Neurona
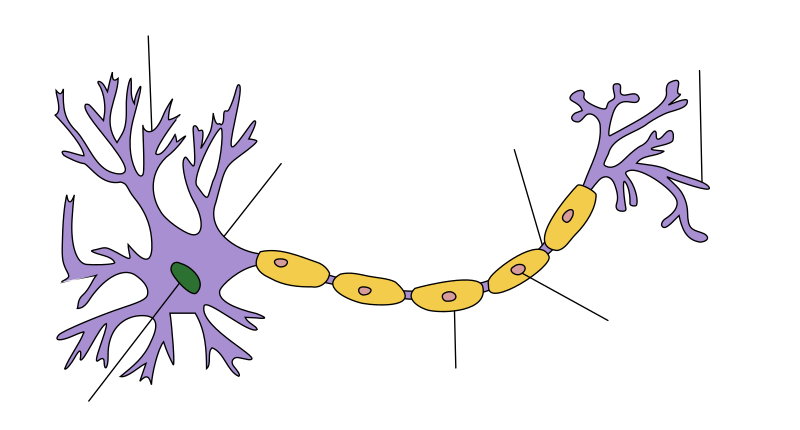
Unha neurona (do grego νεῦρον, corda, nervio[1]) ou célula nerviosa é un tipo de célula pertencente ao sistema nervioso cuxa membrana plasmática ten a característica de excitarse con electricidade para procesar e transmitir información por medio de impulsos eléctricos e químicos. Son un tipo de células altamente diferenciadas e non teñen a habilidade de se dividiren, aínda que existan algunhas que o fagan[2]. As neuronas conéctanse unhas coas outras formando enormes redes neuronais. A súa morfoloxía está adaptada ás funcións que realizan: un corpo celular chamado soma; unha ou varias prolongacións curtas que xeralmente transmiten impulsos cara ao soma, denominadas dendritas; e unha prolongación longa que conduce os impulsos desde o soma ata a seguinte neurona, chamada axón[3].
| Estrutura dunha neurona típica |
|---|

Xeneralidades
A neurona é un tipo especial de célula atopada no corpo dos animais coa excepción das esponxas. As características que definen unha neurona son a excitabilidade eléctrica e a presenza de sinapses, que son complexas unións de membrana que transmiten sinais dunha célula á seguinte. As neuronas do corpo, xunto coas células da glía que lles dan soporte estrutural e metabólico, constitúen o sistema nervioso. Nos vertebrados a maioría das neuronas forman parte do sistema nervioso central, pero hai outras que están situadas nos ganglios periféricos, e moitas neuronas sensoriais están situadas nos órganos dos sentidos, como a retina ou a cóclea do oído, por exemplo.
As neuronas son moi diversas e sempre hai excepcións a case calquera xeneralización, pero é conveniente empezar facendo unha descrición xeral da estrutura e función dunha neurona típica. Unha neurona típica está dividida en tres partes: soma ou corpo celular, dendritas, e axón. O soma é normalmente compacto; o axón e as dendritas son filamentos que se estenden desde o soma. As dendritas ramifícanse profusamente, facéndose máis delgadas en cada ramificación, e chegan coas últimas ramificacións a varios centros de microns do soma. O axón parte do soma desde un avultamento chamado protuberancia do axón ou cono axónico, e pode estenderse a grande distancia, dando lugar ao final a centos de ramificacións, aínda que xeralmente mantén o seu diámetro durante toda a súa extensión. O soma pode orixinar numerosas dendritas, pero nunca máis dun axón. Os sinais sinápticos que chegan das outras neuronas recíbense no soma e dendritas; e os sinais que se envían a outras neuronas saen polo axón. Unha sinapse típica é o contacto entre o axón dunha neurona e unha dendrita ou o soma doutra. Os sinais sinápticos poden ser excitatorios ou inhibitorios. Se a excitación neta recibida por unha neurona é o suficientemente intensa durante un curto período de tempo, a neurona xera un breve pulso eléctrico chamado potencial de acción, o cal se orixina no soma e se propaga rapidamente ao longo do axón, activando sinapses con outras neuronas a medida que se despraza.
Moitas neuronas axústanse perfectamente a este esquema xeral, pero hai excepcións. Non hai neuronas que carezan de soma, pero hai neuronas sen dendritas, ou sen axón. Ademais, xunto coas sinapses típicas axodendríticas e axosomáticas, hai tamén sinapses axoaxónicas (de axón con axón) e dendrodendríticas (de dendrita con dendrita).
A sinalización sináptica é básica no funcionamento neuronal, e é en parte eléctrica e en parte química. A parte eléctrica depende das propiedades da membrana neuronal. Como toda célula, as neuronas están rodeadas por unha membrana plasmática, constituída por unha bicapa lipídica con proteínas estruturais inseridas nela. Unha bicapa lipídica é un potente illante eléctrico, pero nas neuronas moitas das proteínas de membrana son electricamente activas. Entre elas hai proteínas que forman canles iónicas que permiten que os ións flúan a través das membranas, e bombas iónicas que transportan ións activamente dun lado ao outro da membrana. A maioría das canles iónicas son permeables só para tipos específicos de ións. A apertura dalgunhas canles iónicas está controlada pola voltaxe da membrana, o que quere dicir que poden cambiar entre un estado aberto ou pechado segundo varíe a diferenza de potencial (voltaxe) a través da membrana. Outros son de apertura controlada quimicamente, o que significa que se abren ou pechan segundo interaccionen con substancias químicas que difunden polo fluído extracelular. As interaccións entre as canles iónicas e as bombas iónicas produce unha diferenza de potencial a través da membrana, normalmente de algo menos de 1/10 de voltio inicialmente. Esta voltaxe ten dúas funcións: primeira, proporciona unha fonte enerxética para o conxunto de proteínas de membrana dependentes de voltaxe; segunda, proporciona unha base para a transmisión eléctrica do sinal entre diferentes partes da membrana.
As neuronas comunícanse por medio de sinapses químicas ou sinapses eléctricas nun proceso chamado transmisión sináptica. O proceso fundamental que desencadea a transmisión sináptica é o potencial de acción, un sinal eléctrico que se propaga, e que se xera aproveitando as propiedades de excitabilidade eléctrica da membrana da neurona. Este fenómeno denomínase tamén onda de despolarización da membrana.
Anatomía e histoloxía
As neuronas están moi especializadas no procesamento e transmisión de sinais celulares. Dada a gran diversidade de funcións levadas a cabo polas neuronas en diferentes partes do sistema nervioso, estas presentan unha gran variedade de formas, tamaños e propiedades electroquímicas. Por exemplo, o soma dunha neurona pode variar de 4 a 100 micrómetros de diámetro.[4]
- O soma é onde ten lugar case toda a biosíntese proteica e contén o núcleo celular. O citoplasma do soma chámase pericarion. O núcleo ten de 3 a 18 micrómetros de diámetro.[5]
- As dendritas dunha neurona son extensións celulares con moitas ramificacións, e o seu conxunto denomínase árbore dendrítica. Aquí é onde a neurona recibe a maior parte dos sinais.
- O axón é unha fina proxección con forma de cable cunha lonxitude que pode ser decenas, centos ou decenas de miles de veces maior ca diámetro do soma. O axón transporta os sinais nerviosos fóra do soma (pero tamén leva certo tipo de información na dirección contraria). Moitas neuronas teñen só un axón, pero este axón con frecuencia está moi ramificado, permitindo a comunicación con moitas células distintas. A parte do axón onde este emerxe do soma denomínase protuberancia do axón ou cono axónico. Ademais de ser unha estrutura anatómica, esta protuberancia é tamén a parte da neurona que ten a maior densidade de canles de sodio de apertura dependente de voltaxe. Isto fai que sexa a parte máis doadamente excitable da neurona e a zona onde se orixina o potencial inicial do impulso no axón: en termos electrofisiolóxicos ten o limiar do potencial de acción máis negativo. De modo que, aínda que o axón e a protuberancia do axón están xeralmente implicados na transmisión de información a outras células, estas rexións poden tamén recibir impulsos doutras neuronas.
- O terminal do axón contén as sinapses, estruturas especializadas nas que se liberan os neurotransmisores químicos para comunicarse coas neuronas diana.
Aínda que a idea canónica de neurona atribúe funcións específicas a cada un dos seus compoñentes anatómicos, as dendritas e os axóns a miúdo actúan de xeito contrario ás súas funcións habituais.
Os axóns e as dendritas no sistema nervioso central teñen normalmente só un micrómetro de grosor, pero no sistema nervioso periférico son moito máis grosas. O soma ten uns 10–25 micrómetros de diámetro e xeralmente non é moito máis grande ca o núcleo celular que contén. O axón máis longo dunha motoneurona humana pode ter arredor dun metro de lonxitude, chegando desde a base da espiña dorsal á punta dunha deda do pé. As neuronas sensoriais teñen axóns que van desde as dedas á espiña dorsal e chegan a 1,5 metros de lonxitude nos adultos. As xirafas teñen axóns de varios metros que percorren os seus longos pescozos. Moito do que sabemos do funcionamento dos axóns aprendeuse estudando os axóns xigantes da lura, que é unha preparación experimental ideal debido ao seu enorme tamaño (de 0,5–1 milímetro de grosor, e varios centímetros de longo).
As neuronas completamente diferenciadas non experimentan mitose en absoluto;[6] porén, descubrimentos recentes mostraron que se poden orixinar novas neuronas no cerebro a partir da división de células troncais (ou nais) neurais, que se atopan por todo o cerebro pero especialmente na zona subventricular e na subgranular, proceso chamado neuroxénese.[7]
Histoloxía e estrutura interna
As neuronas tinguidas con colorantes básicos mostran no seu soma numerosas agrupacións microscópicas tinguidas denominadas substancia de Nissl ou corpos de Nissl (chamados así polo psiquiatra e neuropatólogo alemán Franz Nissl, 1860–1919), que constan de retículo endoplasmático rugoso con ARNr asociado. A abundancia de corpos de Nissl pode explicarse polo feito de que as células nerviosas son moi activas metabolicamente, e, por tanto, realizan unha intensa síntese de proteínas.
O corpo celular dunha neurona está sostido por unha complexa rede de proteínas estruturais chamadas neurofilamentos, que se ensamblan en grandes neurofibrilas. Algunhas neuronas tamén conteñen gránulos de pigmentos, como a neuromelanina (un pigmento castaño escuro, subproduto da síntese de catecolaminas) e lipofuscina (un pigmento amarelo-castaño que se acumula coa idade).
Os axóns e as dendritas teñen diferentes características estruturais internas. Os axóns típicos case nunca conteñen ribosomas, excepto tal vez algúns no seu segmento inicial. As dendritas conteñen retículo endoplasmático rugoso ou ribosomas, que diminúen en cantidade coa distancia ao soma celular.
Clases
As neuronas poden ter diferentes formas e tamaños e poden clasificarse pola súa morfoloxía ou función. O anatomista Camillo Golgi agrupou as neuronas en dous tipos: tipo I con longos axóns utilizados para enviar sinais a longas distancias, e tipo II con axóns curtos, que ás veces poden confundirse con dendritas. As de tipo I poden subdividirse tendo en conta onde está situado o seu soma celular. A morfoloxía básica das neuronas de tipo I, representadas polas motoneuronas espiñais, consta dun corpo celular chamado soma e un longo axón cuberto dunha vaíña de mielina. Arredor do corpo celular nacen e ramifícanse as dendritas que reciben os sinais procedentes doutras neuronas. O final do axón ten terminais ramificados (terminais axónicos) que liberan neurotransmisores no espazo entre o final do axón e as dendritas da seguinte neurona chamado espazo sináptico ou fenda sináptica. Por tanto, as neuronas non están en contacto físico para realizar a transmisión do impulso, senón que hai un espazo entre elas, o espazo sináptico.
Clasificación estrutural
Polaridade
A maioría das neuronas poden clasificarse anatomicamente como:
- Unipolar ou pseudounipolar: coas dendritas e o axón que nacen da mesma prolongación ou proceso que emerxe da célula.
- Bipolar: co axón e unha única dendrita que saen de partes opostas do soma.
- Multipolar: con máis de dúas dendritas. Poden ser:
Outros
Ademais, foron identificados algúns outros tipos neuronais únicos de acordo coa súa situación no sistema nervioso e forma distintiva. Por exemplo:
- Células cesta, interneuronas que forman un denso plexo de terminais arredor do soma de células diana, que se encontran no córtex e cerebelo.
- Células de Betz, longas motoneuronas.
- Neuronas espiñais medianas, son a maioría das neuronas do corpo estriado (corpus striatum).
- Células de Purkinje, enormes motoneuronas do cerebelo, que son neuronas multipolares do tipo Golgi I.
- Células piramidais, neuronas con soma triangular do tipo Golgi I.
- Células de Renshaw, neuronas cos seus dous extremos unidos a motoneuronas alfa.
- Células granulares, certas neuronas de tipo Golgi.
- Células da hasta anterior da medula espiñal, motoneuronas localizadas na medula espiñal.
Clasificación funcional
Dirección
- Neuronas aferentes, que transportan a información desde os tecidos e órganos ao sistema nervioso central e chámanse tamén ás veces neuronas sensoriais.
- Neuronas eferentes, que transmiten sinais desde o sistema nervioso central ás células efectoras e chámanse ás veces motoneuronas.
- Interneuronas, que conectan neuronas en determinadas zonas do sistema nervioso central.
O termo aferente e eferente tamén se refire xeralmente a neuronas que, respectivamente, traen información ou envían información da rexión do cerebro.
Acción sobre outras neuronas
Unha neurona actúa sobre outras neuronas liberando neurotransmisores que se unen a receptores de membrana. O efecto sobre a neurona diana está determinado non polo tipo de neurona presináptica nin polo neurotransmisor liberado, senón polo tipo de receptor que é activado. Un neurotransmisor pode entenderse como unha chave, e o seu receptor como a pechadura: pero no sistema nervioso o mesmo tipo de chave pode usarse para abrir moitos tipos diferentes de pechaduras. Os receptores poden clasificarse grosso modo como excitadores (que causan un incremento da frecuencia de envío dos impulsos), e inhibidores (que causan un decrecemento da frecuencia de envío dos impulsos), ou moduladores (que causan efectos a longo prazo non relacionados directamente coa frecuencia de envío de impulsos).
En realidade, cada un dos dous neurotransmisores máis comúns no cerebro, o glutamato e o GABA, ten unha acción case sempre do mesmo tipo, sempre excitatoria o glutamato, e sempre inhibitoria o GABA. Aínda que o glutamato actúa sobre diferentes tipos de receptores, a maioría deles teñen efectos excitatorios. De xeito similar o GABA actúa sobre diferentes tipos de receptores, pero todos eles teñen efectos inhibitorios (en animais adultos, polo menos). Debido a esta constancia na súa acción, é moi común que os neurocientíficos simplifiquen a terminoloxía e falen das células que liberan glutamato como "neuronas excitatorias," e das células que liberan GABA como "neuronas inhibitorias." Como arredor do 90% das neuronas do cerebro liberan ou ben glutamato ou ben GABA, estas denominacións abranguen a maior parte das neuronas. Hai tamén outros tipos de neuronas con efectos sempre do mesmo tipo sobre as súas dianas, por exemplo as motoneuronas "excitatorias" da medula espiñal que liberan acetilcolina, e as neuronas espiñais "inhibitorias" que liberan glicina.
Non obstante, a distinción entre neurotransmisores excitatorios e inhibitorios non é absoluta. Máis ben depende da clase de receptores químicos presentes na neurona diana. En principio, unha determinada neurona, liberando un determinado neurotransmisor, pode ter efectos excitatorios sobre determinadas células diana, inhibitorios sobre outras, e modulatorios sobre outras. Por exemplo, as células fotorreceptoras da retina liberan sempre o neurotransmisor glutamato en ausencia de luz. As denominadas células bipolares OFF da retina son, como a maioría das neuronas, excitadas pola liberación de glutamato. Porén, as neuronas diana veciñas, chamadas células bipolares ON da retina son polo contrario inhibidas polo glutamato, porque carecen do receptor de glutamato ionotrópico normal (cando se activa abre directamente algunha canle iónica) e no seu lugar posúen un receptor de glutamato metabotrópico inhibitorio (cando se activa utiliza un segundo mensaxeiro para abrir a canle iónica).[9] Cando hai luz, os fotorreceptores deixan de liberar glutamato, o cal fai que deixe de haber inhibición nas células bipolares ON, que se activan; e isto simultaneamente elimina a excitación das células bipolares OFF, silenciándoas.
Patróns de descarga
As neuronas poden clasificarse de acordo coas súas características electrofisiolóxicas en:
- Tónicas ou de picos regulares. Algunhas neuronas están constantemente (ou tonicamente) activas. Exemplo: as interneuronas do neurostriatum. Envían picos de potencial eléctrico (impulsos) constantemente.
- Fásicas ou explosivas. Estas neuronas envían impulsos só en determinados momentos e chámanse fásicas.
- De picos rápidos. Algunhas neuronas envían impulsos cunha grande frecuencia de disparo, por exemplo algúns tipos de interneuronas inhibitorias corticais, as células do globo pálido (globus pallidus), as células ganglionares da retina.[10][11]
Clasificación polo tipo de neurotransmisor producido
As neuronas diferéncianse tamén polo tipo de neurotransmisor que producen. Algúns exemplos son:
- Neuronas colinérxicas. Liberan acetilcolina. A neurona presináptica libera acetilcolina na fenda sináptica. A acetilcolina actúa como un ligando tanto para canles iónicas de apertura por unión a un ligando coma para receptores muscarínicos metabotrópicos (GPCRs). Os receptores nicotínicos son canles iónicas de apertura por unión a un ligando pentaméricas compostas de subunidades alfa e beta que se unen á nicotina. A unión do ligando abre a canle causando despolarización por entrada de Na+ e incrementando a probabilidade da liberación dos neurotransmisores presinápticos.
- Neuronas GABAérxicas. Liberan ácido gamma aminobutírico. O GABA (ácido gamma aminobutírico) é un dos dous neuroinhibidores do sistema nervioso central; o outro é a glicina. O GABA ten unha función homóloga á da acetilcolina, abrindo canles aniónicas que permiten ao Cl- entrar na neurona postsináptica. Este ión causa hiperpolarización na neurona, diminuíndo a probabilidade de que se dispare o potencial de acción a medida que a voltaxe se fai máis negativa (lembremos que para que un potencial de acción se dispare, debe acadarse un limiar de voltaxe positivo).
- Neurona glutamatérxicas. Liberan glutamato. O glutamato é un dos dous aminoácidos excitadores primarios; o outro é o aspartato. Os receptores do glutamato forman unha das catro categorías que hai, tres das cales son canles iónicas de apertura por unión a un ligando e a outra é o receptor acoplado á proteína G (ou GPCR).
- Receptores AMPA, que funcionan como canles catiónicas permeables ao catión Na+ que median a tranmisión sináptica excitatoria rápida.
- Receptores de cainato, que funcionan igual ca os anteriores.
- Receptores NMDA, que son outra canle catiónica que é máis permeable ao Ca2+. A función dos receptores depende de que o receptor da glicina se una como co-agonista co poro da canle. Os receptores NMDA non funcionan sen que estean presentes ambos os ligandos.
- Receptores metabotrópicos, son os GPCRs que modulan a transmisión sináptica e a excitabilidade postsináptica.
- O glutamato pode causar exotoxicidade cando se interrompe o fluxo sanguíneo no cerebro, o que orixina un dano cerebral. Cando se suprime o fluxo sanguíneo, libérase glutmato das neuronas presinápticas causando unha activación dos receptores NMDA e AMPA maior da que sería normal, o que leva a un incremento da entrada de Ca2+ e Na+ na neurona postsináptica e a un dano celular.
- Neuronas dopaminérxicas. Liberan dopamina. A dopamina é un neurotransmisor que actúa nos receptores acoplados a Gs de tipo D1 (D1 e D5), que incrementan o AMPc e a proteína quinase A (PKA), e nos receptores de tipo D2 (D2, D3, e D4), que activan os receptores acoplados a Gi, que diminúen o AMPc e a PKA. A dopamina está relacionada cos estados de humor e o comportamento, e modula tanto a neurotransmisión presináptica coma postsináptica. A perda das neuronas dopaminérxicas na substancia negra foi relacionada coa enfermidade de Parkinson.
- Neuronas serotoninérxicas. Liberan serotonina. A serotonina, (5-hidroxitriptamine, 5-HT), pode actuar como excitatoria ou inhibitoria. das catro clases de receptores 5-HT que existen, tres son receptores GPC e unha é unha canle iónica de apertura por unión a un ligando. A serotonina sintetízase a partir do triptófano pola triptófano hidroxilase, e pola ácido aromático descarboxilase. A falta de serotonina nas neuronas postsinápticas está asociada coa depresión. As drogas que bloquean o transportador de serotonina presináptico úsanse para o seu tratamento, como o Prozac e Zoloft.
No sistema nervioso vexetativo as neuronas son principalmente ou colinérxicas ou adrenérxicas (liberan noradrenalina).
Conectividade
- Artigo principal: Sinapse.
As neuronas communícanse unhas con outras por medio de sinapses, nas cales o terminal do axón ou os botóns terminais situados nos extremos das ramificacións do axón, dunha neurona (presináptica) actúan sobre as dendritas, soma ou menos comunmente o axón da neurona seguinte (postsináptica). Por tanto, existen sinapses axo-dendríticas, axo-somáticas ou axo-axónicas. Algunhas neuronas como as células de Purkinje do cerebelo poden ter unhas 1000 ramas dendríticas, e establecen conexións con decenas ou miles de células; pero outras neuronas, como as neuronas magnocelulares dos núcleos supraópticos, teñen só unha ou dúas dendritas, cada unha das cales recibe miles de sinapses. As sinapses poden ser excitatorias ou inhibitorias e poden incrementar ou diminuír a actividade na neurona postsináptica. Nas sinapses vértense neurotransmisores químicos, polo que se denominan sinapses químicas, que son con gran diferenza as máis abundantes. Pero algunhas neuronas tamén se comunican por medio de sinapses eléctricas, que son unións comunicantes (gap junctions) directas e electricamente condutivas nas que as células están en estreito contacto.
Nunha sinapse química, o proceso de transmisión sináptica é o seguinte: cando un potencial de acción chega ao terminal do axón, ábrense canles de calcio de apertura dependente da voltaxe, que permiten que o Ca2+ do exterior entre no terminal. O calcio causa que as vesículas sinápticas cheas de neurotransmisores se fusionen coa membrana celular, liberando o seu contido na fenda sináptica. Os neurotransmisores difunden a través da fenda sináptica, únense aos seus receptores da membrana da neurona postsináptica e actívanos, o que causará a despolarización de dita membrana.
O cerebro humano ten un enorme número de sinapses. Cada unha das 1011 neuronas do cerebro ten como media 7.000 conexións sinápticas con outras neuronas. Estimouse que no cerebro dun rapaz de 3 anos hai unhas 1015 sinapses. Este número decrece conforme o rapaz vai cumprindo anos, estabilizándose na idade adulta. As estimacións varían para o adultos, e van desde 1014 a 5 x 1014 sinapses en total.[12]
Mecanismos de propagación dos potenciais de acción
En 1937, John Zachary Young suxeriu que se podía usar o axón xigante da lura para estudar as propiedades eléctricas da neurona.[13] Como son de natureza similar ao axón humano pero moito máis grandes, as neuronas da lura son moito máis fáciles de estudar. Poden inserirse eléctrodos nos axóns xigantes da lura, e medir con precisión os potenciais de membrana.
A membrana celular do axón e do soma contén canles iónicas de apertura por voltaxe que permiten que a neurona xere e propague un impulso eléctrico (un potencial de acción). Estes impulsos son xerados e propagados por ións, que son os que levan as cargas eléctricas, como o sodio (Na+), potasio (K+), cloruro (Cl-), e calcio (Ca2+).
Hai varios estímulos que poden activar unha neurona xerando unha actividade eléctrica, entre os que están: presión, estiramento, transmisores químicos, e cambios no potencial eléctrico a través da membrana.[14] Os estímulos provocan a apertura de canles iónicas específicas na membrana, que orixina un fluxo de ións a través da membrana, que cambia o potencial de membrana.
As neuronas e axóns delgados requiren menos gasto metabólico para producir e transportar os potenciais de acción, pero os axóns de maior grosor transportan os impulsos con maior rapidez. Para minimizar o gasto metabólico mantendo unha condución rápida, moitas neuronas illaron os seus axóns cunha vaíña de mielina que os envolve. As vaíñas están formadas por células de glia: oligodendrocitos no sistema nervioso central, e células de Schwann nos nervios do sistema nervioso periférico. A vaíña permite que os potenciais de acción viaxen máis rápido ca nos axóns non mielinizados do mesmo diámetro, á vez que consomen menos enerxía. A vaíña de mielina dos nervios periféricos normalmente cobre o axón en seccións de aproximadamente 1 mm de longo, separadas por curtas zonas non cubertas chamadas nódulos de Ranvier, que conteñen unha gran densidade de canles iónicas de apertura por voltaxe. A esclerose múltiple é unha doenza neurolóxica causada pola desmielinización dos axóns do sistema nervioso central.
Algunhas neuronas non xeran potenciais de acción, senón que no seu lugar xeran un impulso eléctrico gradual, que á súa vez causa unha liberación de neurotransmisores tamén gradual. Estas neuronas que non teñen un pico no potencial de acción adoitan ser neuronas sensoriais ou interneuronas, porque non poden enviar impulsos a longas distancias.
Codificación neural
A codificación neural trata sobre como as neuronas representan no cerebro a información sensorial ou doutro tipo. O principal obxectivo do estudo da codificación neural é caracterizar as relacións entre o estímulo e as respostas neuronais individuais ou de conxunto, e as relacións entre as actividades eléctricas das neuronas e as do conxunto.[15] Pénsase que as neuronas poden codificar tanto información dixital coma analóxica.[16]
Principio de todo ou nada
A condución de impulsos nerviosos é un exemplo de resposta de "todo ou nada". Noutras palabras, cando as neuronas responden a todo, a súa resposta debe ser completa, máxima. Unha maior intensidade na estimulación non produciría un impulso maior, pero si pode producir máis impulsos por segundo (aumenta a frecuencia de impulsos, non a súa intensidade). Hai diferentes tipos de respostas do receptor ao estímulo. Os receptores adaptativos lentos (ou tónicos) responden a estímulos estables e producen unha frecuencia de envío de impulsos (de "disparo") estable. Estes receptores tónicos xeralmente responden a un incremento na intensidade do estímulo co incremento da súa frecuencia de envío de impulsos.
Pero hai tamén outros tipos de receptores denominados adaptativos rápidos (ou fásicos), nos que o seu envío de impulsos decrece ou para cando reciben estímulos estables. Un exemplo: cando a pel humana é tocada por un obxecto, isto causa que as neuronas da pel envíen impulsos, pero se o obxecto mantén a súa presión contra a pel, as neuronas deixan de enviar impulsos. As neuronas da pel e músculos que responden á presión e vibración teñen estruturas de filtrado accesorias que colaboran na súa función.
O corpúsculo de Pacini é unha desas estruturas. Consta de varias capas concéntricas como as dunha cebola, que se forman arredor do terminal do axón. Cando se aplica a presión e o corpúsculo se deforma, o estímulo mecánico transfírese ao axón, que envía impulsos. Se a presión é constante, non hai xa máis estímulos; deste modo, tipicamente estas neuronas responden cunha despolarización transitoria durante a deformación inicial e responden de novo cando desaparece a presión, xa que nese caso o corpúsculo cambia de forma outra vez. Existen máis tipos de adaptación, que lle dan ás neuronas distintas características funcionais.[17]
Historia
O termo neurona acuñouno o anatomista alemán Heinrich Wilhelm Waldeyer. O papel da neurona como unidade funcional primaria do sistema nervioso foi recoñecida por primeira vez a principios do século XX grazas aos traballos do anatomista español Santiago Ramón y Cajal.[18] Cajal propuxo que as neuronas eran células separadas que se comunicaban unhas con outras por medio de unións especializadas que deixaban un espazo entre as células. Non eran, pois, células que estaban fusionadas polas súas prolongacións formando unha rede continua, que era outra teoría da época defendida por Golgi.[18] Isto chegou a coñecerse como a doutrina neuronal, un dos principios centrais da neurociencia moderna.[18] Para observar a estrutura de neuronas individualizadas, Cajal mellorou o proceso de tinguido con sales de prata coñecido por método de Golgi, que fora desenvolvido polo seu rival, Camillo Golgi.[18] A mellora introducida por Cajal, que consistía nunha técnica chamada "dobre impregnación", aínda se usa. A tinguidura con impregnación de prata é un método extraordinariamente útil en investigacións neuroanatómicas porque, por razóns aínda descoñecidas, tingue un pequeno porcentaxe das células dun tecido, de modo que é posible ver a microestrutura completa de neuronas individualizadas sen que se solapen moito con outras células de tecidos densamente empaquetados como os do cerebro.[19]
A doutrina neuronal
A doutrina neuronal é a idea fundamental actual de que as neuronas son as unidades básicas estruturais e funcionais do sistema nervioso. A teoría foi presentada por Santiago Ramón y Cajal a finais do século XIX. Sostén que as neuronas son células individualizadas (non conectadas formando unha rede), que actúan como unidades metabólicas diferenciadas.
Posteriores descubrimentos engadiron máis refinamentos á formulación simple inicial desta doutrina. Por exemplo, as células de glia, que non se consideran neuronas, xogan un papel esencial no procesamento da información.[20] Ademais, as sinapses eléctricas son máis comúns do que inicialmente se pensaba,[21] o que significa que si hai algunhas conexións directas e citoplasmáticas entre as neuronas. De feito, hai exemplos de neuronas que se unen aínda máis estreitamente: o axón xigante da lura orixínase pola fusión de múltiples axóns.[22]
Cajal tamén postulou a lei da polarización dinámica, que establece que unha neurona recibe sinais polas súas dendritas e corpo celular e transmíteos como potenciais de acción a través do axón nunha soa dirección: saíndo do corpo celular.[23] A lei da polarización dinámica ten importantes excepcións; as dendritas poden servir como saídas sinápticas das neuronas[24] e os axóns poden recibir sinais sinápticos.
As neuronas no cerebro
O número de neuronas no cerebro varía enormemente dunha especie a outra.[25] Unha estimación sobre o número de neuronas do cerebro humano dá unha cifra de 100 mil millóns ( ) de neuronas e 100 billóns ( ) de sinapses.[25] Outras estimacións falan de 86 mil millóns de neuronas, das cales 16,3 miles de millóns están no córtex cerebral, e 69 miles de millóns no cerebelo.[26] En comparación, o verme nematodo Caenorhabditis elegans ten só 302 neuronas, o que fai del un suxeito de experimentación ideal no que os científicos puideron mapear todas as neuronas do seu organismo. A mosca da froita Drosophila melanogaster, outro animal de experimentación habitual, ten arredor de 100.000 neuronas e realiza moitos comportamentos complexos. Moitas propiedades das neuronas, desde o tipo de neurotransmisores que usan ata a composición das súas canles iónicas, son similares en distintas especies, o que permite aos científicos estudar os procesos que ocorren nos organismos complexos experimentando con organismos máis simples.
Trastornos neurolóxicos
- Artigo principal: Neuroloxía.
Son numerosos os trastornos neurolóxicos producidos por danos ou alteracións no funcionamento das neuronas. Algúns exemplos son: A síndrome de Charcot-Marie-Tooth é un trastorno hereditario heteroxéneo dos neuvios (neuropatía) que se caracteriza nos estadios avanzados da doenza pola perda de tecido muscular e da sensación táctil, predominantemente nos pés e pernas pero tamén nas mans e brazos. Actualmente é incurable. É un dos trastornos neurolóxicos hereditarios máis comúns, cunha frecuencia de aparición duns 37 casos cada 100.000 persoas.
A enfermidade de Alzheimer, é unha enfermidade neurodexenerativa caracterizada pola progresiva deterioración cognitiva xunto coa diminución da habilidade para realizar as actividades da vida diaria e síntomas neuropsiquiátricos ou cambios no comportamento. O síntoma inicial máis chocante é a perda de memoria a curto prazo (amnesia), que se manifesta normalmente co esquecemento de pequenas cousas, que se fai cada vez máis pronunciado conforme avanza a doenza, acompañado dunha relativa conservación das lembranzas antigas de anos atrás. Conforme o trastorno progresa, a discapacidade intelectual aumenta afectando á linguaxe (afasia), movementos complicados (apraxia), recoñecemento (agnosia), e a funcións como a toma de decisións e a realización de plans.
A enfermidade de Parkinson, é un trastorno dexenerativo crónico e progresivo do sistema nervioso central que a miúdo afecta ás habilidades motoras e á fala do paciente. A enfermidade de Parkinson pertence a un grupo de doenzas que orixinan trastornos do movemento. Caracterízase pola rixidez muscular, tremores, movementos físicos lentos (bradiquinesia), e en casos extremos, perda da capacidade de movemento (aquinesia). Os síntomas principais son o resultado da diminución da estimulación do córtex motor polos ganglios basais, normalmente causada pola insuficiente formación e actividade da dopamina, que se produce nas neuronas dopaminérxicas do cerebro. Os síntomas secundarios poden incluír disfuncións cognitivas e problemas sutís de linguaxe.
A miastenia gravis é unha doenza neuromuscular que produce unha debilidade muscular flutuante e cansazo ao realizar actividades simples. A debilidade está causada por anticorpos circulantes que bloquean os receptores de acetilcolina na unión neuromuscular postsináptica, que inhiben o efecto estimulante do neurotransmisor acetilcolina, polo que é unha doenza autoinmunitaria. A miastenia trátase con inmunosupresores, inhibidores da colinesterase e, en determinados casos, coa extirpación do timo.
Desmielinización
A desmielinización consiste en que os nervios perden a súa vaíña de mielina illante. Cando se degrada a mielina, a condución de impulsos ao longo dos nervios pode verse afectada ou perderse, e e o nervio finalmente dexenera. Isto causa certos trastornos neurodexenerativos como esclerose múltiple e polirradiculoneuropatía desmielinizante inflamatoria crónica (doenza autoinmune).
Dexeneración dos axóns
A maioría das respostas aos danos nas neuronas consisten nun fluxo sinalizador de calcio que promove o selado das partes rotas da neurona, pero os danos nos axóns inicialmente causan unha aguda dexeneración axonal, que é a rápida separación entre os extremos distal e proximal do axón uns 30 minutos despois do dano. A dexeneración vai seguida do inchado da membrana do axón (axolema) , e posteriormente a formación dunha especie de gránulos. Despois da degradación do axolema ten lugar unha disintegración granular do citoesqueleto do axón e dos orgánulos do seu interior. Os primeiros cambios que se observan son a acumulación de mitocondrias nas rexións paranodais no sitio do dano. O retículo endoplasmático degrádase e as mitocondrias inchan e finalmente desintégranse. A desintegración é dependente da ubiquitina e de calpaína proteases, e está causada por un fluxo de ión calcio, o que suxire que a dexeneración axonal é un proceso activo. Deste modo, o axón sofre unha completa fragmentación. O proceso leva unhas 24 horas no sistema nervioso periférico e é máis longo no sistema nervioso central. As vías de sinalización que levan á dexeneración do axolema non se coñecen actualmente.
Rexeneración de nervios
Está demostrado que a neuroxénese pode ás veces ocorrer no cerebro dos vertebrados adultos, o que produciu unha certa controversia en 1999.[27] Pero os estudos máis recentes da idade das neuronas humanas suxiren que este proceso ocorre só nunha minoría das neuronas, e a inmensa maioría das neuronas, incluídas as do neocórtex, fórmanse antes do nacemento e persisten durante toda a vida sen seren substituídas.[28]
O que si é posible é a recrecemento de axóns periféricos que foron cortados. Este recrecemento pode levarlles moito tempo: despois dun dano nos nervios do brazo humano, por exemplo, pode tardarse meses en volver a sentir o tacto de mans e dedos. Un informe publicado en Nature suxire que se descubriu unha forma de transformar células da pel humana en células nerviosas funcionais usando un proceso chamado transdiferenciación, no cal as "células son forzadas a adoptar novas identidades".[29]
Notas
- ↑ "Henry George Liddell, Robert Scott, A Greek-English Lexicon, νεῦρον".
- ↑ Myriam Cayre, Jordane Malaterre, Sophie Scotto-Lomassese, Colette Strambi e Alain Strambi. The common properties of neurogenesis in the adult brain: from invertebrates to vertebrates Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology. Volume 132, número 1, maio 2002, páxinas 1-15.
- ↑ Paniagua, R.; Nistal, M.; Sesma, P.; Álvarez-Uría, M.; Fraile, B.; Anadón, R. e José Sáez, F. (2002). Citología e histología vegetal y animal. McGraw-Hill Interamericana de España, S.A.U. ISBN 84-486-0436-9.
- ↑ Davies, Melissa (2002-04-09). "The Neuron: size comparison". Neuroscience: A journey through the brain. Consultado o 2009-06-20.
- ↑ Chudler, Eric H. "Brain Facts and Figures". Neuroscience for Kids. Consultado o 2009-06-20.
- ↑ Herrup K, Yang Y (2007). "Cell cycle regulation in the postmitotic neuron: oxymoron or new biology?". Nat. Rev. Neurosci. 8 (5): 368–78. PMID 17453017. doi:10.1038/nrn2124.
- ↑ Alvarez-Buylla A, Garcia-Verdugo JM (February 1, 2002). "Neurogenesis in adult subventricular zone". The Journal of Neuroscience : the Official Journal of the Society for Neuroscience 22 (3): 629–34. PMID 11826091. Consultado o 2009-06-20.
- ↑ http://biology.plosjournals.org/perlserv/?request=get-document&doi=10.1371/journal.pbio.0040029
- ↑ Gerber U (2003). "Metabotropic glutamate receptors in vertebrate retina". Doc Ophthalmol 106 (1): 83–87. PMID 12675489. doi:10.1023/A:1022477203420.
- ↑ Kolodin, YO; Veselovskaia, NN; Veselovsky, NS; Fedulova, SA. Ion conductances related to shaping the repetitive firing in rat retinal ganglion cells. Acta Physiologica Congress. Consultado o 2009-06-20.
- ↑ Ionic conductances underlying excitability in tonically firing retinal ganglion cells of adult rat
- ↑ Drachman D (2005). "Do we have brain to spare?". Neurology 64 (12): 2004–5. PMID 15985565. doi:10.1212/01.WNL.0000166914.38327.BB.
- ↑ Chudler, Eric H. "Milestones in Neuroscience Research". Neuroscience for Kids. Consultado o 2009-06-20.
- ↑ Patlak, Joe; Gibbons, Ray (2000-11-01). "Electrical Activity of Nerves". Aps in Nerve Cells. Consultado o 2009-06-20.
- ↑ Brown EN, Kass RE, and Mitra PP. 2004. Multiple neural spike train data analysis: state-of-the-art and future challenges. Nature Neuroscience 7:456-61
- ↑ Spike arrival times: A highly efficient coding scheme for neural networks, SJ Thorpe – Parallel processing in neural systems, 1990
- ↑ Eckert, Roger; Randall, David (1983). Animal physiology: mechanisms and adaptations. San Francisco: W.H. Freeman. p. 239. ISBN 978-0716718284.
- ↑ 18,0 18,1 18,2 18,3 López-Muñoz, F.; Boya, J., Alamo, C. (16 October 2006). "Neuron theory, the cornerstone of neuroscience, on the centenary of the Nobel Prize award to Santiago Ramón y Cajal". Brain Research Bulletin 70 (4-6): 391–405. PMID 17027775. doi:10.1016/j.brainresbull.2006.07.010. Consultado o 2007-04-02.
- ↑ Grant, Gunnar; Boya, J; Alamo, C (9 January 2007 (online)). "How the 1906 Nobel Prize in Physiology or Medicine was shared between Golgi and Cajal". Brain Research Reviews 55 (4-6): 490. PMID 17306375. doi:10.1016/j.brainresrev.2006.11.004. Consultado o 2007-04-02.
- ↑ Witcher M, Kirov S, Harris K (2007). "Plasticity of perisynaptic astroglia during synaptogenesis in the mature rat hippocampus". Glia 55 (1): 13–23. PMID 17001633. doi:10.1002/glia.20415.
- ↑ Connors B, Long M (2004). "Electrical synapses in the mammalian brain". Annu Rev Neurosci 27: 393–418. PMID 15217338. doi:10.1146/annurev.neuro.26.041002.131128.
- ↑ R. W. Guillery, Observations of synaptic structures: origins of the neuron doctrine and its current status, Phil. Trans. R. Soc. B (2005) 360, 1281–1307 (p 1300).
- ↑ Sabbatini R.M.E. April–July 2003. Neurons and Synapses: The History of Its Discovery. Brain & Mind Magazine, 17. Retrieved on March 19, 2007.
- ↑ Djurisic M, Antic S, Chen W, Zecevic D (2004). "Voltage imaging from dendrites of mitral cells: EPSP attenuation and spike trigger zones". J Neurosci 24 (30): 6703–14. PMID 15282273. doi:10.1523/JNEUROSCI.0307-04.2004.
- ↑ 25,0 25,1 Williams RW, Herrup K (1988). "The control of neuron number". Annual Review of Neuroscience 11: 423–53. PMID 3284447. doi:10.1146/annurev.ne.11.030188.002231.
- ↑ Azevedo FA, Carvalho LR, Grinberg LT; et al. (2009). "Equal numbers of neuronal and nonneuronal cells make the human brain an isometrically scaled-up primate brain". The Journal of Comparative Neurology 513 (5): 532–41. PMID 19226510. doi:10.1002/cne.21974.
- ↑ Brain may grow new cells daily, New York Times 1999.
- ↑ Stable neuron numbers from cradle to grave. PNAS.
- ↑ Ewen Callaway (26 May 2011). "How to make a human neuron". NatureNews. Consultado o 2011-05-24.
Transformando células da pel humana en células nerviosas funcionais, os investigadores poden ter ideado un modelo para as enfermidades do sistema nervioso e quizais para terapias rexeneratias baseadas en transplantes de células. Este logro, do que se informa online hoxe en Nature, é o último no campo en rápido desenvolvemento da chamada transdiferenciación, no cal as células son forzadas a adoptar novas identidades. O pasado ano, os investigadores converteron células do tecido conectivo que se encontran na pel en células cardíacas, células do sangue e do fígado.
Véxase tamén
| Wikimedia Commons ten máis contidos multimedia na categoría: Neurona |
Bibliografía
- Kandel E.R., Schwartz, J.H., Jessell, T.M. 2000. Principles of Neural Science, 4th ed., McGraw-Hill, New York.
- Bullock, T.H., Bennett, M.V.L., Johnston, D., Josephson, R., Marder, E., Fields R.D. 2005. The Neuron Doctrine, Redux, Science, V.310, p. 791-793.
- Ramón y Cajal, S. 1933 Histology, 10th ed., Wood, Baltimore.
- Roberts A., Bush B.M.H. 1981. Neurones Without Impulses. Cambridge University Press, Cambridge.
- Peters, A., Palay, S.L., Webster, H, D., 1991 The Fine Structure of the Nervous System, 3rd ed., Oxford, New York
Ligazóns externas
- IBRO (International Brain Research Organization). Fomento da investigación en neurociencia especialmente en países con menor financiamento.
- NeuronBank unha ferramenta online para a catalogación de tipos de neuronas e conectividades sinápticas.
- Imaxes neuroanatómicas de alta resolución de cerebros de primates e non primates.
- O Department of Neuroscience da Wikiversity, ofrece actualmente dous cursos de: Fundamentos das of Neurociencias e Neurociencias comparativas.
- NIF Search – Renshaw Cell via the Neuroscience Information Framework.
- Cell Centered Database – Neuron
- NeuroMorpho.Org base de datos online de reconstrucións dixitais da morfoloxía neuronal.
- Immunohistochemistry Image Gallery: Neuron.