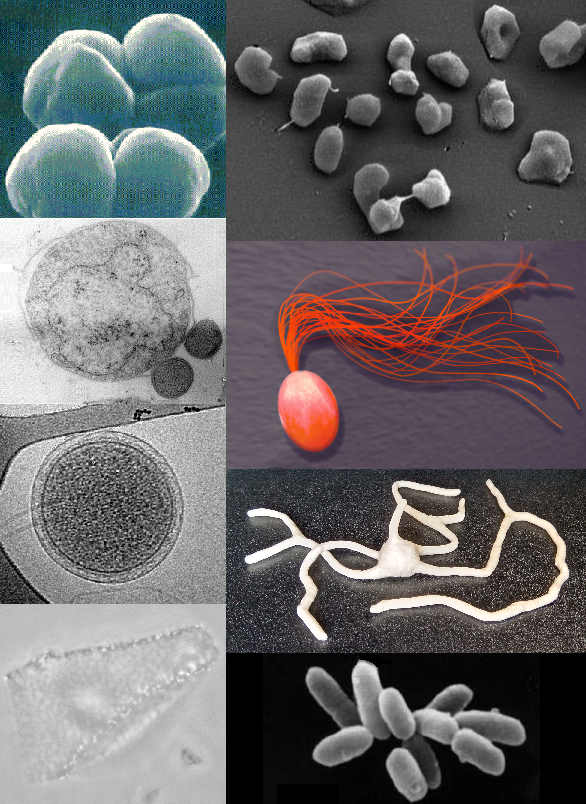
Arĥeoj aŭ Arkeoj (latine Archaea) estas unu el tri biologiaj domanioj. Arĥeoj estas unuĉelaj kaj prokariotaj estaĵoj (do ne havas ĉelkernojn), tamen ili ne estas bakterioj, kvankam ili dekomence estis klasigitaj kiel bakterioj, kaj ricevis la nomon prabakterioj (latine archaebacteria, en la Prabakteria regno) - tiu klasigo eksmodiĝis post la studoj fare de Carl Woese.[1] Sur la genetika nivelo, ili estas pli similaj al eŭkariotoj kiel ni mem. Estis Carl Woese, kiu eltrovis la arĥeojn en 1977.
Priskribo
[redakti | redakti fonton]Arĥeaj ĉeloj havas unikajn trajtojn kiuj apartigas ilin el la aliaj du vivoregnoj, nome Bakterioj kaj Eŭkariotoj. La arĥeoj estas krome subdividitaj en multajn agnoskitajn filumojn. Ilia klasigo estas malfacile farebla, ĉar plimulto el ili ankoraŭ ne estas izolita en laboratorioj kaj estas detektita nur pere de analizo de la nukleataj acidoj en montraĵoj elprenitaj el koncernaj medioj.
Arĥeoj kaj bakterioj estas ĝenerale similaj laŭ grando kaj formo, kvankam kelkaj arĥeoj havas formojn tre malsimilajn al tiuj de bakterioj, kiaj la ebenaj kaj kvadrat-formaj ĉeloj de Haloquadratum walsbyi.[2] Spite tiun morfologian similecon al la bakterio, arĥeoj posedas genojn kaj kelkajn metabolajn sistemojn kiuj estas pli proksime rilataj al tiuj de eŭkariotoj, ĉefe la enzimoj kiuj funkcias en la genetikaj procezoj de transskribado kaj tradukado. Aliaj aspektoj de arĥea biokemio estas unikaj, kiaj ilia dependo el eteraj lipidoj en iliaj ĉelmembranoj, kiel ĉe la arĥeoloj. Arĥeoj uzas pli da energifontoj ol la eŭkariotoj: tiuj gamas el organikaj komponaĵoj, kiel sukeroj, al amoniako, metalaj jonoj aŭ eĉ hidrogenhava gaso. Sal-toleraj arĥeoj (nome Haloarchaea aŭ Salarĥeoj) uzas sunlumon kiel energifonto, kaj aliaj specioj de arĥeoj fiksigas karbonon, sed malkiel ĉe plantoj kaj cianobakterioj, neniu konata specio de arĥeo plenumas ambaŭ procezojn. Arĥeoj reproduktiĝas nesekse pere de binara fisio, fragmentado, aŭ burĝonado; malkiel ĉe bakterioj kaj eŭkariotoj, neniu konata specio de arĥeo formas sporojn.

Multaj arĥeoj estas ekstremofiloj, tio estas vivas en tre akraj medioj — ekzemple, varmegaj, salegaj, kaj acidaj lagoj. Inter tiuj estis registritaj dekomence termofontoj kaj sallagoj, sed ekde tiam ili troviĝis ankaŭ en ampleksa gamo de habitatoj, kiel grundoj, oceanoj kaj marĉoj. Ili estas ankaŭ parto de la homa mikrobaro, kiu troviĝas en kojlo, buŝo kaj haŭto.[3] Arĥeoj estas partikulare multnombraj en la oceanoj, kaj la aĥeoj de la planktono povas esti el la plej abundaj grupoj de organismoj sur la planedo. Arĥeoj estas la ĉefa parto de la surtera vivo kaj ili povas ludi gravajn rolojn kaj en la karbona kaj en la nitrogena cikloj.
Oni ne konas klarajn ekzemplojn de arĥeaj patogenoj aŭ parazitoj, sed ili estas ofte mutualistoj aŭ kunmanĝantoj. Unu ekzemplo estas la metanogenoj kiuj loĝas en la digesta sistemo de homoj aŭ remaĉuloj, kie ilia multnombreco fakte helpas la digestadon. Metanogenoj estas uzataj ankaŭ en produktado de biogaso kaj en procezoj de akvopurigado, kaj bioteknologio ekspluatas enzimojn el ekstremofilaj arĥeoj kiuj povas rezisti altajn temperaturojn kaj organikajn solvigaĵojn. Laŭ nuna scio, la arĥeoj kaŭzas neniun ajn homan aŭ bestan malsanon — ili estas plej interesaj laŭ la vidpunkto de evoluo. Multaj sciencistoj nun kredas ke la komuna parencaro de ĉiuj eŭkariotoj estis simila al arĥeoj.
Klasigo
[redakti | redakti fonton]
Nova regno
[redakti | redakti fonton]Dum multo de la 20a jarcento, prokariotoj estis konsiderataj kiel aparta grupo de organismoj kaj klasigitaj baze sur sia biokemio, morfologio kaj metabolo. Por ekzemplo, mikrobiologoj klopodis klasigi mikroorganismojn baze sur la strukturoj de iliaj ĉelmuroj, ilia formoj kaj la substancojn kiujn ili konsumas.[4] En 1965, Emile Zuckerkandl kaj Linus Pauling[5] proponis anstataŭe uzi la sekvencojn de la genoj en diversaj prokariotoj por studi kiel ili estas rilataj unu al la alia. Tiu filogenetika aliro estas la ĉefa metodo uzata nuntempe.
Arĥeoj estis dekomence klasigitaj kiel aparta grupo de prokariotoj en 1977 fare de Carl Woese kaj George E. Fox baze sur la sekvencoj de ribosomaj genoj RRNA.[6] Tiuj du grupoj estis origine nomitaj Arĥeobakterioj kaj Eŭbakterioj kaj estis termigitaj Urkingdoms fare de Woese kaj Fox; aliaj esploristoj traktis ilin kiel regnoj aŭ subgenroj. En tiu publikigo Woese kaj Fox prezentis la unuajn pruvarojn por la koncepto de arĥeobakterioj - kiuj tiam enhavis nur la metanogenojn - kiel aparta "devenlinio":
- manko de peptidoglikano en iliaj ĉelmuroj,
- du maloftaj kunenzimoj,
- rezultoj de 16S ribosoma RNA gensekvencaro.

Por emfazi tiun diferencon, Woese, Otto Kandler kaj Mark Wheelis poste proponis novan naturan sistemon de organismoj kun tri apartaj domanioj: nome Eŭkariotoj, Bakterioj kaj Arĥeoj,[7] en tio kio estas nune konata kiel "La Woese-a Revolucio". La historio de la esplorado pri Arĥeoj estas dokumentita detale fare de Jan Sapp.[8]
La vorto archaea de la scienca nomo derivas el la antikva greka ἀρχαῖα, signife "antikvaj aĵoj",[9] ĉar la unuaj reprezentantoj de la domanio Archaea estis metanogenoj kaj oni supozis, ke ilia metabolo spegulis la praatmosferon de la Tero kaj la antikveco de la organismoj, sed kiam aliaj novaj habitatoj estis studitaj, pliaj organismoj estis malkovritaj. Ankaŭ ekstremaj halofiloj[10] kaj hipertermofilaj mikroboj[11] estis inkluditaj en Archaea. Dum longe, archaea estis konsideritaj kiel ekstremofiloj kiuj ekzistas nur en ekstremaj habitatoj kiaj termofontoj kaj sallagoj. Fine de la 20a jarcento, archaea estis identigitaj ankaŭ en ne-ekstremaj medioj. Nuntempe, ili estas konata kiel granda kaj diversa grupo de organismoj kiuj estas amplekse distribuataj en naturo kaj estas oftaj en ĉiuj habitatoj.[12] Tiu nova alproksimiĝo al la gravo kaj ĉieeco de la arĥeoj venis el la uzado de polimeraza ĉen-reakcio (PĈR) por detekti prokariotojn el mediaj montraĵoj (kiel akvo aŭ grundo) per de la multobligo de iliaj ribosomaj genoj. Tio ebligas la detekton kaj identigon de organismoj kiuj ne estis produktitaj en la laboratorio.[13][14]
Klasigo
[redakti | redakti fonton]
La klasigo de arĥeoj, kaj de prokariotoj ĝenerale, estas rapide moviĝanta kaj polemikiga kampo. Nuntempaj klasigsistemoj celas organizi arĥeojn en grupoj de organismoj kiuj kunhavas strukturajn trajtojn kaj komunajn praulojn.[15] Tiuj klasigoj ege dependas el la uzado de la sekvenco de ribosomaj RNA genoj por montri la rilatojn inter organismoj (molekula filogenetiko).[16] Plimulto de la kultureblaj kaj bone esploritaj specioj de arĥeoj estas membroj de du ĉefaj filumoj, nome "Euryarchaeota" kaj la Thermoproteota (iam Crenarchaeota). Aliaj grupoj estis provizore kreitaj, kiel la aparta specio Nanoarchaeum equitans, kiun oni malkovris en 2003 kaj ricevis sian propran filumon, nome "Nanoarchaeota".[17] Oni proponis ankaŭ novan filumon "Korarchaeota". Ĝi enhavas malgrandan grupon de nekutimaj termofilaj specioj kiuj kunhavas trajtojn de ambaŭ ĉefaj filumoj, sed ĝi estas pli proksime rilata al la Thermoproteota.[18][19] Aliaj detektitaj specioj de arĥeoj estas nur pli fore rilataj al iu el tiuj grupoj, kiel la grupo nomita ARMAN, enhavanta Micrarchaeota kaj Parvarchaeota, kiujn oni malkovris en 2006[20] kaj estas kelkaj el la plej malgrandaj organismoj konataj.[21]
Superfilumo – TACK – kiu inkludas Thaumarchaeota (nune Nitrososphaerota), "Aigarchaeota", Crenarchaeota (nune Thermoproteota), kaj "Korarchaeota" estis proponita en 2011 por esti rilata al la origino de eŭkariotoj.[22] En 2017, la ĵus malkovrita kaj ĵus nomita superfilumo Asgard estis proponita por esti pli proksime rilata al la originaj eŭkariotoj kaj frata grupo TACK.[23]
En 2013 oni proponis la superfilumon DPANN por la grupoj "Nanoarchaeota", "Nanohaloarchaeota", ARMAN (enhavanta "Micrarchaeota" kaj "Parvarchaeota"), kaj aliajn similajn arĥeojn. Tiu arĥea superfiluma enhavas almenaŭ 10 diferencajn stirpojn kaj inkludas organismojn kun tre malgrandaj ĉelaj kaj genaran grandojn kaj limigitan metabolajn kapablojn. Tiel, multaj membroj de DPANN povus esti devige dependantaj de simbioza interagado kun aliaj organismoj kaj povas eĉ inkludi novmalkovrotajn parazitojn. Tamen, laŭ aliaj filogenetikaj analizoj oni trovis, ke DPANN ne formas monofiletikan grupon kaj ke tion okazigas la longa branĉa altiro (LBA), sugestante, ke ĉiuj tiuj stirpoj apartenas al "Euryarchaeota".[24][25]
Koncepto de specio
[redakti | redakti fonton]
Ankaŭ la klasigo de arĥeoj en specioj estas polemikiga. Ernst Mayr difinis specion kiel grupo de interreproduktaj organismoj kiuj estas reprodukte izolaj, sed tio ne helpas ĉar arĥeoj reproduktiĝas nur sensekse.[26]
Arĥeoj montras altajn nivelojn de "horizontala gentransigo" inter stirpoj. Kelkaj esploristoj sugestas, ke oni povas unuope grupigi en speci-ecaj populacioj pro la tre altaj similaj genaroj kaj neofta gentransigo al/el ĉeloj kun malpli rilataj genaroj, ĉar en la genro Ferroplasma.[27] Aliflanke, studoj pri Halorubrum trovis gravan genetikan transigon al/el malpli rilataj populacioj, limiganta la aplikeblon de kriterioj.[28] Kelkaj esploristoj pridubas ĉu tiaj specinomigoj havas praktikan signifon.[29]
Nuntempa sciaro pri genetika diverseco en arĥeoj estas fragmenta, kaj tial la totala nombro de specioj povas esti ĉirkaŭkalkulita per neniu akurateco.[16] Ĉirkaŭkalkuloj de la nombro de filumoj gamas el 18 ĝis 23, de kiuj nur 8 havas reprezentantojn kiujn oni kultivis kaj studis rekte. Multaj el tiuj hipotezaj grupoj estas konataj el ununura sekvenco de rRNA, indikante, ke la diverseco inter tiuj organismoj restas neklara.[30] La taksono Bacteria inkludas ankaŭ multajn nekultivitajn mikrobojn kun similaj karakteroj.[31]
Filumoj
[redakti | redakti fonton]Validaj filumoj
[redakti | redakti fonton]La jenaj filumoj estis valide publikigitaj laŭ la Bakteriologia Kodo:[32]
- Nitrososphaerota
- Thermoproteota
Provizoraj filumoj
[redakti | redakti fonton]Oni proponis jenajn filumojn, sed ne estis valide publikigitaj laŭ la Bakteriologia Kodo (inklude tiujn kiuj havas statuson de candidatus (kandidato)):
- "Candidatus Aenigmarchaeota"
- "Candidatus Aigarchaeota"
- "Candidatus Altiarchaeota"
- "Candidatus Asgardaeota"
- "Candidatus Bathyarchaeota"
- "Candidatus Brockarchaeota"
- "Candidatus Diapherotrites"
- "Euryarchaeota"
- "Candidatus Geoarchaeota"
- "Candidatus Hadarchaeota"
- "Candidatus Hadesarchaeota"
- "Candidatus Halobacterota"
- "Candidatus Heimdallarchaeota"
- "Candidatus Helarchaeota"
- "Candidatus Huberarchaeota"
- "Candidatus Hydrothermarchaeota"
- "Candidatus Korarchaeota"
- "Candidatus Lokiarchaeia"
- "Candidatus Lokiarchaeota"
- "Candidatus Mamarchaeota"
- "Candidatus Marsarchaeota"
- "Candidatus Methanobacteriota"
- "Candidatus Micrarchaeota"
- "Candidatus Nanoarchaeota"
- "Candidatus Nanohaloarchaeota"
- "Candidatus Nezhaarchaeota"
- "Candidatus Odinarchaeota"
- "Candidatus Pacearchaeota"
- "Candidatus Parvarchaeota"
- "Candidatus Thermoplasmatota"
- "Candidatus Thorarchaeota"
- "Candidatus Undinarchaeota"
- "Candidatus Verstraetearchaeota"
- "Candidatus Woesearchaeota"
Origino kaj evolucio
[redakti | redakti fonton]
La Aĝo de la Tero estas ĉirkaŭ 4.54 mil milionoj da jaroj.[33][34][35] Scienca pruvaro sugestas, ke la vivo startis en la Tero almenaŭ antaŭ 3.5 mil milionoj da jaroj.[36][37] La plej fruaj pruvoj de vivo en la Tero estas grafito trovita kiel biogeniko en 3.7-mil milion-jaraj metasedimentaj rokaĵoj malkovritaj en Okcidenta Gronlando[38] kaj mikrobtavolaj fosilioj trovitaj en 3.48-mil milion-jara sabloŝtono malkovrita en Okcidenta Aŭstralio.[39][40] En 2015, eblaj restaĵoj de biotika materialo estis trovitaj en 4.1-mil milion-jaraj rokaĵoj en Okcidenta Aŭstralio.[41][42]
Kvankam probablaj prokariotaj ĉel-fosilioj datas de antaŭ preskaŭ 3.5 mil milionoj da jaroj, plej prokariotoj ne havas distingajn morfologiojn, kaj fosiliaj formoj ne povas esti uzataj por identigi ilin kiel arĥeoj.[43] Anstataŭe, kemiaj fosilioj de unikaj lipidoj estas pli informak ĉar tiuj komponantoj ne ekzistas en aliaj organismoj.[44] Kelkaj publikaĵoj sugestas, ke arĥeaj aŭ eukariotaj lipidoj restas en argilecaj tavolrokoj datitaj el 2.7 mil milionoj da jaroj,[45] kvankam tiuj datumoj ekde tiam estis pridubitaj.[46] Tiuj lipidoj estis detektitaj ankaŭ en eĉ pli aĝaj rokoj el Okcidenta Gronlando. La plej aĝa el tiu retaĵoj venas el la distrikto Isua, kiu inkludas la plej antikvajn konatajn sedimentojn de la Tero, formitaj antaŭ 3.8 mil milionoj da jaroj.[47] La arĥea stirpo eble estas la plej antikva kiu ekzistas en la Tero.[48]
Woese argumentis, ke Bacteria, Archaea, kaj Eukaryotes estas apartaj linioj de descendi kiu diverĝis frue el prakolonio de organismoj.[49][50] Une eblo[50][51] estas, ke tio okazis antaŭ la evoluo de ĉeloj, kiam la manko de tipa ĉelmembrano ebligis senrestriktan flankan gentransigon, kaj ke la komunaj prauloj de la tri domajnoj aperis per fiksado de specifaj subaroj de genoj.[50][51] Eble la lasta komuna praulo de bakterioj kaj arĥeoj estis termofila, kio starigas la eblon ke pli malaltaj temperaturoj estis "ekstremaj medioj" por arĥeoj, kaj ke organismoj kiuj vivas en pli malvarmaj medioj aperis nur poste.[52] Ĉar arĥeoj kaj bakterioj jam ne estas plu rilataj unuj al aliaj ol ili estas al eŭkariotoj, la termino prokarioto povas sugesti falsan similecon inter ili.[53] Tamen, strukturaj kaj funkciaj similaĵoj inter stirpoj ofte ekzistas pro kunhavitaj praulaj trajtoj aŭ evolua konverĝo. Tiuj similaĵoj estas konataj kiel grado, kaj oni komprenas prokariotoj plej bone kiel grado de vivo, karakterizita per trajtoj kiel foresto de membran-ligitaj organetoj.
Referencoj
[redakti | redakti fonton]- ↑ Pace NR (May 2006). "Time for a change". Nature. 441 (7091): 289. Bibcode:2006Natur.441..289P. doi:10.1038/441289a. PMID 16710401.
- ↑ Stoeckenius W (Oktobro 1981). "Walsby's square bacterium: fine structure of an orthogonal procaryote". Journal of Bacteriology. 148 (1): 352–60. PMC 216199 Libere alirebla, alirita la 22an de Julio 2018. PMID 7287626.
- ↑ Bang C, Schmitz RA (Septembro 2015). "Archaea associated with human surfaces: not to be underestimated". FEMS Microbiology Reviews. 39 (5): 631–48. doi:10.1093/femsre/fuv010. PMID 25907112.
- ↑ (November 2006) “The bacterial species dilemma and the genomic-phylogenetic species concept”, Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 361 (1475), p. 1899–909. doi:10.1098/rstb.2006.1914.
- ↑ (Marto 1965) “Molecules as documents of evolutionary history”, Journal of Theoretical Biology 8 (2), p. 357–66. doi:10.1016/0022-5193(65)90083-4.
- ↑ Woese CR, Fox GE (November 1977). "Phylogenetic structure of the prokaryotic domain: the primary kingdoms". Proceedings of the National Academy of Sciences of the United States of America. 74 (11): 5088–90. Bibcode:1977PNAS...74.5088W. doi:10.1073/pnas.74.11.5088. PMC 432104 Libere alirebla. Alirita la 23an de Julio 2018. PMID 270744.
- ↑ (June 1990) “Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya”, Proceedings of the National Academy of Sciences of the United States of America 87 (12), p. 4576–9. doi:10.1073/pnas.87.12.4576. Bibkodo:1990PNAS...87.4576W.
- ↑ Sapp, Jan. (2009) The new foundations of evolution: on the tree of life. Nov-Jorko: Oxford University Press. ISBN 978-0-199-73438-2.
- ↑ Archaea. (2008). En Merriam-Webster Online Dictionary. Alirita la 23an de Julio, 2018
- ↑ Magrum LJ, Luehrsen KR, Woese CR (May 1978). "Are extreme halophiles actually "bacteria"?". Journal of Molecular Evolution. 11 (1): 1–8. Bibcode:1978JMolE..11....1M. doi:10.1007/bf01768019. PMID 660662.
- ↑ Stetter KO (1996). "Hyperthermophiles in the history of life". Ciba Foundation Symposium. 202: 1–10; discussion 11–8. PMID 9243007.
- ↑ DeLong EF (December 1998). "Everything in moderation: archaea as 'non-extremophiles'". Current Opinion in Genetics & Development. 8 (6): 649–54. doi:10.1016/S0959-437X(98)80032-4. PMID 9914204.
- ↑ Theron J, Cloete TE (2000). "Molecular techniques for determining microbial diversity and community structure in natural environments". Critical Reviews in Microbiology. 26 (1): 37–57. doi:10.1080/10408410091154174. PMID 10782339.
- ↑ Schmidt TM (Septembro 2006). "The maturing of microbial ecology" (PDF). International Microbiology. 9 (3): 217–23. PMID 17061212. Arkivita el la originalo (PDF) en 11a de Septembro 2008.
- ↑ (Novembro 2006) “Stepping stones towards a new prokaryotic taxonomy”, Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 361 (1475), p. 1911–16. doi:10.1098/rstb.2006.1915.
- ↑ 16,0 16,1 (December 2005) “Phylogenetic diversity and ecology of environmental Archaea”, Current Opinion in Microbiology 8 (6), p. 638–642. doi:10.1016/j.mib.2005.10.003.
- ↑ (May 2002) “A new phylum of Archaea represented by a nanosized hyperthermophilic symbiont”, Nature 417 (6884), p. 63–67. doi:10.1038/417063a. Bibkodo:2002Natur.417...63H. 4395094.
- ↑ (August 1996) “Perspectives on archaeal diversity, thermophily and monophyly from environmental rRNA sequences”, Proceedings of the National Academy of Sciences of the United States of America 93 (17), p. 9188–93. doi:10.1073/pnas.93.17.9188. Bibkodo:1996PNAS...93.9188B.
- ↑ (June 2008) “A korarchaeal genome reveals insights into the evolution of the Archaea”, Proceedings of the National Academy of Sciences of the United States of America 105 (23), p. 8102–07. doi:10.1073/pnas.0801980105. Bibkodo:2008PNAS..105.8102E.
- ↑ (Decembro 2006) “Lineages of acidophilic archaea revealed by community genomic analysis”, Science 314 (5807), p. 1933–35. doi:10.1126/science.1132690. Bibkodo:2006Sci...314.1933B. 26033384.
- ↑ (Majo 2010) “Enigmatic, ultrasmall, uncultivated Archaea”, Proceedings of the National Academy of Sciences of the United States of America 107 (19), p. 8806–11. doi:10.1073/pnas.0914470107. Bibkodo:2010PNAS..107.8806B.
- ↑ (Decembro 2011) “The archaeal 'TACK' superphylum and the origin of eukaryotes”, Trends in Microbiology 19 (12), p. 580–87. doi:10.1016/j.tim.2011.09.002.
- ↑ (Januaro 2017) “Asgard archaea illuminate the origin of eukaryotic cellular complexity”, Nature 541 (7637), p. 353–58. doi:10.1038/nature21031. Bibkodo:2017Natur.541..353Z. 4458094.
- ↑ Nina Dombrowski, Jun-Hoe Lee, Tom A Williams, Pierre Offre, Anja Spang (2019). Genomic diversity, lifestyles and evolutionary origins of DPANN archaea. Nature.
- ↑ Petitjean C, Deschamps P, López-García P, Moreira D (Decembro 2014). "Rooting the domain archaea by phylogenomic analysis supports the foundation of the new kingdom Proteoarchaeota". Genome Biology and Evolution. 7 (1): 191–204. doi:10.1093/gbe/evu274. PMC 4316627. PMID 25527841.
- ↑ (Majo 2005) “Ernst Mayr and the modern concept of species”, Proceedings of the National Academy of Sciences of the United States of America 102 (Supplement 1), p. 6600–6007. doi:10.1073/pnas.0502030102. Bibkodo:2005PNAS..102.6600D.
- ↑ (Septembro 2007) “Genetic exchange across a species boundary in the archaeal genus ferroplasma”, Genetics 177 (1), p. 407–16. doi:10.1534/genetics.107.072892.
- ↑ (Aŭgusto 2007) “Searching for species in haloarchaea”, Proceedings of the National Academy of Sciences of the United States of America 104 (35), p. 14092–97. doi:10.1073/pnas.0706358104. Bibkodo:2007PNAS..10414092P.
- ↑ (July 2005) “The net of life: reconstructing the microbial phylogenetic network”, Genome Research 15 (7), p. 954–59. doi:10.1101/gr.3666505.
- ↑ (2002) “Exploring prokaryotic diversity in the genomic era”, Genome Biology 3 (2), p. REVIEWS0003. doi:10.1186/gb-2002-3-2-reviews0003.
- ↑ (2003) “The uncultured microbial majority”, Annual Review of Microbiology 57, p. 369–94. doi:10.1146/annurev.micro.57.030502.090759. 10781051. Arkivigite je 2019-03-02 per la retarkivo Wayback Machine
- ↑ (2021) “Valid publication of the names of forty-two phyla of prokaryotes”, Int J Syst Evol Microbiol 71 (10), p. 5056. doi:10.1099/ijsem.0.005056. 239887308.
- ↑ Age of the Earth. U.S. Geological Survey (1997). Arkivita el la originalo je 23a de Decembro 2005. Alirita 2006-01-10.
- ↑ (2001) “The age of the Earth in the twentieth century: a problem (mostly) solved”, Special Publications, Geological Society of London 190 (1), p. 205–21. doi:10.1144/GSL.SP.2001.190.01.14. Bibkodo:2001GSLSP.190..205D. 130092094.
- ↑ (1980) “Lead isotope study of basic-ultrabasic layered complexes: Speculations about the age of the earth and primitive mantle characteristics”, Earth and Planetary Science Letters 47 (3), p. 370–82. doi:10.1016/0012-821X(80)90024-2. Bibkodo:1980E&PSL..47..370M.
- ↑ (Oktobro 1995) “The Beginnings of Life on Earth”, American Scientist. Alirita 15a de Januaro 2014.. Arkivigite je 2017-06-06 per la retarkivo Wayback Machine
- ↑ 3.5 billion year old organic deposits show signs of life. 'Ars Technica' (4a de Septembro 2012). Alirita 15a de Januaro 2014.
- ↑ (8a de Decembro 2013) “Evidence for biogenic graphite in early Archaean Isua metasedimentary rocks”, 'Nature Geoscience' 7 (1), p. 25. doi:10.1038/ngeo2025. Bibkodo:2014NatGe...7...25O.
- ↑ "Oldest fossil found: Meet your microbial mom", 13a de Novembro 2013.
- ↑ (Decembro 2013) “Microbially induced sedimentary structures recording an ancient ecosystem in the ca. 3.48 billion-year-old Dresser Formation, Pilbara, Western Australia”, Astrobiology 13 (12), p. 1103–24. doi:10.1089/ast.2013.1030. Bibkodo:2013AsBio..13.1103N.
- ↑ "Hints of life on what was thought to be desolate early Earth", 'Excite', 'Mindspark Interactive Network', 19a de Oktobro 2015.
- ↑ (Novembro 2015) “Potentially biogenic carbon preserved in a 4.1 billion-year-old zircon”, Proceedings of the National Academy of Sciences of the United States of America 112 (47), p. 14518–21. doi:10.1073/pnas.1517557112. Bibkodo:2015PNAS..11214518B.
- ↑ (Junio 2006) “Fossil evidence of Archaean life”, Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 361 (1470), p. 869–85. doi:10.1098/rstb.2006.1834.
- ↑ (Julio 1982) “Polar lipids of archaebacteria in sediments and petroleums”, Science 217 (4554), p. 65–66. doi:10.1126/science.217.4554.65. Bibkodo:1982Sci...217...65C. 42758483.
- ↑ (August 1999) “Archean molecular fossils and the early rise of eukaryotes”, Science 285 (5430), p. 1033–36. doi:10.1126/science.285.5430.1033. Bibkodo:1999Sci...285.1033B.
- ↑ (Oktobro 2008) “Reassessing the first appearance of eukaryotes and cyanobacteria”, Nature 455 (7216), p. 1101–4. doi:10.1038/nature07381. Bibkodo:2008Natur.455.1101R. 4372071.
- ↑ (1986) “Traces of Archaebacteria in ancient sediments”, System Applied Microbiology 7 (Archaebacteria '85 Proceedings), p. 178–83. doi:10.1016/S0723-2020(86)80002-9.
- ↑ (November 2007) “Reductive evolution of architectural repertoires in proteomes and the birth of the tripartite world”, Genome Research 17 (11), p. 1572–85. doi:10.1101/gr.6454307.
- ↑ (January 1981) “Are archaebacteria merely derived 'prokaryotes'?”, Nature 289 (5793), p. 95–96. doi:10.1038/289095a0. Bibkodo:1981Natur.289...95W. 4343245.
- ↑ 50,0 50,1 50,2 (June 1998) “The universal ancestor”, Proceedings of the National Academy of Sciences of the United States of America 95 (12), p. 6854–59. doi:10.1073/pnas.95.12.6854. Bibkodo:1998PNAS...95.6854W.
- ↑ 51,0 51,1 (August 1998) “The early diversification of life and the origin of the three domains: a proposal.”, Thermophiles: the keys to molecular evolution and the origin of life.. Athens: Taylor and Francis, p. 19–31. ISBN 978-1-4822-7304-5.
- ↑ (June 2006) “The origin and evolution of Archaea: a state of the art”, Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 361 (1470), p. 1007–22. doi:10.1098/rstb.2006.1841.
- ↑ (March 1994) “There must be a prokaryote somewhere: microbiology's search for itself”, Microbiological Reviews 58 (1), p. 1–9. doi:10.1128/MMBR.58.1.1-9.1994.
Literaturo
[redakti | redakti fonton]- Howland JL (2000). The Surprising Archaea: Discovering Another Domain of Life. Alirita la 22an de Julio 2018. Oxford University. ISBN 978-0-19-511183-5.
- Martinko JM, Madigan MT (2005). Brock Biology of Microorganisms (11th ed.). Englewood Cliffs, N.J: Prentice Hall. ISBN 0-13-144329-1.
- Garrett RA, Klenk H (2005). Archaea: Evolution, Physiology and Molecular Biology. WileyBlackwell. ISBN 1-4051-4404-1.
- Cavicchioli R (2007). Archaea: Molecular and Cellular Biology. American Society for Microbiology. ISBN 1-55581-391-7.
- Blum P, eld. (2008). Archaea: New Models for Prokaryotic Biology. Caister Academic Press. ISBN 978-1-904455-27-1.
- Lipps G (2008). "Archaeal Plasmids". Plasmids: Current Research and Future Trends. Caister Academic Press. ISBN 978-1-904455-35-6.
- Sapp J (2009). The New Foundations of Evolution: On the Tree of Life. New York: Oxford University Press. ISBN 0-19-538850-X.
- Schaechter M (2009). Archaea (Overview) in The Desk Encyclopedia of Microbiology (2nd ed.). San Diego and London: Elsevier Academic Press. ISBN 978-0-12-374980-2.
Vidu ankaŭ
[redakti | redakti fonton]Eksteraj ligiloj
[redakti | redakti fonton]- http://www.ncbi.nlm.nih.gov/genomes/lproks.cgi
- http://www.microbe.org/microbes/archaea.asp Arkivigite je 2006-07-20 per la retarkivo Wayback Machine Arĥeoj
- http://www.archaea.unsw.edu.au Arkivigite je 2007-02-11 per la retarkivo Wayback Machine Informo pri Arĥeoj
- http://www.mediscover.net/Extremophiles.cfm Arkivigite je 2006-04-23 per la retarkivo Wayback Machine
- En tiu ĉi artikolo estas uzita traduko de teksto el la artikolo Archaea en la angla Vikipedio.