Nostoc
| Nostoc | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 N. azollae | |||||||||||||
| Clasificación científica | |||||||||||||
| |||||||||||||
| Especies | |||||||||||||
|
N. azollae | |||||||||||||

Nostoc é un xénero de cianobacterias da familia Nostocaceae. A etimoloxía da palabra Nostoc é moi discutida[1], e crese que a puido inventar Paracelso (Nostoch). Designa hoxe un xénero de bacterias que realizan a fotosíntese e poden facer a fixación do nitróxeno.
Son moi parecidos a algunhas algas, mais poden vivir sobre substratos terrestres orixinando colonias, e formar masas xelatinosas semellantes a certas algas, especialmente cando as súas estruturas se inchan de auga despois da chuvia ou en período húmido.
Como pasan case desapercibidos cando están deshidratados e debido á rapidez con que se poñen turxentes cando se expoñen á humidade, existía a crenza popular de que caían do ceo, de aí o nome que teñen nalgúns países de "cuspiñazo de lúa" (crachat de lune en francés) ou "xelatina de estrelas" ou "manteiga de bruxa" (star jelly, witch's butter en inglés), entre outros.
Son especies pioneiras que poden vivir en medios pobres e básicos. Forman ás veces auténticos tapetes, nódulos[2] ou bólas, que acadan nas zonas polares varios centímetros de diámetro[3], e exercen localmente unha función moi importante protexendo os solos contra a erosión eólica e hídrica e a deshidratación en certos solos en pendente ou semidesérticos. Cando morren e se degradan contribúen a formar a materia orgánica do solo. A súa recolección intensiva na China é unha das causas da erosión e degradación de certos solos moi vulnerables.[4].
Identificación e taxonomía
[editar | editar a fonte]

A clasificación das especies destre xénero variou moito ao longo dos anos, e poderia aínda ser modificada conforme progrese o coñecemento da xenética do xénero.[5]
A súa identificación, igual que a das cianobacterias en xeral[6], necesita especialistas, que se basean para a determinación das especies na morfoloxía dos filamentos bacterianos, das células vexetativas, dos heterocistos e dos acinetos (todos observados con microscopio nunha mostra representativa). Características necesarias para a identificación de certos xéneros son a forma e o aspecto da colonia, a das células terminais dos filamentos ou a presenza de vesículas, mucilaxes, dunha vaíña máis ou menos grosa ou de gas, e as características do ciclo de vida.
Ao inicio da década do 2000, baseándose en resultados de estudos xenómicos, a separación dos xéneros Nostoc e Anabaena foi obxecto de discusións, como as feitas por Henson et al.[7] e Tamas et al.[8] por exemplo, Trichormus azollae podería ter que clasificarse como unha especie de Nostoc.[7].
Actualmente empeza a utilizarse con estas bacterias a PCR.[9]. Os datos son accesibles en GenBank.[10]
Segundo as primeiras análises filoxenéticas baseadas no estudo do ARN, a importante heteroxeneidade das cepas de Nostoc fai pensar que este grupo podería conter máis dun xénero.[11].
Estudáronse as proporcións e composición en ácidos graxos das cianobacterias e de diferentes cepas de Nostoc, como marcadores quimiotaxonómicos[12]. Un estudo feito en máis de 20 cepas indica que as cepas hepatotóxicas poderían ser reagrupadas por este método. Certas cepas de Nostoc producen cantidades importantes de ácidos graxos esenciais (omega 5, 6, 7, 9, 12)[12].
Descrición
[editar | editar a fonte]As especies do xénero Nostoc pode ter as formas máis variadas, e preséntanse a miúdo con aspectos moi diferentes vistos a escala macroscópica (colonias) o microscópica (filamentos).
- Certas especies, cando viven en colonias, teñen a aparencia de pequenas bólas (desde uns milímetros de diámetro ata un centímetro, e poden chagar a 10 cm excepcionalmente), polo que poden confundirse con fungos.
- Outras especies coloniais forman ensamblaxes lamelares, moi finos e case translúcidos, a miúdo circulares con beiras inchadas ou con ampolas e poden acadar de 4 a 6 cm de diámetro. A súa semellanza cunha alga ou ás veces cun lique pode inducir a confusións, mais trátase de procariotas.
- A forma que presentan outras especies lembra a cabelos finos longos verdes escuros. Os que viven en medios oligótrofos escasos en nitróxeno da meseta de Qinghai e do deserto do Gobi son utilizados na cociña chinesa, na que se chaman Fat Choy (Nostoc flagelliforme), e son cosumidos durante festas como as do ano novo lunar. O seu consumo está actualmente prohibido na China porque a especie está en perigo de desaparición e foi incluída na lista vermella, e ademais segrega toxinas (beta-N-metilamino-L-alanina) que poden afectar gravemente á saúde do consumidor. Outra especie, con forma de bóla, véndese deshidratado para a cociña na China[13].
Certas especies de Nostoc poden ás veces estar asociadas simbioticamentemente a plantas ou fungos.
Hábitat
[editar | editar a fonte]Poden atoparse en lugares onde outras especies non compiten con eles, é dicir, en medios moi pobres (ou novos) e expostos a unha forte insolación, por exemplo en certos camiños, na grava dos taludes, nas canteiras, sobre rochas, e mesmo sobre o asfalto, que son medios a priori pouco favorables ao seu desenvolvemento en canto á dispoñibilidade de materia mineral nitroxenada.
Certas especies poden atoparse preto do mar.
Características biolóxicas
[editar | editar a fonte]
O xénero Nostoc pertence ao grupo Hormogoneae.
Forman estruturas coloniais (aéreas ou acuáticas) moi resistentes aos raios UV, á radioactividade[14], aos pHs básicos, ó desecamento[15],[16] e a diversos estreses ambientais (choques térmicos fundamentalmente).
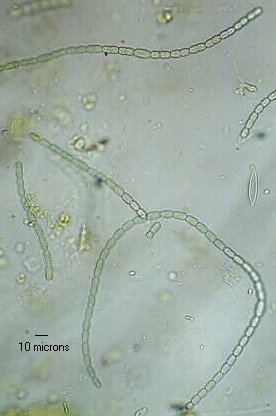
Teñen a capacidade de fixar o nitróxeno atmosférico por medio de células especiais: os heterocistos, que son células diferenciadas de paredes máis grosas, claramente visibles ao microscopio. Os heterocistos aparecen xeralmente nos extremos das cadeas de células esféricas, ovoides ou cilíndricas. Estas cadeas son máis ou menos longas. Nos grandes filamentos, os heterocistos poden tamén aparecer en posición intercalar. Son o resultado da diferenciación de células vexetativas. O proceso de produción de heterocistos comeza cando o organismo está estresado e carente de nitróxeno mineral. Efectúase só no 3 ao 7 % das células da colonia.
Hai especies do xénero Nostoc nunha gran variedade de medios, desde certos desertos, ao límite das zones polares, pasando por arrozais, onde fertilizan con nitróxeno os campos. Certas especies de Nostoc viven en epífitas, sobre a base dos talos das plantas de arroz, ou sobre certas especies de Characeae[17]. Nas súas células especializadas absorben o nitróxeno do aire sen que sexa transformado en amoníaco previamente, grazas a un encima que posúen chamado nitroxenase, que unicamente se expresa nos heterocistes.[18] Polo menos en certas especies ou cepas a luz é importante neste proceso[19], igual que a temperatura.[20] Comprobouse (2004) que as cepas de Nostoc que proceden de campos cultivados presentan neste sentido capacidades de fixación de nitróxeno moi diferentes en comparación coas que proceden de ambientes "silvestres" ou non cultivados.[19]
Reprodución
[editar | editar a fonte]Realízana de tres maneiras complementarias:
- Por hormogonios (estruturas móbiles, producidas por tricomas novos que presentan heterocistos nos dous extremos da cadea celular). Os acinetos, se están presentes, atópanse en posición intermedia (entre dous heterocistos e aparecen a miúdo en cadea).
- Por rotura do tricoma.
- Por xerminación dos acinetos.
Coñécese polo menos un caso (Nostoc punctiforme) de simbiose (concretamente endosimbiose) cun fungo, o da especie Geosiphon pyriformis (Kütz.) v. Wettstein (v. Wettstein, 1915)[21] e outras poden estar asociadas aos fungos de certos liques.[22]
Historia, evolución e paleoecoloxía
[editar | editar a fonte]Segundo os ficólogos, hai formas fósiles que semellan ser formas de Nostoc próximas ás que coñecemos actualmente, que datan polo menos do Precámbrico[23] Nostoc seguramente constituíu un elemento importante de numerosas comunidades terrestres e acuáticas desde esa época. As cianobacterias filamentosas do xénero Nostoc adquiriron, probablemente desde hai longo tempo, a capacidade de resistir en ambientes difíciles e de formar estruturas coloniais microscópicas e macroscópicas, mesmo xigantes a escala dunha bacteria. Son xeralmente discretas, pero comúns en todos os hábitats terrestres e acuáticoss, ata viven nos glaciares de montaña e nos ou sobre os xeos polares.[23]
Ecoloxía
[editar | editar a fonte]


En relación con outras cianobacterias[24], o xénero Nostoc presenta importantes especificidades ecolóxcas.
Unha gran parte do seu éxito nos ambientes emerxentes difíciles está ligada á súa capacidade de sobrevivir en forma deshidratada durante meses ou anos, e despois de rehidratarse reactivando rapidamente o seu metabolismo (en poucas horas despois dunha chuvia ou en contacto con auga líquida).
Nostoc resiste igualmente ciclos repetidos de conxelación e desconxelación e aos raios ultravioletas (maiormente en hábitats bénticos pouco profundos e moi expostos á luz solar). Grazas a isto, son un compoñente importante nos ambientes chamados extremos como os hábitats terrestres do Ártico e Antártico.[23]
Posúe ademais a propiedade de fixar o nitróxeno, o que lle dá vantaxe como especie pioneira en certos medios oligótrofos (que son pobres en nitróxeno[23]).
Este xénero parece desempeñar tamén un importante papel no cultivo do arroz, mellorando a súa produtividade biolóxica ao fixar nos campos de arroz o nitróxeno que pode despois liberarse e reutilizarse pola planta do arroz. Xoga tamén un papel na formación do neosol (solo en vías de formación) e no enriquecemento dos solos (achegan nitróxeno aos ecosistemas naturais e acuáticos).[23]
As súas aptitudes fan que poida competir cos liques, e Nostoc podería tamén xogar un papel aínda mal delimitado nas interaccións duradeiras e máis ou menos simbióticas con outras especies (fungos, liques, hepáticas, antocerotas, musgos, fentos, Cycas, e a anxiosperma Gunnera.[23]
Nostoc é moi resistente á predación ao formar colonias (ás veces de consistencia como de caucho, e resistentes a ser pisadas) bastante grandes para resistir aos alxívoros. As toxinas (de tipo microcistina) que produce tamén contribúen á súa resistencia á predación.[23]
Ten unha serie de inimigos naturais que regulan as súas poboacións, como son:
- diversos organismos que comen algas ou bacterias: algúns poden subsistir alimentándose só de Nostoc, aínda que este non sexa a súa fonte de alimentación preferida[23],
- cianófagos líticos que poden infectar a Nostoc[23]…
Pero aínda se ignora como se controlan ou retrocontrolan as dinámicas das poboacións de Nostoc no ambiente natural.[23]
Xenética
[editar | editar a fonte]Debido ás súas características, o xenoma deste xénero interesa aos xenetistas e á industria biotecnolóxica.[25] Entre estas características interesantes están que se trata dun organismo fotoautótrofo, que ten un modo de crecemento diazótrofo, pero facultativamente pode crecer como heterótrofo ; e que as súas células vexetativas teñen varias alternativas de desenvolvemento, incluída a diferenciación terminal ; e que pode fixar o nitróxeno e producir moléculas complexas que lle confiren oportunidades de sobrevivir en ambientes moi difíciles… A especie modelo estudada é Nostoc punctiforme, caracterizada por ter unha grande variabilidade fenotípica e grandes capacidades de establecer simbioses con fungos e plantas terrestres (briófitas, ximnospermas e anxiospermas). O seu xenoma publicouse en 2001.[25]
Comprobouse que esta especie de Nostoc é un organismo máis complexo do agardado, cun xenoma sorprendentemente importante para unha bacteria. Ten 8,9 Mb segundo a secuenciación dunha cepa de N. punctiforme (ATCC 29133) feita polo Joint Genome Institute ; o 45 % dos seus xenes codifican proteínas de función coñecida ou probable, e o 29 % codifican proteínas a priori exclusivas de N. punctiforme e de función descoñecida, pero posiblemente ligadas ás súas capacidades adaptativas ou simbióticas. O seu xenoma é complexo, con numerosas insercións, xenes codificantes de encimas transposases e modificacións do ADN.[25]
N. punctiforme posúe uns 400 xenes coñecidos que codifican proteínas quinases implicando sensores, reguladores da resposta e outros factores de trranscrición, o que demostra que é unha especie que pode percibir as modificacións ambientais e de realizar numerosas adaptacións.[25]
Substancias de interese bioquímico e farmacéutico
[editar | editar a fonte]Toxinas
[editar | editar a fonte]Numerosas cianobacterias e todas as cianobcterias filamentosas, producen toxinas chamadas ficotoxinas, que son neurotóxicas, hepatotóxicas ou presentan efectos biocidas sobre as algas, fungos, animais predadores ou outros.[26]. Producen ás veces varias toxinas diferentes; por exemplo, unha cepa de Nostoc sp. atopada nun lago de Finlandia demostrou in vitro poder producir unha batería de polo menos nove péptidos hepatotóxicos. Estas toxinas foron illadas, purificadas e estudadas.[26] Algunhas delas presentaban propiedades químicas e toxicolóxicas semellantes ás dos hepta- e pentapéptidos hepatotóxicos producidos por outras cianobacterias, pero algunhas (P14, P15 et P16) presentaban tipos novos de homólogos da microcistina-LR. A toxicidade destes homólogos non é significativamente diferente da dos homólogos xa coñecidos,[26] mais podería xogar un papel na resistencia da bacteria á evolución de capacidades de detoxificación nos seus predadors.
Resistencia aos raios ultravioleta
[editar | editar a fonte]Nun experimento realizado en 1990 os cultivos líquidos de Nostoc (mostras tomadas in situ) e expostas a unha irradiación con raios UV-B e UV-A sobreproduciron certos pigmentos protectores. Os UV-B fixeron incrementar a produción de carotenoides (equinenona e mixoxantofila fundamentalmente), sen inhibir a produción de clorofila a. Ao mesmo tempo, a bacteria producía en maior csntidade un pigmento extracelular, soluble en auga, unha micosporina que absorbe os raios UV-A/B, asociada á síntese de glicano extracelular. Sintetizaba tamén scytoneminas (pigmentos extracelulares liposolubles, coñecidos por funcionar como unha protección solar UV-A). Despois dunha longa exposición, o efecto dos UV-B sobre a síntese de carotenoides e da scytonemina cesa, aínda que o contido en micosporina permanece constantemente elevado. A produción de micosporina (que protexe contra os raios UV-B) é inducida exclusivamente pola exposición aos raios UV-B (<315 nm). A síntese de scytonemina (que protexe contra os raios UV-A solares) só é lixeiramente inducida polos UV-B (<315 nm), e moi fortemente polas radiacións próximas aos UV-A (de 350 a 400 nm), pero en absoluto polos UV-A (de 320 a 350 nm). Estes resultados fan pensar que as sínteses destes tres tipos de protectores solares anti-UV son activadas por tres fotorreceptores de UV diferentes, que aínda non se descubriron.[27].
Adaptación aos sales (osmorregulación)
[editar | editar a fonte]Certas especies presentan adaptacións osmorreguladoras que lles permiten adaptarse nalgunhas xeracións ao sal mariño. Por exemplo Nostoc muscorum, exposta ao sal, modifica a súa fisioloxía en varias etapas, e desenvolve múltiples mecanismos de resistancia celular, estables e duradeiros ante o sal (NaCl), e faino en só unhas poucas xeracións.[28]
Un mecanismo inicial combina unha estimulación da actividade fotosintética, que permite unha acumulación de sacarosa utilizada pola célula como osmorregulador. Un mecanismo secundario implica unha adaptación utilizando a fixación do nitróxeno (N2) e a biosíntese de novas proteínas útiles.[28]
Comprobouse no laboratorio que esta adaptación (a mái eficaz en resposta ao estrés inducido polo sal miriño, NaCl) só funciona parcialmente ante o estrés inducido por KCl ou por desequilibrios osmóticos de natureza non iónica por exemplo os debidos ao manitol.[28]
Antitumoral
[editar | editar a fonte]Certas toxinas celulares (criptoficinas) producidas de forma natural por certas cepas de Nostoc, e anteriormente identificadas na cepa Nostoc (ATCC 53789), parecen ser boas candidatas a ser compostos antitumorais e contra o cancro. Foron illadas en 1994 nunha cepa (GSV 224) de Nostoc.[29]. Teñen tamén actividade funxicida.
Confusións posibles
[editar | editar a fonte]Os Nostoc poden confundirse con algas ou liques. Tamén se poden confundir con certos fungos debido ao aspecto en tamaño, presenza de vesículas en certas especies de Nostoc que teñen tons pardos escuros ou anegrados.
Usos e supostos riscos sanitarios
[editar | editar a fonte]Diversas especies de Nostoc producen toxinas, a pesar de que son consumidas tradicionalmente en determinados lugares.
Así, no Perú as poboacións autóctonas apañaban nos lagos de montaña colonias de Nostoc (Nostoc commune globular, chamado localmente llullucha[30]). Estas colonias son comidas, intercambiadas por millo, ou vendidas nos mercados de Cuzco e outras cidades veciñas. No altiplano peruano é un alimento estacional importante, consumido só en «Picante» (un tipo de guiso tradicional)[30] que se di que é moi nutritivo.[30].
O Nostoc commune produce aminoácidos non habituais, dos cales algúns están en moléculas do grupo das micosporina, moléculas que poden contribuír á prevención dos danos pola exposición aos raios ultravioletas, aos cales Nostoc é especialmente resistente.
Nun estudo analizáronse 21 mostras de Nostoc commune esférico da rexión de Cuzco. Contiñan o aminoácido beta-N-metilamino-L-alanina (BMAA) producido por diversos taxons de cianobacterias chamadas neurotóxicas.[30]. A BMAA suponse que está implicada en varias doenzas neurodexenerativas, e os investigadores suxeriron que a enfermidade de Alzheimer ou a de Parkinson son máis frecuentes entre as persoas que consomen llullucha no Perú.[30]
Outro dos novos compostos illados dunha cepa de Nostoc (Nostoc 78-12A atopada nos Estados Unidos) foi o alcaloide nostocarbolina (β-carbolina, alcaloide cuaternario).[31] Esta molécula parece prometedora para aplicala ao tratamento da enfermidade de Alzheimer, ao inhibir o encima butirilcolinesterase (BChE)[32] tan eficazmente como a galantamina (medicamento utilizado actualmente). Foi patentada pola Escola Politécnica Federal de Zúric (EPFZ).[33] Ademais esta nostocarbolina é biodegradable e doada de producir, e parece ter virtudes biocidas (alxicidas[34]) polo que podería talvez algún día utilizarse en agricultura.[33]
Galería
[editar | editar a fonte]-
Nostoc en graveiras.
-
Nostoc en cultivo.
-
Colonia de Nostoc despois de dous días de chuvia.
-
Colonias de Nostoc aliñadas durante varios metros.




Lista de especies
[editar | editar a fonte]Segundo AlgaeBase [35]:
- Nostoc agglutinans Meneghini ex Kützing (Sen verificación)
- Nostoc album Gardner (Estatuto incerto)
- Nostoc alpinum Kützing (Sen verificación)
- Nostoc ambiceps C.C.Jao (Sen verificación)
- Nostoc amplissimum Setchell
- Nostoc anisococcum Sprengel (Sen verificación)
- Nostoc antarctica West & G.S.West (Sen verificación)
- Nostoc antillarum (P.L.Crouan & H.M.Crouan) P.L.Crouan & H.M.Crouan (Sen verificación)
- Nostoc apuanum De Notaris (Sen verificación)
- Nostoc apuanum Savi (Sen verificación)
- Nostoc arctum Kützing (Sen verificación)
- Nostoc arenarium Desmazières (Sen verificación)
- Nostoc atrovirens Chevallier (Sen verificación)
- Nostoc austinii H.C.Wood (Sen verificación)
- Nostoc beilschmiedianum Kützing (Sen verificación)
- Nostoc belmonticum C.G.M.Archibald (Sen verificación)
- Nostoc bicalyptratum Skuja (Sen verificación)
- Nostoc birnbaumii Corda (Sen verificación)
- Nostoc bohemicum Rabenhorst (Sen verificación)
- Nostoc borneti Gain
- Nostoc bornetii Gain (Sen verificación)
- Nostoc borzioides S.Skinner & T.J.Entwisle (Sen verificación)
- Nostoc botryoides (Linnaeus) C.Agardh (Sen verificación)
- Nostoc botryosum Rabenhorst (Sen verificación)
- Nostoc brittoni Gardner (Estatuto incerto)
- Nostoc brittonii Gardner (Sen verificación)
- Nostoc bullatum (A.P.de Candolle) Duby (Sen verificación)
- Nostoc caerulescens Rabenhorst (Estatuto incerto)
- Nostoc caeruleum Lyngbye ex Bornet & Flahault
- Nostoc caladarium H.C.Wood (Sen verificación)
- Nostoc caladarium Wood (Sen verificación)
- Nostoc calcicola Brébisson ex Bornet & Flahault
- Nostoc carneum C.Agardh ex Bornet & Flahault
- Nostoc catenatum P.A.Dangeard (Sen verificación)
- Nostoc cesatii Balsamo (Sen verificación)
- Nostoc cesatii Rabenhorst (Sen verificación)
- Nostoc ciniflonum Tournefort ex Bornet (Sen verificación)
- Nostoc ciniflorum C.Agardh (Sen verificación)
- Nostoc citrisporum Prasad & Mehrotra (Sen verificación)
- Nostoc coccymelon Kützing (Sen verificación)
- Nostoc coimbatorense Laloraya & Mitra (Sen verificación)
- Nostoc coimbatorense Laloraya (Sen verificación)
- Nostoc collinum Kützing (Sen verificación)
- Nostoc comminutum Kützing (Sen verificación)
- Nostoc commune Vaucher ex Bornet & Flahault (especie tipo)
- Nostoc confluens Kützing (Sen verificación)
- Nostoc confusum C.Agardh illeg. (Sen verificación)
- Nostoc conglomeratum Hirose (Sen verificación)
- Nostoc conicocellulare Huber-Pestalozzi (Sen verificación)
- Nostoc copiosum Kogan & Jazkulieva (Sen verificación)
- Nostoc cordubense C.H.Prosperi (Estatuto incerto)
- Nostoc coriaceum Vaucher (Sen verificación)
- Nostoc crassisporum Geitler (Sen verificación)
- Nostoc crassisporum Meneghini (Sen verificación)
- Nostoc crispum De Notaris (Sen verificación)
- Nostoc cristatum Bailey ex Harvey (Sen verificación)
- Nostoc cristatum J.Bailey (Sen verificación)
- Nostoc cuticulare (Brébisson) Bornet & Flahault (Sen verificación)
- Nostoc cycadae Maruyama & Fukushima (Sen verificación)
- Nostoc cylindrosporum De Notaris (Sen verificación)
- Nostoc delpinoi Borzí (Sen verificación)
- Nostoc depressum H.C.Wood (Sen verificación)
- Nostoc desertorum Reháková & Johansen
- Nostoc diamorphoticum Itzigsohn (Sen verificación)
- Nostoc dickiaei Wood (Sen verificación)
- Nostoc diplonema Montagne (Sen verificación)
- Nostoc disciforme F.E.Fritsch
- Nostoc edaphicum (Sen verificación)
- Nostoc edule Montagne & Berkeley (Sen verificación)
- Nostoc elgonense Naumann
- Nostoc ellipsoideum Gardner (Estatuto incerto)
- Nostoc ellipsosporum (Desmazieres) Rabenhorst (Sen verificación)
- Nostoc ellipsosporum Rabenhorst ex Bornet & Flahault
- Nostoc endophytum Bornet & Flahault
- Nostoc entophytum Bornet & Flahault
- Nostoc epilithicum Ercegovic
- Nostoc expansum Harvey & Bailey (Sen verificación)
- Nostoc falsum "Kützing" (Sen verificación)
- Nostoc filarzskyi De Toni (Sen verificación)
- Nostoc flavicans Bory nom.nud. (Sen verificación)
- Nostoc flosaquae (Linnaeus) Lyngbye (Sen verificación)
- Nostoc fluviatile Liljeblad (Sen verificación)
- Nostoc foliaceum Mougeot
- Nostoc fonticola Brabez (Sen verificación)
- Nostoc fragiforme (Roth) Brébisson (Sen verificación)
- Nostoc fritschii (Mitra) Schwabe & El Ayouty (Sen verificación)
- Nostoc furfuraceum Kützing (Sen verificación)
- Nostoc fuscescens F.E.Fritsch
- Nostoc fuscum Kützing (Sen verificación)
- Nostoc geitleri De Toni (Sen verificación)
- Nostoc gelatinosum Schousboe
- Nostoc ghotgewadense G.R.Hegde & K.Somanna (Estatuto incerto)
- Nostoc giganteum Mohr (Sen verificación)
- Nostoc glomeratum Kützing (Sen verificación)
- Nostoc granulare (Kützing) Rabenhorst (Sen verificación)
- Nostoc gregarium Thuret ex Kützing (Sen verificación)
- Nostoc gymnosphaericum Kützing (Sen verificación)
- Nostoc halophilum Hansgirg
- Nostoc hatei S.C.Dixit (Estatuto incerto)
- Nostoc heredulae (Meneghini) Bornet & Flahault (Sen verificación)
- Nostoc heterothrix Zeller (Sen verificación)
- Nostoc humifusum Carmichael ex Bornet & Flahault
- Nostoc hyalinum Bennett (Sen verificación)
- Nostoc hyalinum Roemer (Sen verificación)
- Nostoc hydrocoleoides Reinsch (Sen verificación)
- Nostoc imperfectum Schwabe & El Ayouty (Sen verificación)
- Nostoc indistinguendum Reháková & Johansen
- Nostoc insulare Borzi
- Nostoc intricatum Meneghini
- Nostoc inundatum Kützing (Sen verificación)
- Nostoc irregulare Wartmann (Sen verificación)
- Nostoc itzigsohnii Rabenhorst (Sen verificación)
- Nostoc kihlmanii Lemmermann (Sen verificación)
- Nostoc kihlmaniiLemmermann (Sen verificación)
- Nostoc killiasii Cramer (Sen verificación)
- Nostoc lacerum Kützing (Sen verificación)
- Nostoc lacunosum Montagne (Sen verificación)
- Nostoc lacustre Kützing (Sen verificación)
- Nostoc laevigatum Kützing (Sen verificación)
- Nostoc leptonema Reinsch (Sen verificación)
- Nostoc letestui Frémy
- Nostoc lichenoides Reháková & Johansen
- Nostoc lichenoides Vaucher ex Frank (Sen verificación)
- Nostoc limosum Zeller (Estatuto incerto)
- Nostoc linckia (Roth) Bornet ex Bornet & Flahault (Sen verificación)
- Nostoc litorale Kunze (Sen verificación)
- Nostoc littorale Kützing (Sen verificación)
- Nostoc lobatus Wood (Sen verificación)
- Nostoc longstaffi F.E.Fritsch
- Nostoc longstaffii F.E.Fritsch (Sen verificación)
- Nostoc macrosporum Meneghini ex Bornet & Flahault
- Nostoc maculiforme Bornet & Flahault
- Nostoc mamillosum C.C.Jao (Sen verificación)
- Nostoc margaritaceum (Kützing) Rabenhorst (Sen verificación)
- Nostoc membranaceum Gardner (Estatuto incerto)
- Nostoc microscopicum Carmichael ex Bornet & Flahault
- Nostoc microtis Montagne (Sen verificación)
- Nostoc minimum Currey (Sen verificación)
- Nostoc minutum Desmazières
- Nostoc molle C.Agardh (Sen verificación)
- Nostoc mougeotii Brébisson ex Kützing (Sen verificación)
- Nostoc muscorum C.Agardh ex Bornet & Flahault (Sen verificación)
- Nostoc myriococcum Montagne (Sen verificación)
- Nostoc natans Kützing
- Nostoc nivale Kützing (Sen verificación)
- Nostoc notarisii Franzoni (Sen verificación)
- Nostoc nylstromicum Claassen (Sen verificación)
- Nostoc opalinum Bennett (Sen verificación)
- Nostoc oryzae (F.E.Fritsch) J.Komárek & K.Anagnostidis
- Nostoc pachydermaticum Gain (Sen verificación)
- Nostoc palmelioides Kützing
- Nostoc paludosum Kützing ex Bornet & Flahault
- Nostoc papillosum Kurz (Sen verificación)
- Nostoc papyraceum S.F.Gray (Sen verificación)
- Nostoc paradoxum Welwitsch ex West & G.S.West (Sen verificación)
- Nostoc parietinum Rabenhorst (Sen verificación)
- Nostoc parmelioides Kützing ex Bornet & Flahault
- Nostoc parvulum C.C.Jao (Sen verificación)
- Nostoc passerinianum (De Notaris) Bornet (Sen verificación)
- Nostoc passerinianum (De Notaris) Bornet ex Bornet & Flahault (Sen verificación)
- Nostoc pellucidum Kützing (Sen verificación)
- Nostoc peloponnesiacum Kützing (Sen verificación)
- Nostoc peltigerae Letellier (Sen verificación)
- Nostoc piscinale Kützing ex Bornet & Flahault
- Nostoc planctonicum Poretsky & Tschernow
- Nostoc polysaccum Reinsch (Sen verificación)
- Nostoc polysporum Reinsch (Sen verificación)
- Nostoc populorum (Geit) Hollenb.
- Nostoc prismaticum Cesati (Sen verificación)
- Nostoc pruniforme C.Agardh ex Bornet & Flahault
- Nostoc pseudogelatinosum Claassen (Sen verificación)
- Nostoc punctatum H.C.Wood (Sen verificación)
- Nostoc punctiforme (Kützing) Hariot (Sen verificación)
- Nostoc purpurascens Kützing (Sen verificación)
- Nostoc purpureum G.Dickie (Sen verificación)
- Nostoc quoji C.Agardh (Sen verificación)
- Nostoc ramosum Ercegovi?
- Nostoc repandum West & G.S.West
- Nostoc reticulatum Roussel (Sen verificación)
- Nostoc rivulare Kützing ex Bornet & Flahault
- Nostoc rothii C.Agardh (Sen verificación)
- Nostoc rufescens C.Agardh (Sen verificación)
- Nostoc rugosum Kützing (Sen verificación)
- Nostoc rupestre Kützing (Sen verificación)
- Nostoc salsum Kützing (Sen verificación)
- Nostoc saxatile Zeller (Estatuto incerto)
- Nostoc sergianum Borzí (Sen verificación)
- Nostoc shensiense C.C.Jao (Sen verificación)
- Nostoc simulans Gardner (Estatuto incerto)
- Nostoc sphaericum Vaucher ex Bornet & Flahault
- Nostoc sphaeroides Kützing
- Nostoc sphaerosporum Gardner (Estatuto incerto)
- Nostoc spinosum Tiwari (Sen verificación)
- Nostoc spongiaeforme C.Agardh ex Bornet & Flahault
- Nostoc spongiiformeC .Agardh (Sen verificación)
- Nostoc strictum Martens (Sen verificación)
- Nostoc sudeticum Kützing (Sen verificación)
- Nostoc symbioticum F.V.Wettstein
- Nostoc tenax Thuret (Sen verificación)
- Nostoc tenuissimum (Kützing) Rabenhorst (Sen verificación)
- Nostoc tibeticum C.C.Jao & Y.Y.Lee (Sen verificación)
- Nostoc torulosum Hirose (Sen verificación)
- Nostoc trentepohlii Mohr (Sen verificación)
- Nostoc tuberculosum Kützing (Sen verificación)
- Nostoc undulatum Archibald (Sen verificación)
- Nostoc variegatum Moore (Sen verificación)
- Nostoc verrucosum Vaucher ex Bornet & Flahault
- Nostoc vesicarium (Bull.) De Candolle (Sen verificación)
- Nostoc vesicarium (Bull.) De Candolle ex Frank (Sen verificación)
- Nostoc vulgare Schrank (Sen verificación)
- Nostoc wichmannii Weber-van Bosse
- Nostoc willei Gardner (Estatuto incerto)
- Nostoc wollnyanum P.Richter (Sen verificación)
- Nostoc zetterstedtii Areschoug
Segundo o NCBI[36] :
Segundo WoRMS [37]:
- Nostoc album
- Nostoc ramosum Ercegovic, 1925
Segundo o ITIS [38]:
- Nostoc caeruleum Lyngby
- Nostoc calcicola Brebisson
- Nostoc coeruleum Lyngbya
- Nostoc comminutum Kuetzing
- Nostoc commune Vaucher Ex Bornet & Flahault
- Nostoc depressum Wood
- Nostoc disciforme Fritsch
- Nostoc ellipsosporum (Des.) Rabenh.
- Nostoc kihlmani Lemmermann
- Nostoc macrosporum Menegh.
- Nostoc microscopicum Carm.
- Nostoc minutum Desmaz
- Nostoc muscorum C. Agardh
- Nostoc parmelioides Kuetzing
- Nostoc peltigerae Letellier
- Nostoc pruniforme C. Agardh
- Nostoc sphaericum Vaucher
- Nostoc spumigena (Mert.)
- Nostoc verrucosum Vauch.
Notas
[editar | editar a fonte]- ↑ Potts, M. (1997). Etymology of the genus name Nostoc (Cyanobacteria). Int J Syst Bacteriol 47, 584. (Ficha INIST/CNRS Arquivado 10 de xuño de 2015 en Wayback Machine.)
- ↑ University of California, Berkeley, Photo de Nostoc sp., por John Dittes
- ↑ leculler, [1] Dartmouth IGERT – Polar Environmental Change «The Arctic finally warms up : Getting ready for an expedition!» Local Knowledge 15 de xuño de 2011
- ↑ ver por exemplo Ver parágrafo de « Fat Choy » (Nostoc flagelliforme) Arquivado 10 de outubro de 2004 en Wayback Machine.
- ↑ Komarek, J., & Anagnostidis, K. (1989). Modern approach to the classification system of cyanophytes. ; Nostocales. Arch Hydrobiol Suppl 823 (Algol Stud 56), 247-345.
- ↑ Rippka, R. (1988). Recognition and identification of cyanobacteria. Methods Enzymol 167, 28-67.
- ↑ 7,0 7,1 Henson, B. J., Watson, L. E., & Barnum, S. R. (2002). Molecular differentiation of the heterocystous cyanobacteria, Nostoc and Anabaena, based on complete nifD sequences. Curr Microbiol 45, 161–164.
- ↑ Tamas, I., Svircev, Z., & Andersson, S. G. (2000). Determinative value of a portion of the nifH sequence for the genera Nostoc and Anabaena (cyanobacteria). Curr Microbiol 41, 197–200.
- ↑ Rasmussen, U., & Svenning, M. M. (1998). Fingerprinting of cyanobacteria based on PCR with primers derived from short and long tandemly repeated repetitive sequences. Appl Environ Microbiol 64, 265-272.
- ↑ The GenBank/EMBL/DDBJ accession numbers are AJ630408-AJ630458 for the 16S rRNA gene sequences, AJ632022–AJ632070 for the rbcLX gene sequences and AJ628068–AJ628134 for rpoB gene sequences determined in this study.
- ↑ Pirjo Rajaniemi, Pavel Hrouzek, Klára Kaštovská, Raphaël Willame, Anne Rantala, Lucien Hoffmann, Jiří Komárek et Kaarina Sivonen, [kaarina.sivonen{at}helsinki.fi Phylogenetic and morphological evaluation of the genera Anabaena, Aphanizomenon, Trichormus and Nostoc (Nostocales, Cyanobacteria)] ; doi : 10.1099/ijs.0.63276-0 IJSEM January 2005 vol. 55 n° 1 11-26 (resumo Arquivado 28 de febreiro de 2015 en Wayback Machine.)
- ↑ 12,0 12,1 Gugger, M., Lyra, C., Suominen, I., Tsitko, I., Humbert, J. F., Salkinoja-Salonen, M. S., & Sivonen, K. (2002). Cellular fatty acids as chemotaxonomic markers of the genera Anabaena, Aphanizomenon, Microcystis, Nostoc and Planktothrix (cyanobacteria). Int J Syst Evol Microbiol 52, 1007–1015.
- ↑ S. Facciola, Cornucopia II: A Sourcebook of Edible Plants, 1998
- ↑ Kraus, M. P. (1969). Resistance of blue-green algae to Co gamma radiation. Radiat Bot 9, 481-489.
- ↑ Shirkey, B., Kovarcik, D. P., Wright, D. J., Wilmoth, G., Prickett, T. F., Helm, R. F., Gregory, E. M. & Potts, M. (2000). Active Fecontaining superoxide dismutase and abundant sodF mRNA in Nostoc commune (Cyanobacteria) after years of desiccation. J Bacteriol 182, 189±197.
- ↑ Xie, W.-Q., Tice, D. & Potts, M. (1995). Cell water de®cit regulates expression of rpoC1C2 (RNA polymerase) at the level of mRNA in desiccation-tolerant Nostoc commune UTEX 584 (Cyanobacteria). FEMS Microbiol Lett 126, 159±164.
- ↑ Jean-Louis Garcia, Taxonomie des Procaryotes ; Cyanobacteria: Phylum BXI du Domaine Bacteria Arquivado 10 de decembro de 2015 en Wayback Machine., consultado o 22 de xaneiro de 2012
- ↑ Hrouzek, P., Šimek, M., & Komárek, J. (2003). Nitrogenase activity (acetylene reduction activity) and diversity of six soil Nostoc strains. Arch Hydrobiol Suppl 146, 87–101.
- ↑ 19,0 19,1 P. Hrouzek, A. Lukešová et M. Šimek, Comparison of light and dark nitrogenase activity in selected soil cyanobacteria ; Folia Microbiologica Volume 49, Number 4, 435-440, 2004 ; DOI : 10.1007/BF02931606 (resumo Arquivado 13 de setembro de 2019 en Wayback Machine.)
- ↑ A. K. Kashyap, K. D. Pandey et R. K. Gupta (1991), Nitrogenase activity of the antarctic cyanobacterium Nostoc commune : Influence of temperature ; Folia Microbiologica Volume 36, Number 6, 557-560 ; DOI : 10.1007/BF02884036 ([resumo])
- ↑ Geosiphon - endocyanosis ; The Geosiphon pyriformis symbiosis - fungus 'eats' cyanobacterium Arquivado 30 de xuño de 2019 en Wayback Machine.{
- ↑ Paulsrud, P., Rikkinen, J., & Lindblad, P. (2000). Spatial patterns of photobiont diversity in some Nostoc-containing lichens. New Phytol 146, 291±299.
- ↑ 23,00 23,01 23,02 23,03 23,04 23,05 23,06 23,07 23,08 23,09 Dodds, W. K., Gudder, D. A. and Mollenhauer, D. (1995), The Ecology of Nostoc. Journal of Phycology, 31 : 2–18. DOI : 10.1111/j.0022-3646.1995.00002.x ; xuño de 2008 (resumo).
- ↑ Potts, M. (2000). Nostoc. In The Ecology of Cyanobacteria, páxinas 465-504. Edited by B. A. Whitton & M. Potts. Dordrecht : Kluwer Academic.
- ↑ 25,0 25,1 25,2 25,3 John C. Meeks, Jeff Elhai, Teresa Thiel, Malcolm Potts, Frank Larimer, Jane Lamerdin, Paul Predki & Ronald Atlas, An overview of the genome of Nostoc punctiforme, a multicellular, symbiotic cyanobacterium ; Photosynthesis Research 70 : 85–106, 2001. ; Kluwer Academic Publishers (Netherlands). Minireview ; Section of Microbiology, University, PDF, 22 pages, (résumé Arquivado 27 de setembro de 2013 en Wayback Machine.).
- ↑ 26,0 26,1 26,2 K. Sivonen, W. W. Carmichael, M. Namikoshi, K. L. Rinehart, A. M. Dahlem et S. I. Niemelä, Isolation and characterization of hepatotoxic microcystin homologs from the filamentous freshwater cyanobacterium Nostoc sp. strain 152 ; Appl. Environ. Microbiol. septembre 1990 vol. 56 n° 9 2650-2657 (résumé Arquivado 29 de novembro de 2017 en Wayback Machine.)
- ↑ M. Ehling-Schulz, W. Bilger et S. Scherer, UV-B-induced synthesis of photoprotective pigments and extracellular polysaccharides in the terrestrial cyanobacterium Nostoc commune ; J. Bacteriol. marzo de 1997 vol. 179 n° 6 1940-1945 (resumo Arquivado 13 de setembro de 2019 en Wayback Machine.)
- ↑ 28,0 28,1 28,2 Eduardo Blumwald et Elisha Tel-Or, Osmoregulation and cell composition in salt-adaptation of Nostoc muscorum ; Archives of Microbiology Volume 132, 1981, Number 2, 168-172 ; DOI : 10.1007/BF00508725 (résumé Arquivado 13 de setembro de 2019 en Wayback Machine.)
- ↑ Golakoti Trimurtulu, Ikuko Ohtani, Gregory M. L. Patterson, Richard E. Moore, Thomas H. Corbett, Frederick A. Valeriote, Lisa Demchik, Total Structures of Cryptophycins, Potent Antitumor Depsipeptides from the Blue-Green Alga Nostoc sp. Strain GSV 224 ; J. Am. Chem. Soc., 1994, 116 (11), páxinas 4729–4737 xuño de 1994, ; DOI : 10.1021/ja00090a020 (resumo / Primeira páxina do artigo)
- ↑ 30,0 30,1 30,2 30,3 30,4 Johnson HE, King SR, Banack SA, Webster C., Callanaupa WJ, Cox PA., Cyanobacteria (Nostoc commune) used as a dietary item in the Peruvian highlands produce the neurotoxic amino acid BMAA, J Ethnopharmacol. 19 de xuño de 2008 ; 118(1) : 159-65. Epub 2008 Apr 14.
- ↑ Paul G. Becher, Julien Beuchat, Karl Gademann, et Friedrich Jüttner ; Nostocarboline : Isolation and Synthesis of a New Cholinesterase Inhibitor from Nostoc 78-12A ; J. Nat. Prod., 2005, 68 (12), páxinas 1793–1795. DOI : 10.1021/np050312l. Data de publicación (Web) : 2005-12-02 (resumo).
- ↑ cun IC50 de 13,2 μM (fonte)
- ↑ 33,0 33,1 SuisseInfo La lutte contre l'Alzheimer part des eaux usées Arquivado 13 de setembro de 2019 en Wayback Machine. (2006-01-06), citando a revista universitaria « ETHLife » e « unipublic ». Consultado o 20-01-2012.
- ↑ Judith F. Blom, Tobias Brütsch, Damien Barbaras, Yann Bethuel, Hans H. Locher, Christian Hubschwerlen et Karl Gademann, Potent Algicides Based on the Cyanobacterial Alkaloid Nostocarboline ; Organic Letters 2006 8 (4), páxinas 737–740 (résumé)
- ↑ Guiry, M.D.; Guiry, G.M. (2008). "42976". AlgaeBase. World-wide electronic publication, National University of Ireland, Galway.
- ↑ NCBI Nostoc
- ↑ WoRMS. "Nostoc, Vaucher ex Bornet et Flahault, 1886". World Register of Marine Species.
- ↑ ITIS Nostoc
Referencias taxonómicas
[editar | editar a fonte]- Guiry, M.D.; Guiry, G.M. (2008). "42976". AlgaeBase. World-wide electronic publication, National University of Ireland, Galway.
- Sistema Integrado de Información Taxonómica. "Nostoc (TSN 1146)" (en inglés).
- WoRMS. "Nostoc, Vaucher ex Bornet et Flahault, 1886". World Register of Marine Species.
- Páxina da taxonomía do NCBI para Nostoc
- Procurar páxinas da taxonomía de Tree of Life para Nostoc
- Procurar páxinas en Species2000 para Nostoc
- Páxina de MicrobeWiki para Nostoc
Véxase tamén
[editar | editar a fonte]| Commons ten máis contidos multimedia sobre: Nostoc |
| Wikispecies posúe unha páxina sobre: Nostoc |
Bibliografía
[editar | editar a fonte]- (en francés) "Pluie de viande du Kentucky". In L'Exposition universelle de 1878 illustrée [París], ISSN 2118-8262, n° 67, août 1876, pp. 54–55.
- (en inglés) Adams DG et Duggan PS (1999) Heterocyst differentiation and akinete differentiation in cyanobacteria. New Phytol 144 : 3–33.
- (en inglés) Bhaya D, Schwarz R e Grossman AR (2000) Molecular responses to environmental stress. In : Whitton BA et Potts M. (eds) The Ecology of Cyanobacteria. Their Diversity in Time and Space, páxinas 397–442. Kluwer Academic Publishers, Dordrecht, Pays-Bas.
- (en inglés) Buikema WJ et Haselkorn R. (1993) Molecular genetics of cyanobacterial development. Ann Rev Plant Physiol Plant Mol Biol 44 : 33–52.
- (en inglés) Campbell EL e Meeks JC (1992) Evidence for plant-mediated regulation of nitrogenase expression in the Anthoceros-Nostoc symbiotic association. J Gen Microbiol 138 : 473–480.
- (en inglés) Campbell EL, Brahamsha B. et Meeks JC (1998) Mutation of an alternative sigma factor in the cyanobacterium Nostoc punctiforme results in increased infection of its symbiotic plant partner, Anthoceros punctatus. J Bacteriol 180 : 4938–4941.
- (en inglés) Cohen MF et Meeks JC (1997) A hormogonium regulating locus hrmUA, of the cyanobacterium Nostoc punctiforme strain ATCC 29133 and its response to an extract of a symbiotic plant partner Anthoceros punctatus. Mol Plant Microbe Inter 10 : 280–289.
- (en inglés) Dow CS et Swoboda UK (2000) Cyanotoxins. In : Whitton BA et Potts M. (eds) The Ecology of Cyanobacteria Their Diversity in Time and Space, páxinas 613–632. Kluwer Academic Publishers, Dordrecht, Pays-Bas.
- (en inglés) Enderlin CS et Meeks JC (1983) Pure culture and reconstitution of the Anthoceros-Nostoc symbiotic association. Planta 158 : 157–165.
- (en inglés) Helm, R. F., Huang, Z., Edwards, D., Leeson, H., Peery, W. 0., & Potts, M. (2000). Structural characterization of the released polysaccharide of desiccation-tolerant Nostoc commune DRH-1. J Bacteriol 182, 974±982.
- (en inglés) Hill, D. R. H., Hladun, S. L., Scherer, S. 0., & Potts, M. (1994b). Water stress proteins of Nostoc commune (Cyanobacteria) are secreted with UV-A}B-absorbing pigments and associate with 1,4-b-dxylanxylanohydrolase activity. J Biol Chem 269, 7726±7734.
- (en inglés) Mollenhauer D., Mollenahuer R. et Kluge M. (1996) Studies on initiation and development of the partner association in Geosiphon pyriforme (Kutz.) v.Wettstein, a unique encocytobiotic system of a fungus (Glomales) and the cyanobacterium Nostoc punctiforme (Kutz.). Hariot Protoplasma 193 : 3–9.
- (en inglés) Summers ML, Wallis JG, Campbell EL et Meeks JC (1995) Genetic evidence of a major role for glucose-6-phosphate dehydrogenase in nitrogen fixation and dark growth of the cyanobacterium Nostoc sp. strain ATCC 29133. J Bacteriol 177 : 6184–6194
- (en inglés) Wong FCY et Meeks JC (2001) The hetF gene product is essential to heterocyst differentiation and affects HetR function in the cyanobacterium Nostoc punctiforme. J Bacteriol 183 : 2654–2661.
- (en inglés)Wright, D.; Prickett, T.; Helm, R. F.; Potts, M. (2001-09-01). "Form species Nostoc commune (Cyanobacteria)". International Journal of Systematica and Evolutionary Microbiology (en inglés) 51 (5): 1839–1852. ISSN 1466-5026. doi:10.1099/00207713-51-5-1839.