Glikolisis

Glikolisis ("penguraian glukosa") ialah sebuah laluan metabolisme, yakni rantaian tindak balas biokimia yang berlaku dalam sitosol sel untuk memproses molekul gula lazim, glukosa (C
6H
12O
6) menjadi piruvat CH
3COCO
2H). Semua tisu menggunakan laluan glikolisis untuk memecahkan glukosa, menjanakan tenaga dan bahan perantara untuk tindak balas biosintesis. Glikolisis ialah pusat untuk metabolisme karbohidrat.
Glikolisis dibahagikan kepada dua peringkat iaitu:
1- Penukaran heksosa kepada dua triosa
2- Penukaran triosa fosfat kepada piruvat
Hasil
[sunting | sunting sumber]Glikolisis menghasilkan empat ATP (Adenosina trifosfat). Namun, 2 ATP terpaksa digunakan dalam memulakan langkah-langkah awal glikolisis. Maka, hasil bersih ATP yang dihasilkan dalam glikolisis adalah dua sahaja. Selain ATP, glikolisis juga menghasilkan dua NADH. NADH akan memasuki pemfosforilan oksidatif manakala piruvat memasuki kitar Krebs.
Proses tindak balas Glikolisis
[sunting | sunting sumber]Rajah Laluan Glikolisis
[sunting | sunting sumber]ATP
ADP
ATP
ADP
+ +
2 ×
 2 ×
 2 × 3-Fosfogliserat 2 ×
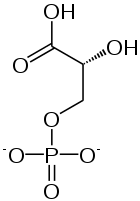 2 × 2-Fosfogliserat 2 ×
 2 × Fosfoenolpiruvat 2 ×
 ADP
ATP
2 × Piruvat 2 ×
|





Langkah-langkah dalam laluan glikolisis:
1- Pemfosforilan glukosa kepada glukosa 6-fosfat
2- Pengisomeran glukosa 6-fosfat kepada fruktosa 6-fosfat
3- Pemfosforilan fruktosa 6-fosfat kepada fruktosa 1,6-bisfosfat
4- Pembelahan fruktosa 1,6-bisfosfat kepada dua molekul tiga-karbon
5- Pengisomeran dihidroksiaseton fosfat kepada gliseraldehid 3-fosfat
6- Pembentukan 1,3-bisfosfogliserat daripada gliseraldehid 3-fosfat
7- Penukaran 1,3-bisfosfogliserat kepada 3-fosfogliserat
8- Penukaran 3-fosfogliserat kepada 2-fosfogliserat
9- Pendehidratan 2-fosfogliserat kepada fosfoenolpiruvat
10- Pembentukan piruvat daripada fosfoenolpiruvat
Fasa Penyediaan
[sunting | sunting sumber]Langkah pertama glikolisis adalah fosforilasi D-glukosa (Glc) kepada Glukosa-6-fosfat (G6P). Bergantung kepada jenis sel, tindak balas ini menjadi pemangkin oleh hexokinase atau glucokinase (hexokinase IV), di mana molekul ATP diperlukan. Ini mempunyai dua kelebihan: Di satu pihak, membran sel telap kepada glukosa kerana saluran glukosa yang terdapat di dalamnya (contohnya GLUT-1), tetapi tidak kepada glukosa-6-fosfat yang dihasilkan oleh fosforilasi. Akibatnya, ia berkumpul di dalam sel. Sebaliknya, fosforilasi glukosa mengurangkan kepekatan glukosa dalam sel, manakala kepekatan G6P meningkat sebaliknya. Fosforilasi awal menyebabkan kurang glukosa di dalam sel daripada di luar sel. Oleh kerana kepekatan glukosa intrasel tidak seimbang dengan ekstraselular, glukosa selanjutnya mengalir bersama-sama ini menyebabkan kecerunan kepekatan melalui saluran glukosa sel. Akibatnya, glukosa dioksidakan.
Dalam bakteria, fosforilasi dalam langkah pertama glikolisis tidak pemangkin oleh hexo- atau glucokinases, tetapi oleh sistem phosphoenolpyruvate (PEP) yang bergantung kepada gula-phosphotransferase.
Langkah Kedua, Glukosa-6-fosfat kemudiannya ditukar daripada isomerase glukosa-6-fosfat ke dalam fruktosa-6-fosfat isomerik (F6P). Enzim lebih suka mengikat alpha-anomer G6P, sebagai produk tindak balas α-D-fruktosa-6-fosfat terbentuk. Di bawah keadaan standard, keseimbangan tindak balas isomerisasi berada di sisi G6P. Tetapi sejak F6P terus bertindak balas dengan cepat, ia dikeluarkan dari sistem tindak balas, supaya tiada keseimbangan ditubuhkan dan tindak balas isomerisasi berjalan memihak kepada F6P.
Fruktosa-6-fosfat kemudiannya fosforlated dengan molekul ATP kepada fruktosa-1,6-bisphosphate (F1,6bP) di bawah tindakan enzim utama glikolis, phosphofructokinase 1, menghasilkan ADP. Enzim lebih suka beta-anomer F6P, manakala alpha-anomer terbentuk dalam pra-tindak balas. Walau bagaimanapun, ini tidak menjadi masalah kerana kedua-dua anomer berada dalam keseimbangan (mutarotation). Dalam bakteria anaerobik, sesetengah tumbuh-tumbuhan, eukariot primitif dan beberapa archaea, langkah ini menjadi pemangkin oleh fosfofruktokinase yang bergantung kepada pirofosfat (EC 2.7.1.90), di mana pirofosfat (PPi, bukan organik) digunakan dan bukannya ATP.
Pelaburan tenaga yang diperbaharui adalah murah dan perlu kerana dua sebab: Di satu pihak, langkah ini - sebagai tambahan kepada glucokinase dan pyruvate kinase - menjadikan glikolisis tidak dapat dipulihkan di bawah keadaan fisiologi. Sebaliknya, tenaga yang dibekalkan di sini membolehkan belahan hexose dalam langkah seterusnya dan dengan itu pembentukan dua trio fosforlated untuk kemerosotan selanjutnya, fosfat dihydroxyacetone (DHAP) dan gliseraldehid-3-fosfat (GAP). Atom karbon C1-C3 F1.6bP ditemui dalam DHAP, manakala atom C dalam GAP berasal dari unit C4-C6 F1.6bP.
Tindak balas pembelahan (lisis) sangat tidak menguntungkan di bawah keadaan standard (ΔG0' = +24 kJ / mol) dan tidak akan berlaku. Oleh kerana metabolisme pesat kedua-dua produk tindak balas, bagaimanapun, ia berlaku di bawah keadaan fisiologi dalam keseimbangan anggaran. dihidroksiaseton fosfat akan ditukar oleh enzim isomerase fosfat triose (TIM) kepada D-gliseraldehid 3-fosfat. Isomerisasi stereospesifik ini ke arah GAP disukai oleh fakta bahawa GAP terus merosot dalam glikolisis dan dengan itu kepekatan dalam sel disimpan rendah. Tanpa metabolisme lanjut, keseimbangan antara DHAP dan GAP akan kuat pada bahagian keton, iaitu DHAP (22:1).
Fasa Bayaran Balik
[sunting | sunting sumber]Setiap satu daripada dua molekul gliseraldehid-3-fosfat yang terhasil teroksida oleh glyceraldehyde-3-fosfat dehidrogenase (GAPDH) pada permulaan fasa bayaran balik glikolisis. Dalam tindak balas, NAD + dikurangkan kepada NADH. Pengoksidaan kumpulan aldehid (GAP) kepada kumpulan karboksi sangat bertenaga. Ia dieksploitasi untuk menghubungkan fosfat bukan organik kepada kumpulan karboksi. Ini menghasilkan asid campuran anhidrida 1,3-bisphosphoglycerate (1,3-BPG). Keseimbangan tindak balas ini adalah pada bahagian GAP bahan permulaan berbanding 1,3-BPG (10:1). Oleh kerana pelaksanaan pesat produk, bagaimanapun, pelarasan keseimbangan dicegah dan 1,3-BPG sentiasa terbentuk, di samping itu, kepekatan NAD + yang tinggi berbanding NADH menyokong penukaran dalam satu arah.
Laluan sekunder alternatif yang terdapat dalam erythrocytes, dari 1,3-bisphosphoglycerate hingga 3-phosphoglycerate, adalah kitaran Rapoport-Luebering melalui perantaraan 2,3-bisphosphoglycerate, yang enzim pusatnya adalah bisphosphoglycerate mutase trifunctional.
Dalam langkah seterusnya, kinase phosphoglycerate menghasilkan satu molekul ATP setiap satu semasa penukaran 1,3-bisphosphoglycerate kepada 3-phosphoglycerate dengan memindahkan residu fosfat kepada ADP. Tenaga yang dilepaskan semasa pengoksidaan sebelumnya dipulihara dengan membina ATP. Pembentukan ATP dari ADP yang berlaku di sini adalah contoh fosforilasi rantaian substrat. Jika sel sudah mempunyai banyak ATP (dan dengan itu sedikit ADP), tindak balas berterusan pada ketika ini sehingga ADP yang mencukupi tersedia semula. Peraturan maklum balas ini penting kerana ATP mereput dengan cepat apabila tidak digunakan. Oleh itu, pengeluaran ATP yang berlebihan dihalang.
Keseimbangan tenaga glikolisis seimbang pada langkah ini: dua molekul ATP telah digunakan dan dua pulih
Mutase fosfogliserat bebas kofaktor (PGM) kemudian memangkin penukaran 3-phosphoglycerate kepada 2-phosphoglycerate. Semasa proses itu, kumpulan fosfat dipindahkan sementara ke sisa asid amino enzim. Dalam erythrocytes, tindak balas ini menjadi pemangkin oleh PGM yang bergantung kepada kofaktor di mana 2,3-bisphosphogylcerate dibentuk sebagai perantaraan.
2-phosphoglycerate kemudiannya dehidrasi kepada phosphoenolpyruvate (PEP) dengan bantuan enolase. Oleh itu, enzim ini juga dipanggil 2-phosphoglycerate dehydratase. PEP adalah sebatian fosforilasi dengan potensi pemindahan kumpulan yang sangat tinggi. Ini digunakan dalam langkah terakhir glikolisis untuk mendapatkan molekul ATP yang lain. Di sini, kinase pyruvate (PK) memangkin penukaran PEP kepada pyruvate (= anion asid piruvat) di bawah keuntungan ATP. Walau bagaimanapun, ini tidak menghasilkan pyruvate secara langsung, tetapi keseimbangan enolpyruvate. Pada pH 7, keseimbangan berada di sisi bentuk keton. Langkah ini juga dikawal selia ADP, ia adalah tindak balas ketiga yang tidak dapat dipulihkan dalam perjalanan.
Dalam kes kekurangan fosfat, tumbuh-tumbuhan boleh menghidrolisis PEP untuk pyruvate tanpa keuntungan ATP, yang berlaku di vakuol. Enzim yang terlibat adalah fosfatase PEP (EC 3.1.3.60), yang melepaskan fosfat bukan organik dan dengan itu mengatasi kekurangan fosfat.
Pautan luar
[sunting | sunting sumber]- Enzim glikolisis di RCSB PDB
- Kitaran glikolisis beranimasi di wdv.com
- Poster laluan biokimia Expasy di ExPASy
- MedicalMnemonics.com: 317 5468
{{#switch:||Kategori=