Migración das aves

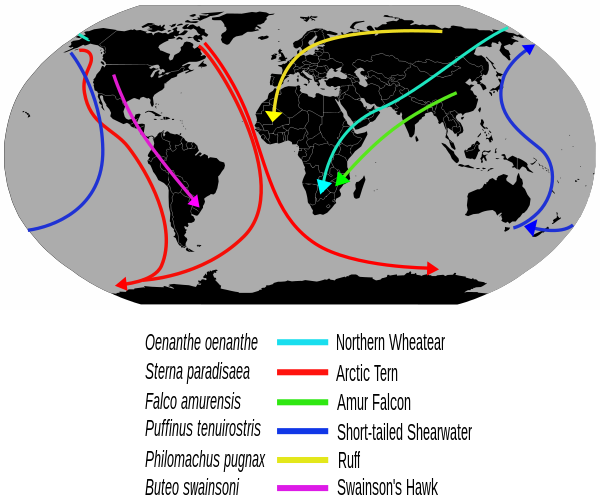
A migración das aves é o movemento regular estacional que fan moitas especies de aves, xeralmente na dirección norte-sur/sur-norte seguindo unha ruta de voo, entre os seus territorios de cría e os de invernada. Moitas especies de aves migran, o que lles supón un custo en predación e mortalidade, incluíndo a caza polos humanos. A migración está impulsada principalmente pola dispoñibilidade de comida, aínda que tamén inflúen outros factores. Ocorre principalmente no hemisferio norte, onde as aves canalizan a súa migración por rutas específicas determinadas por barreiras naturais como o mar Mediterráneo ou o Caribe.
A migración de especies como cegoñas, rulas, e andoriñas foi rexistrada desde hai máis de 3 000 anos por autores da Antiga Grecia, como Homero e Aristóteles e no Libro de Xob da Biblia. Máis recentemente, Johannes Leche empezou a rexistrar os datos das chegadas de migrantes primaverais en Finlandia en 1749, e os modernos estudos científicos utilizan técnicas como o anelamento de aves e o rastreo por satélite para coñecer o movemento das aves migrantes. As ameazas que afrontan as aves migratorias aumentaron coa destrución de hábitats especialmente nos sitios de parada e de invernada, e coa proliferación de estruturas como liñas de alta tensión e muíños para producir enerxía eólica.
O carrán ártico ten a mellor marca de migración a longa distancia, xa que viaxa entre os seus territorios de cría no Ártico e Antártico todos os anos. Algunhas especies de aves con tubos nasais (Procellariiformes) como os albatros circundan a Terrra, voando sobre os océanos do sur, mentres que outras como as pardelas furabuchos migran 14 000 km entre os seus territorios de cría do norte e os océanos do sur. As migracións máis curtas son moi comúns, entre elas están as migracións altitudinais en montañas como os Andes e os Himalaias.
O momento en que se produce a migración parece que está controlado principalmente polos cambios na duración do día. As aves migrantes navegan usando pistas celestes como a posición do Sol e as estrelas, e tamén usan o campo magnético terrestre e mapas mentais.
Perspectiva histórica
[editar | editar a fonte]Rexistrouse o fenómeno da migración das aves desde hai máis de 3 000 anos na Antiga Grecia, onde escritores como Hesíodo, Homero, Heródoto e Aristóteles a mencionaron.[1] Na Biblia tamén se fai mención destas migracións, como no Libro de Xob,[2] onde se fai esta pregunta: "Es ti quen lle ensina a voar ao falcón, tendendo as ás no vento do sur?". O autor do Libro de Xeremías escribiu: "As cegoñas do ceo coñecen todas o tempo da súa volta, as rulas, as andoriñas e os grous volven no seu tempo".[3]
Aristóteles decatouse que os grous viaxaban desde as estepas de Escitia aos pantanos e nacemento do Nilo. Plinio o Vello, na súa Historia Naturalis, repite as observacións de Aristóteles.[4]
Migración fronte a hibernación das andoriñas
[editar | editar a fonte]
Porén, Aristóteles suxeiru que as andoriñas e outras aves hibernaban (quedaban aletargadas no lugar e non migraban). Esta crenza persistiu ata 1878, cando Elliott Coues compilou os títulos de non menos de 182 publicacións que trataban da hibernación das andoriñas. Incluso o "moi observador"[5] Gilbert White, na súa obra publicada postumamente en 1789 The Natural History of Selborne (A historia natural de Selborne), citaba a historia contada por un home de que as andoriñas foran atopadas no derrubamento dun cantil de greda "cando el era un escolar en Brighthelmstone", aínda que o home negou ser testemuña visual.[6] Porén, tamén escribiu que "en canto a que as andoriñas fosen atopadas en estado de letargo durante o inverno na illa de Wight ou en calquera parte deste país, nunca oín un relato dese tipo que pagase a pena escoitar",[6] e que se as andoriñas temperás "atopan casualmente xeo e neve, inmediatamente se retiran durante un certo tempo, unha circunstancia que vai máis en favor de que se esconden que da hibernación", polo que dubida "que volvan por unha semana ou dúas a latitudes máis cálidas".[7]
Ata finais do século XVIII non se aceptou que a migración como explicación da desaparición durante o inverno de aves dos climas do norte.[4] Na obra de Thomas Bewick A History of British Birds (Unha historia das aves británicas), Volume 1, 1797, menciona un informe dun "capitán de barco moi intelixente" que, "entrre as illas de Menorca e Mallorca, viu un gran número de andoriñas voando cara ao norte",[8] e presenta a situación nas illas Británicas da seguinte maneira:

As andoriñas frecuentemente póusanse de noite, despois de que empezan a congregarse, polas beiras de ríos e lagoas, e por tales cirunstancias supúxose erradamente que se retiraban na auga.Bewick[9]
Bewick describe despois un experimento que conseguiu manter anduriñas vivas nas illas Británicas durante varios anos, onde permanecían quentes e secas durante os invernos. Conclúe que:
Estes experimentoss foron desde entón amplamente confirmados por ... M. Natterer, de Viena ... e o resultado proba claramente, o que de feito é agora admitido por todos, que as andoriñas non difiren en ningún caso material doutras aves na súa natureza e tendencias [para a vida no aire]; senón que nos abandonan cando este país xa non pode proporcionarlles unha subministración da súa comida apropiada e natural ...Bewick[10]
Pfeilstörche (cegoñas con frechas cravadas)
[editar | editar a fonte]En 1822, atopouse unha cegoña branca no estado alemán de Mecklenburg cunha frecha cravada feita de madeira de África central, o que era unha das primeiras evidencias da migración de gran distancia que fan as cegoñas.[11][12][13] Esta ave foi denominada en alemán Pfeilstorch, que significa 'cegoña con frecha' (cravada). Desde entón, documentáronse unhas 25 Pfeilstörche.
Padróns xerais
[editar | editar a fonte]

A migración é o movemento estacional regular, xeralmente na dirección norte a sur e viceversa, que realizan moitas especies de aves. O movemento migratorio das aves pode deberse a cambios na dispoñibilidade de alimentos, cambios no hábitat ou no tempo meteorolóxico. Ás veces, estas viaxes non se denominan "verdadeiras migracións" porque son irregulares (nomadismo, invasións, irrupcións) ou só nunha dirección (dispersión, movemento das crías que se van da súa área natal). A migración está marcado pola súa estacionalidade anual.[14] As aves non migratorias dise que son residentes ou sedentarias. Aproximadamente 1 800 das 10 000 especies de aves do mundo son migrantes de longas distancias.[15][16]
Moitas poboacións de aves migran a gran distancia seguindo unha ruta de voo. O padron máis común, no hemisferio norte, consiste en voar ao norte en primavera para reproducirse no verán temperado do Ártico e regresar en outono aos territorios de invernada nas rexións cálidas do sur. Por suposto, no hemisferio sur as direccións son as contrarias, pero hai menos superficie de terra no distante sur para soster unha migración de longa distancia.[17]
A principal motivación para a migración parece ser a procura de alimento; por exemplo, algúns colibrís elixen non migrar se son alimentados durante o inverno.[18] Ademais, os días máis longos do verán do norte proporcionan máis tempo ás aves reprodutoras para buscar comida para as crías. Isto axuda aos paxaros diúrnos a producir postas maiores que as que poñen especies de paxaros relacionados non migratorios que se quedan nos trópicos. A medida que se acurtan os días en outono, as aves volven ás rexións máis cálidas, onde a dispoñibilidade de comida varía pouco durante a estación.[19]
Estas vantaxes compensan o grande estrés, os custos do cansazo físico, e outros riscos que ten a migración. A predación pode intensificarse durante a migración: o falcón da raíña (Falco eleonorae), que se reproduce en illas do mar Mediterráneo, ten unha tempada reprodutora moi tardía, porque está coordinada co paso outonal cara ao sur de paseriformes migrantes, cos que alimentan as súas crías. Unha estratexia similar é a que adopta o morcego Nyctalus lasiopterus, que tamén depreda nos paseriformes migrantes.[20][21][22] As maiores concentracións de aves migrantes en sitios de parada fan que sexan máis susceptibles á infestación por parasitos e patóxenos, o que require un aumento da resposta inmune.[17]
Nunha especie non todas as poboacións poden ser migratorias; isto denomínase "migración parcial". A migración parcial é moi común nos continentes do sur; en Australia o 44% das aves non paseriformes e o 32% dos paseriformes son parcialmente migratorias.[23] Nalgunhas especies, a poboación en latitudes máis altas tende a ser migratoria e adoita invernar en latitudes máis baixas. As aves migrantes pasan de longo polas latitudes onde outras poboacións poden ser sedentarias, onde os hábitats axeitados para invernar probablemente xa están ocupados. Isto é un exemplo de migración a salto de ra.[24] Moitas especies completamente migratorias mostran este tipo de migración a salto de ra (aves que aniñan en latitudes máis altas pasan o inverno en latitudes máis baixas), e moitas mostran o tipo alternativo de migración, chamado migración en cadea, na que as poboacións 'deslízanse' máis gradualmente de norte a sur sen reverter a orde.[25]
Nunha poboación, é común que diferentes idades ou sexos presenten diferentes padróns de tempo e distancia na migración. As femias do pimpín Fringilla coelebs no leste de Fennoescandia migran antes no outono que os machos,[26] e os páridos europeos dos xéneros Parus e Cyanistes só migran no seu primeiro ano.[27]
A maioría das migracións empezan coas aves saíndo formando unha ampla fronte. A miúdo esta fronte estréitase formando unha ou máis rutas preferidas chamadas vías de voo. Estas rutas seguen tipicamente cadeas montañosas ou liñas de costa, ás veces ríos, e poden aproveitarse de correntes ascendentes ou padróns dos ventos ou evitar barreiras xeográficas como longas extensións de augas abertas. As rutas específicas poden ser programadas xeneticamente ou aprendidas en varios graos. As rutas que toman nas migracións de ida e de volta son con frecuencia diferentes.[17] Un padrón común en Norteamérica é unha migración no sentido das agullas do reloxo, na que as aves que voan cara ao norte tenden a pasar máis ao oeste, e as que voan cara ao sur tenden a viaxar desprazadas ao leste.
Moitas, se non a maioría, das aves migran en bandadas. Nas aves máis grandes, voar en bandadas reduce o custo de enerxía. Os gansos nunha formación en V poden conservar do 12 ao 20% da enerxía que necesitarían se voasen sós.[28][29] En estudos por radar encontrouse que os pilros Calidris canutus e Calidris alpina voaban 5 km/h máis rápidos en bandadas que voando sós.[17]

As aves voan durante a migración a varias altitudes. Unha expedición ao Monte Everest encontrou esqueletos de pato rabilongo (Anas acuta) e de mazarico rabinegro (Limosa limosa) a 5 000 m de altura no glaciar de Khumbu.[30] O ganso Anser indicus foi detectado por GPS voando a 6 540 m ao cruzar os Himalaias, e ademais ten a maior velocidade ascensional de calquera ave. Informes illados de voos desta ave aínda máis altos non foron confirmados debidamente.[31] As aves mariñas voan a baixa altura sobre a auga pero gañan altura cando cruzan a terra, e obsérvase o padrón inverso en aves terrestres.[32][33] Porén, a maioría das migracións das aves son de entre 150 e 600 m. Os rexistros de aviación sobre choques con aves nos Estados Unidos indican que a maioría das colisións ocorren por debaixo dos 600 m e case ningunha por riba dos 1 800 m.[34]
A migración das aves non está limitada a aves que poden voar. A maioría das especies de pingüíns (Spheniscidae) migran nadando. Estas rutas poden ser de 1 000 km. O tetraonino Dendragapus obscurus realiza unha migración altitudinal principalmente camiñando. Os emús (Dromaius novaehollandiae) de Australia emprenden movementos a gran distancia a pé durante os períodos de grandes secas.[17]
Comportamento migratorio nocturno
[editar | editar a fonte]Cando realizan migracións nocturnas moitas aves emiten 'chamadas de voo nocturno', que son chamadas curtas de contacto.[35] Estas chamadas probablemente serven para manter a composición dunha bandada migrante, e poden ás veces codificar o xénero dun individuo migrante.[36] Tamén serven probablemente para evitar o choque das aves no aire.[35] A migración nocturna pode monitorizarse usando datos de radar meteorolóxico,[37] que pode servir para estimar o número de aves que migran nunha determinada noite, así como a dirección da migración.[38] Entre as investigacións futuras previstas neste campo están a detección automática e a identificación das chamadas nocturnas das aves migrantes,[39] o cal podería ter grande importancia na conservación das especies e a xestión do territorio.
Os migrantes nocturnos toman terra pola mañá e poden alimentarse en terra durante uns poucos días antes de continuar coa migración. Estas aves son denominadas migrantes de paso nas rexións onde aparecen durante breve tempo entre a orixe e o destino.[40]
Os migrantes nocturnos minimizan a depredación, evitan o sobrequecemento, e poden alimentarse durante o día.[4] Un custo que ten viaxar pola noite é a perda do tempo para durmir. Os migrantes poden ás veces alterar a calidade do seu sono para compensar esta perda.[41]
Migración de longa distancia
[editar | editar a fonte]A imaxe típica dunha migración é a de aves terrestres do norte, como as andoriñas (Hirundinidae) e aves de rapina, que fan longos voos aos trópicos. Porén, moitas especies de anátidos e frinxílidos holárticos invernan na zona temperada do norte, en rexións con invernos máis suaves que os territorios de cría de verán. Por exemplo, o ganso picocurto migra desde Islandia a Gran Bretaña e países veciños, mentres que o xunco Junco hyemalis migra desde as zonas de clima subártico e ártico aos Estados Unidos[42] e o frinxílido Spinus tristis desde a taiga aos terreos de invernada que se estenden desde o sur dos Estados Unidos cara ao noroeste ata o oeste de Oregón.[43] Algúns patos, como a cerceta albela (Anas querquedula), móvense total ou parcialmente cara aos trópicos. O papamoscas negro (Ficedula hypoleuca) tamén segue esta tendencia migratoria, reproducíndose en Asia e Europa e invernando en África.
As rutas de migración e os territorios de invernada están ambos determinados xenetica e tradicionalmente dependendo do sistema social da especies. En especies sociais de vida longa como as cegoñas brancas (Ciconia ciconia), as bandadas son con frecuencia guiadas polos membros máis vellos e as cegoñas novas aprenden a ruta na súa primeira viaxe.[44] En especies de vida curta que migran soas, como a papuxa das amoras (Sylvia atricapilla) ou o cuco Coccyzus americanus, os migrantes do primeiro ano seguen unha ruta determinada xeneticamente que é alterable con apareamento selectivo.[45][46]
A miúdo, a ruta da migración dunha ave migratoria de longa distancia non segue unha liña recta entre os territorios de reprodución e os de invernada. Ao contrario, pode seguir unha dirección curva en arco ou gancho, con desvíos arredor das barreiras xeográficas ou cara a sitios de parada con hábitats axeitados. Para a maioría das aves terrestres, tales barreiras poderían consistir en mares, grandes corpos de auga ou altas cadeas montañosas, unha falta de sitios de parada para a alimentación, ou unha falta de columnas térmicas de aire (importante en aves de ás anchas).[14] Ademais, moitas rutas de migración son enrevesadas debido á historia evolutiva: a área reprodutora do pedreiro cincento (Oenanthe oenanthe) expandiuse para cubrir todo o hemisferio norte, pero a especie aínda migra ata 14 500 km para chegar aos seus ancestrais terreos de invernada na África subsahariana en vez de establecer novos territorios de invernada máis próximos ás súas áreas de cría.[47]
As mesmas consideracións sobre as barreiras e desvíos que se aplican á migración de aves terrestres de gran distancia tamén se aplican ás aves acuáticas, pero ao revés: unha grande área de terra sen corpos de auga que ofrezan sitios para a alimentación pode tamén ser unha barreira para unha ave que se alimente nas augas costeiras. Tamén se producen desvíos para evitar tales barreiras: por exemplo, o ganso de cara negra (Branta bernicla) que migra desde a península de Taymyr ao mar de Wadden viaxa pola costa do mar Branco e do Báltico en vez de facelo directamente a través do océano Glacial Ártico e o norte de Escandinavia.[48]
En limícolas
[editar | editar a fonte]
Unha situación similar ocorre coas limícolas. Moitas especies como o pilro común (Calidris alpina)[49] e o pilro de Maur (Calidris mauri),[50] emprenden longos desprazamentos desde os seus territorios de cría do Ártico a sitios máis cálidos no mesmo hemisferio, pero outros, como o pilro semipalmado (C. pusilla) viaxan a maiores distancias aos trópicos do hemisferio sur.[51]
Nalgunhas especies de limícolas, o éxito reprodutor depende da dispoñiilidade de certas fontes clave de aimentación en sitios onde fan parada ao longo da ruta de migración. Isto dálle aos migrantes a oportunidade de recargar enerxía para a seguinte etapa da viaxe. Algúns exemplos de importantes sitios de parada son a baía de Fundy e a de Delaware.[52][53]
Algúns mazaricos rubios (Limosa lapponica) realizan o voo máis longo sen escalas coñecido de calquera ave migrante, voando 11 000 km desde Alasca ás súas áreas de cría de Nova Zelandia.[54] Antes da migración, o 55% do seu peso corporal almacénase como graxa, que servirá de fonte de enerxía para a súa viaxe ininterrompida.
En aves mariñas
[editar | editar a fonte]
A migración das aves mariñas segue un padrón similar á dos limícolas e anátidos. Algúns, como o arao de á branca (Cepphus grylle) e algunhas gaivotas, son bastante sedentarios; outros, como a maioría dos carráns e álcidos, reprodúcense no hemisferio norte temperado, e móvense a diversas distancias cara ao sur durante o inverno setentrional. O carrán ártico (Sterna paradisaea) fai a migración de máis longa distancia de todas as aves, e ve máis luz do día ao longo do ano que calquera outra ave, xa que se move desde os territorios de cría árticos ás áreas non reprodutoras antárticas.[55] Un exemplar de carrán ártico que fora anelamento cando era un poliño nas illas Farne da costa leste británica, chegou a Melbourne, Australia en só tres meses despois de emplumar, facendo unha viaxe duns 22 000 km. Moitas aves con tubos nasais (procelariformes) reprodúcense no hemisferio sur e migran ao norte durante o inverno do sur.[56]
A maioría das especies peláxicas, principalmente as da orde Procellariiformes, son grandes aves errantes, e os albatros dos océanos do sur poden circundar o globo voando na zona dos ventos "corenta ruxentes" (da latitude 40º S) fóra da súa tempada reprodutora. Os procelariformes espállanse moito en enormes áreas do océano aberto, pero congréganse onde hai abundancia de comida. Moitos están tamén entre os migradores de maiores distancias; as pardelas escuras (Puffinus griseus) que aniñan nas illas Malvinas migran 14 000 km entre as colonias reprodutoras e o Atlántico norte á altura de Noruega. Algúns furabochos atlánticos (Puffinus puffinus) fan a mesma viaxe á inversa. Como son aves de vida longa, poden cubrir enormes distancias durante as súas vidas; un furabochos atlántico plusmarquista calcúlase que voou 8 millóns de km durante os seus 50 anos de vida.[57]
Migración diúrna en aves de rapina
[editar | editar a fonte]
Algunhas aves de ás anchas grandes dependen das columnas térmicas de aire quente ascendente que lles permiten remontarse e planar. Entre estas están moitas aves de rapina como os voitres, aguias e miñatos, pero tamén outras como as cegoñas. Estas aves migran durante as horas diúrnas. As especies migratorias destes grupos teñen grandes dificultades para cruzar grandes corpos de auga, xa que as térmicas só e forman en terra, e estas aves non poden manter un voo activo durante longas distancias. O mar Mediterráneo e outros mares supoñen un obstáculo enorme para as aves que voan remontándose e planando, que deben cruzar, como moito, só polos puntos máis estreitos destes mares. Cantidades masivas de grandes aves de rapina e cegoñas pasan por áreas como o estreito de Messina,[58] Xibraltar, Falsterbo, e o Bósforo na época da migración. Especies máis comúns como o miñato abelleiro (Pernis apivorus), poden contarse por centos ou miles no outono neses lugares. Outras barreiras, como as cadeas montañosas, poden tamén canalizar as rutas de migración, especialmente no caso dos grandes migrantes diúrnos. Este é un importante factor no pescozo de botella migratorio de América Central. O pescozo de botella de Batumi no Cáucaso é un dos funís migratorios máis conxestionados da Terra. Para evitaren voar sobre o mar Negro e atravesar montañas, centos ou miles de aves remontadoras e planadoras canalizanse pola área da cidade de Batumi, Xeorxia.[59] As aves de presa como os miñatos abelleiros que migran usando as térmicas perden só do 10 ao 20% do seu peso durante a migración, o que pode explicar por que se alimentan menos durante a migración que as aves de presa máis pequenas cun voo máis activo, como os falcóns, azores e gabiáns.[60]

Observando a migración de once especies de aves remontadoras e planadoras polo estreito de Xibraltar, en relación ao cambio climático e outros factores, as especies que non adiantaban as súas datas de migración outonais eran aquelas con poboacións reprodutoras en declive en Europa.[61]

Migración a curta distancia e altiudinal
[editar | editar a fonte]
Moitos migrantes de longa distancia parecen estar xeneticamente programados para responder aos cambios na duración da luz diúrna. Porén, as especies que se moven a curtas distancias, parece que non necesitan ese mecanismo para elixir o momento, senón que se moven en resposta ás condicións meteorolóxicas locais. Así os que se reproducen en montañas e páramos, como o gabeador vermello (Tichodroma muraria) e o merlo rieiro (Cinclus cinclus), poden moverse só altitudinalmente para escapar dos terreos máis altos fríos. Outras especies, como o esmerillón (Falco columbarius) e a laverca (Alauda arvensis) móvense a maiores distancias, ata a costa ou cara ao sur. Especies como o pimpín son moito menos migratorios en Gran Bretaña que en Europa continental, e a maioría non se moven mís de 5 km durante a súa vida.[62]
Os paseriformes migrantes de curta distancia teñen dúas orixes evolutivas. Aqueles que teñen migrantes de longa distancia na súa familia, como o picafollas europeo (Phylloscopus collybita), son especies orixinadas no hemisferio sur que progresivamente acurtaron a súa migración de retorno permanecendo no hemisferio norte.[63]
As especies que non teñen parentes migradores de longa distancia, como os picoteiros (Bombycilla), móvense en resposta ao tempo invernal e á perda do seu alimento usual invernal, en vez de para mellorar as súas oportunidades de reprodución.[64]
Nos trópicos hai pouca variación na lonxitude do día ao longo do ano, e alí sempre fai calor dabondo como para ter dispoñibilidade de comida, pero a migración altitudinal ocorre tamén nalgunhas aves tropicais. Hai probas de que isto permite que os migrantes obteñan máis cantidade da súa comida preferida, como a froita.[65]
A migración altitudinal é común nas montañas de todo o mundo, como os Himalaias e os Andes.[66]
Moitas especies de aves de rexións áridas do sur de Australia son nómades; seguen as fontes de alimento e de auga ao longo do país nun padrón irregular, o cal non está relacionado coas estacións senón coas chuvias. Poden pasar varios anos entre as visitas a unha área feitas por unha determinada especie.[67]
Irrupcións e dispersión
[editar | editar a fonte]
Algunhas veces danse circunstancias como unha boa tempada de reprodución seguida dunha escaseza de fontes de alimentación no seguinte ano que levan a irrupcións nas que numerosas especies se moven alén da súa área normal. Un exemplo disto son as invasións de picoteiros e cruzabicos rexistradas en Inglaterra nos últimos douscentos anos. Os picoteiros (Bombycilla garrulus) mostran ben estas variacións impredicibles nas súas cantidades anuais, que deron lugar a cinco grandes chegadas de aves a Gran Bretaña durante o século XIX, e 18 entre os anos 1937 e 2000.[64] Os cruzabicos (Loxia curvirostra) tamén fan estas irrupcións, con amplas invasións en Inglaterra en 1251, 1593, 1757 e 1791.[68]
A migración das aves é principalmente, pero non exclusivamente, un fenómeno do hemisferio norte.[69] Isto débese a que nas latitudes altas do norte, onde o alimento se fai escaso en inverno, as aves parten a áreas máis ao sur (incluso chegan ao hemisferio sur) para invernar, e porque as masas continentais son moito maiores no hemisferio norte. Ao contrario, para as aves mariñas (peláxicas) do hemisferio sur é máis doado migrar, porque hai unha ampla extensión oceánica, e máis illas axeitadas para que as aves mariñas aniñen.[70]
Fisioloxía e control
[editar | editar a fonte]O control da migración, o seu momento e resposta están xeneticamente controladas e parecen ser un carácter primitivo que está presente mesmo en especies de aves non migratorias. A capacidade de navegar e orientarse que teñen durante a migración é un fenómeno moito máis complexo que pode estar determinado por programas endóxenos e tamén a aprendizaxe.[71]
Momento da migración
[editar | editar a fonte]O primeiro sinal fisiolóxico para estimular a migración son os cambios na lonxitude do día. Estes cambios tamén están relaciondos con cambios hormonais nas aves. No período anterior á migración, moitas aves mostran unha maior actividade ou Zugunruhe (termo alemán que significa 'inquedanza migratoria'), descrita primeiramente por Johann Friedrich Naumann en 1795, e cambios fisiolóxicos como un incremento no depósito de graxas. A aparición do Zugunruhe incluso en aves criadas en gaiolas sen sinais ambientais (como o acurtamnto do día ou a baixada das temperaturas) indica o papel que xogan os programas endóxenos circanuais no control da migración das aves.[72] As aves en gaiolas mostan unha dirección de voo preferente que corresponde coa dirección migratoria que tomarían na natureza, e cambin a súa dirección preferente aproximadamente aomesmo tempo que os seus conxéneres silvestres libres.[73]
En especies polixinias con considerable dimorfismo sexual, os machos tenden a regresar antes dos sitios de cría que as femias. Isto denomínase protandria.[74][75]
Orientación e navegación
[editar | editar a fonte]
A navegación animal está baseada en diversos sentidos. Moitas aves usan un compás solar. Usar o sol para determinar a dirección implica ter que facer unha compensación baseada no tempo. A navegación noutros casos está baseada nunha combinación doutras habilidades como a de detectar os campos magnéticos (magnetocepción), uso de puntos de referencia visuais xeográficos e pistas olfactivas.[76]
Pénsase que os migrantes de longa distancia se dispersan cando son novos e quedan atraídos a sitios potenciais de reprodución e a sitios favoritos de invernada. Unha vez que se crea esta atracción ou fixación ao sitio mostran unha gran fidelidade a dito sitio, visitando os mesmos lugares de invernada ano tras ano.[77]
A capacidade das aves de navegar durante as migracións non pode explicarse totalmente pola existencia dun programa endóxeno, incluso coa axuda de respostas a sinais ambientais. A capacidade de realizar con éxito migracións de longa distancia probablemente só se pode explicar completamente contando coa capacidade cognitiva das aves para recoñecer os hábitats e formar mapas mentais. O seguimento por satélite das aves de presa que migran polo día, como as aguias pescadoras e miñatos abelleiros mostrou que os individuos máis vellos son mellores facendo correccións á deriva dos ventos que os desvía.[78] As aves navegan por un sentido biolóxico innato resultado da súa evolución. As aves migratorias poden usar dúas ferramentas electromagnéticas para encontrar os seus destinos: unha que é enteiramente innata e outra que depende da experiencia. Unha ave nova na súa primeira migración voa na correcta dirección de acordo co campo magnético terrestre, pero non sabe o longa que será a viaxe. Faino por medio dun mecanismo de par radical no que reaccións químicas, en especial fotopigmentos sensibles a lonxitudes de onda curtas de luz, son afectadas polo campo magnético. Aínda que isto só funciona durante as horas diúrnas, non usa a posición do sol de ningunha maneira. Neste estadio a ave está na mesma posición dun Boy Scout cun compás pero sen mapa, ata que se vai acostumando á viaxe e pode poñer a funcionar as súas outras capacidades. Con experiencia aprende varios puntos senlleiros xeográficos que lle serven de referencia e este "mapado" faise grazas aos cristais de magnetita que teñen no seu sistema trixémino, que lle indican á ave a forza que ten o campo magnético. Como as aves migran entre as rexións norte e sur da Terra, a distinta forza que ten o campo magnético en diferentes latitudes permítelles interpretar o mecanismo do par radical de forma máis precisa e poden saber cando chegaron ao seu destino.[79] Hai unha conexión neural entre o ollo e o chamado "Clúster N", a parte do prosencéfalo que é activa durante a orientación migracional, o que suxire que as aves poden realmente ver o campo magnético da Terra.[80][81]
Aves errantes
[editar | editar a fonte]As aves migrantes poden perder o seu rumbo e acabar fóra das súas áreas normais. Isto pode deberse a que voan pasando de largo o seu lugar de destino como ocorre no chamado "sobrepasamento primaveral", no cal aves que regresan das súas áreas de cría superan o punto de destino e acaban máis o norte do que pretendían. Certas áreas, debido á súa localización, adquiriron sona como puntos de observación de tales aves. Exemplos son o Parque Nacional de Point Pelee do Canadá, e Spurn en Inglaterra.
A migración inversa, na que a programación xenética das aves novas non funciona axeitadamente, e van en sentido diferente do normal, pode facer que aparezan aves errantes raras miles de quilómetros fóra da súa área normal.[82]
A deriva migratoria de aves desviadas polo vento pode ter como resultado a chegada de gran número de migrantes a sitios costeiros.[83]
Un fenómeno relacionado chamado "abmigración" supón que aves dunha rexión se unen a aves similares doutra diferente rexión de cría nos territorios de invernada comúns e despois migrn de volta xunto coa nova poboación. Isto é especialmente común nalgúns anátidos, que cambian dunha ruta a outra.[84]
Aprendizaxe por condicionamento da migración
[editar | editar a fonte]Foi posible ensinar unha ruta de migración a unha bandada de aves, por exemplo en programas de reintrodución de especies. Despois dun ensaio con gansos do Canadá (Branta canadensis), utilizouse un avión ultralixeiro nos EUA para ensinar rutas de migración seguras aos grous Grus americana reintroducidos .[85][86]
Adaptacións
[editar | editar a fonte]As aves necesitan alterar o seu metabolismo para afrontar as demandas da migración. O almacenamento de enerxía por medio da acumulación de graxa e o control do tempo de sono nos migrantes nocturnos precisa adaptacións fisiolóxicas especiais. Ademais, as plumas das aves sofren polo desgaste e desgarre e deben ser mudadas. O momento desta muda, xeralmente unha vez ao ano pero ás veces dúas, varía nalgunhas especies que mudan antes de desprazarse aos territorios de invernada e outras mudan antes de volver aos seus territorios de cría.[87][88] Á parte das adaptacións fisiolóxicas, a migración ás veces require cambios de comportamento como voar en bandadas para reducir a enerxía usada na migración ou diminuír o risco de depredación.[89]
Factores evolutivos e ecolóxicos
[editar | editar a fonte]A migración das aves é moi feble e crese que se desenvolveu independentemente en moitas liñaxes das aves.[90] Aínda que hai acordo en que as adaptacións de comportamento e fisiolóxicas necesarias para a migración están baixo control xenético, algúns autores argumentaron que non cómpre ningún cambio xenético para que se desenvolva un comportamento migratorio nunha especie sedentaria porque o armazón xenético para o comportamento migratorio existe en case todas as liñaxes de aves.[91] Isto explica a rápida aparición do comportamento migratorio despois do máximo glacial máis recente.[92]
As análises teóricas mostran que desvíos que incrementan a distancia de voo nun 20% son a miúdo adaptativos desde o punto de vista aerodinámico, xa que unha ave que se carga con moito alimento para cruzar unha longa barreira que atopa no seu camiño voa menos eficientemente. Porén algunhas especies mostran unhas complicadas rutas migratorias que reflicten as expansións históricas da súa área de disgtribución e están lonxe de ser rutas óptimas en termos ecolóxicos. Un exemplo é a migración de poboacións continentais do paxaro túrdido Catharus ustulatus, que voa moi ao leste atravesando Norteamérica antes de xirar ao sur a través de Florida para chegar ao norte de Suramérica; esta ruta crese que é a consecuencia dunha ampliación do seu rango de distribución que ocorreu hai uns 10 000 de anos. Os desvíos poden tamén ser causadas por condicións do vento diferenciais, risco de predación ou outros factores.[93]
Cambio climático
[editar | editar a fonte]Os cambios climáticos a grande escala, como os que se experimentaron no pasado da Terra, espérase que ten un efecto sobre a elección do momento da migración. Os estudos mostraron unha variedade de efectos, incluíndo o cambio no momento da migración, reprodución[94] e as variacións na poboación.[95][96]
Efectos ecolóxicos
[editar | editar a fonte]A migración das aves tamén axuda ao movemento doutras especies, incluíndo os ectoparasitos como carrachas e piollos,[97] que á súa vez poden transportar microorganismos incluíndo algúns que poden afectar a saúde humana. Debido ao espallamento da gripe aviaria, a migración das aves foi estudada como posible mecanismo de transmisión de doenzas, mais atopouse que non supoñía un risco especial; ao contrario, a importación de mascotas e aves domésticas é unha grande ameaza.[98] Algúns virus que se manteñen no corpo das aves sen efectos letais para elas, como o virus do Nilo Occidental poden ser espallados polas aves migratorias.[99] As aves poden tamén ter un papel na dispersión de propágulos de plantas e plancto.[100][101]
Algúns predadores aprovéitanse da concentración de aves que se produce durante a migración. O morcego Nyctalus lasiopterus aliméntase de paseriformes migrantes nocturnos.[21] Algunhas aves de presa especialízanse en capturar limícolas migrantes.[102]
Técnicas de estudo
[editar | editar a fonte]
Os primeiros estudos sobre o momento da migración empezaron en 1749 en Finlandia, onde Johannes Leche de Turku recompilou datos sobre as chegadas de aves migrantes en primavera.[103]
As rutas de migración das aves foron estudadas por medio de diversas técnicas, a máis antiga das cales foi a marcaxe das aves. Os cisnes foron marcados cunha amosega no peteiro desde o ano 1560 en Inglaterra. O anelamento de aves científico foi iniciado por Hans Christian Cornelius Mortensen en 1899.[104] Outras técnicas son o uso do radar[105] e o seguimento por satélite.[106] Observouse que a taxa de migración das aves sobre os Alpes (ata unha altura de 150 m) era moi comparable entre os datos medidos por radar de feixe fixo e os recontos feitos visualmente, destacando o uso potencial desta técnica como maneira obxectiva de cuantificar a migración das aves.[107]
Os isótopos estables do hidróxeno, oxíxeno, carbono, nitróxeno e xofre poden establecer a conexión migratoria das aves entre os sitios de invernada e os terriotorios de reprodución. Estes métodos dependen das diferenzas isotópicas espaciais na composición da dieta das aves que son incorporados nos tecidos inertes como as plumas, ou en tecidos en crecemento como garras e músculo ou sangue.[108][109]
Unha estratexia para identificar a intensidade da migración utiliza micrófonos que apuntan cara a arriba para rexistrar as chamadas de contacto nocturnas das bandadas que pasan voando por encima. Estas son despois analizadas nun laboratorio para medir o tempo, frecuencia e especies.[110]

Unha técnica máis vella desenvolvida por George Lowery e outros para cuantificar a migración consiste en observar a lúa chea cun telescopio e contar as siluetas das bandadas de aves que voan diante da lúa pola noite.[111][112]
Os estudos de comportamento de orientación foron realizados tradicionalmente usando variantes dun dispositivo chamado funil de Emlen, que consiste nunha gaiola circular coa parte superior cuberta con cristal ou pantalla de arame para que sexa visible o ceo ou para que o dispositivo se coloque nun planetario ou con outros controis sobre os sinais ambientais. O comportamento de orientación das aves dentro da gaiola estúdase cuantitativaemnte usando as marcas de distribución que as aves deixan nas paredes da gaiola.[113] Outras estratexias utilizadas nos estudos de pombas que regresan a casa usan a dirección na cal as aves desaparecen polo horizonte.[114]
Ameazas e conservación
[editar | editar a fonte]
As actividades humanas ameazan moitas aves migratorias. As distancias implicadas na migración das aves fan que a miúdo crucen as fronteiras políticas de países, polo que as medidas de conservación requiren a cooperación internacional. Asináronse varios tratados internacionais para protexer as especies migratorias, como a Lei do Tratado de Aves Migratorias de 1918 dos Estados Unidos, que aplicaba a convención para a protección de aves migratorias entre Estados Unidos e o Canadá,[115] e o Acordo para a conservación das aves acuáticas migratorias africano-euroasiáticas[116]
A concentración de aves durante a migración pode poñer en risco unha especie. Algúns migrantes espectaculares xa se extinguiron; durante a migración da hoxe extinguida pomba viaxeira (Ectopistes migratorius) en Norteamérica xuntábanse enormes bandadas de 1,6 km de ancho e de 480 km de longo, que escurecían o ceo e tardaban varios días en pasar.[117] A especie foi cazada masivamente para comercializar a súa carne no século XIX e tamén foi afectada pola deforestación.
Outras áreas significativas son os sitios de parada entre os territorios de cría e de invernada.[118] Un estudo de captura e recaptura de paseriformes migrantes con gran fidelidade para os sitios de cría e invernada non mostrou unha asociación estrita cos sitios de parada.[119]
A caza ao longo das rutas de migración ameaza algunhas especies de aves. As poboacións de grúa siberiana (Leucogeranus leucogeranus) que invernaban na India diminuíron debido á caza ao longo da ruta, especialmente en Afganistán e Asia central. As aves foron vistas por última vez nos seus territorios favoritos de invernada do Parque Nacional de Keoladeo en 2002.[120] Estruturas como as liñas de alta tensión, muíños de enerxía eólica e plataformas petrolíferas no mar tamén afectan as aves migratorias.[121] Outros perigos son a contaminación, tormentas, incendios forestais e destrución do hábitat ao longo das rutas migratrorias, que impiden que os migrantes atopen comida nos sitios de parada.[122] Por exemplo, na vía migratoria desde o leste de Asia a Australasia, ata o 65% dos hábitats intermareais claves no pescozo de botella da migración no mar Amarelo foron destruídos desde a década de 1950.[123][124]
Notas
[editar | editar a fonte]- ↑ "Essential Migration: A Study of Surjeet Kalsey’s "Migratory Birds"". www.academia.edu. Consultado o 2016-04-04.
- ↑ A Biblia. 2ª edición. SEPT. 1992. Vigo. Libro de Xob, capítulo 39, versículo 26. Páxina 633. ISBN 84-7337-040-6.
- ↑ A Biblia. 2ª edición. SEPT. 1992. Vigo. Libro de Xeremías, capítulo 8, versículo 7. Páxina 1067. ISBN 84-7337-040-6.
- ↑ 4,0 4,1 4,2 Lincoln, F. C. (1979). Migration of Birds. Circular 16 (Fish and Wildlife Service).
- ↑ Cocker, Mark; Mabey, Richard (2005). Birds Britannica. Chatto & Windus. p. 315. ISBN 0-7011-6907-9.
- ↑ 6,0 6,1 White, 1898. pp. 27–28
- ↑ White, 1898. pp. 161–162
- ↑ Bewick, 1797. p. xvii
- ↑ Bewick, 1797. p. 300
- ↑ Bewick, 1797. pp. 302–303
- ↑ Zoologische Sammlung der Universität Rostock (en alemán) artigo cunha imaxe dunha Rostocker Pfeilstorch
- ↑ Flyer for the Rostock University Zoological Collection Arquivado 22 de marzo de 2012 en Wayback Machine. (en inglés)
- ↑ Der Sproessling 3 Arquivado 25 de novembro de 2014 en Wayback Machine. (en alemán) edición da revista da asociaciónn de estudantes local que contén un artigo sobre a Pfeilstorch
- ↑ 14,0 14,1 Peter Berthold; Hans-Günther Bauer; Valerie Westhead (2001). Bird Migration: A General Survey. Oxford: Oxford University Press. ISBN 0-19-850787-9.
- ↑ Sekercioglu, C.H. (2007). "Conservation ecology: area trumps mobility in fragment bird extinctions". Current Biology 17 (8): 283–286. doi:10.1016/j.cub.2007.04.045.
- ↑ Rolland J, Jiguet F, Jønsson KA, Condamine FL & Morlon H (2014). "Settling down of seasonal migrants promotes bird diversification". Proceedings of the Royal Society B 281 (1784): 20140473. PMC 4043101. PMID 24759866. doi:10.1098/rspb.2014.0473.
- ↑ 17,0 17,1 17,2 17,3 17,4 Newton, I. (2008). The Migration Ecology of Birds. Elsevier. ISBN 978-0-12-517367-4.
- ↑ "Migration Basics". Hummingbirds.net. Consultado o 10 de abril de 2014.
- ↑ Ramachandra, T.V.; et al. (febreiro de 2011). "Environmental Impact Assessment of the National Large Solar Telescope Project and its Ecological Impact in Merak Area". p. 71. Consultado o 10 de abril de 2014.
- ↑ Dondini, G.; Vergari, S. (2000). "Carnivory in the greater noctule bat (Nyctalus lasiopterus) in Italy". Journal of Zoology 251 (2): 233–236. doi:10.1111/j.1469-7998.2000.tb00606.x.
- ↑ 21,0 21,1 Popa-Lisseanu, A. G., Delgado-Huertas, A., Forero, M. G., Rodriguez, A., Arlettaz, R. & Ibanez, C. (2007). Rands, Sean, ed. "Bats' Conquest of a Formidable Foraging Niche: The Myriads of Nocturnally Migrating Songbirds". PLOS ONE 2 (2): e205. PMC 1784064. PMID 17299585. doi:10.1371/journal.pone.0000205.
- ↑ Ibáñez, C.; Juste, J.; García-Mudarra, J. L.; Agirre-Mendi, P. T. (2001). "Bat predation on nocturnally migrating birds". PNAS 98 (17): 9700–9702. PMC 55515. PMID 11493689. doi:10.1073/pnas.171140598. Arquivado dende o orixinal o 21 de agosto de 2008. Consultado o 21 de agosto de 2018.
- ↑ Chan K (2001). "Partial migration in Australian landbirds: a review". Emu 101 (4): 281–292. doi:10.1071/MU00034.
- ↑ Boland, J. M. (1990). "Leapfrog migration in North American shorebirds: intra- and interspecific examples" (PDF). The Condor 92 (2): 284–290. JSTOR 1368226. doi:10.2307/1368226.
- ↑ Berthold, Peter (2001). Bird Migration: A General Survey. Oxford University Press. p. 67.
- ↑ Panov, Ilya N. (2011). "Overlap between moult and autumn migration in passerines in northern taiga zone of Eastern Fennoscandia" (PDF). Avian Ecology and Behaviour 19: 33–64.
- ↑ Ketterson, E. D., and V. Nolan. 1985. Intraspecific variation in avian migration: evolutionary and regulatory aspects, Pages 553-579 in M. A. Rankin, ed. Migration: mechanisms and adaptive significance, University of Texas, Austin.
- ↑ Hummel D.; Beukenberg M. (1989). "Aerodynamische Interferenzeffekte beim Formationsfl ug von Vogeln". J. Ornithol 130: 15–24. doi:10.1007/BF01647158.
- ↑ Cutts, C. J. & J R Speakman (1994). "Energy savings in formation flight of Pink-footed Geese" (PDF). J. Exp. Biol. 189 (1): 251–261. PMID 9317742.
- ↑ Geroudet, P. (1954). "Des oiseaux migrateurs trouvés sur la glacier de Khumbu dans l'Himalaya". Nos Oiseaux 22: 254.
- ↑ Swan, L. W. (1970). "Goose of the Himalayas". Nat. Hist. 79 (10): 68–75.
- ↑ Dorst, J. (1963). The migration of birds. Houghton Mifflin Co., Boston. p. 476.
- ↑ Eastwood, E. & G. C. Rider. (1965). "Some radar measurements of the altitude of bird flight". British Birds 58: 393–426.
- ↑ Williams, G. G. (1950). "Weather and spring migration". Auk 67: 52–65. doi:10.2307/4080769.
- ↑ 35,0 35,1 Farnsworth, Andrew (2005-07-01). "Flight calls and their value for future ornithological studies and conservation research". The Auk 122 (3): 733–746. ISSN 0004-8038. doi:10.1642/0004-8038(2005)122[0733:FCATVF]2.0.CO;2.
- ↑ Griffiths, Emily T.; Keen, Sara C.; Lanzone, Michael; Farnsworth, Andrew (2016-06-10). "Can Nocturnal Flight Calls of the Migrating Songbird, American Redstart, Encode Sexual Dimorphism and Individual Identity?". PLOS ONE 11 (6): e0156578. ISSN 1932-6203. doi:10.1371/journal.pone.0156578.
- ↑ Farnsworth, Andrew; Van Doren, Benjamin M.; Hochachka, Wesley M.; Sheldon, Daniel; Winner, Kevin; Irvine, Jed; Geevarghese, Jeffrey; Kelling, Steve (2016-04-01). "A characterization of autumn nocturnal migration detected by weather surveillance radars in the northeastern USA". Ecological Applications (en inglés) 26 (3): 752–770. ISSN 1939-5582. doi:10.1890/15-0023.
- ↑ Dokter, Adriaan M.; Liechti, Felix; Stark, Herbert; Delobbe, Laurent; Tabary, Pierre; Holleman, Iwan (2011-01-06). "Bird migration flight altitudes studied by a network of operational weather radars". Journal of The Royal Society Interface (en inglés) 8 (54): 30–43. ISSN 1742-5689. PMC 3024816. PMID 20519212. doi:10.1098/rsif.2010.0116.
- ↑ Salamon, Justin; Bello, Juan Pablo; Farnsworth, Andrew; Robbins, Matt; Keen, Sara; Klinck, Holger; Kelling, Steve (2016-11-23). "Towards the Automatic Classification of Avian Flight Calls for Bioacoustic Monitoring". PLOS ONE 11 (11): e0166866. ISSN 1932-6203. doi:10.1371/journal.pone.0166866.
- ↑ Schmaljohann, Heiko, Felix Liechti and Bruno Bruderer (2007). "Songbird migration across the Sahara: the non-stop hypothesis rejected!". Proceedings of the Royal Society B 274 (1610): 735–739. PMC 2197203. PMID 17254999. doi:10.1098/rspb.2006.0011.
- ↑ Rattenborg, N.C.; Mandt, B.H.; Obermeyer, W.H.; Winsauer, P.J.; Huber, R. (2004). "Migratory Sleeplessness in the White-Crowned Sparrow (Zonotrichia leucophrys gambelii)". PLoS Biol. 2 (7): e212. PMC 449897. PMID 15252455. doi:10.1371/journal.pbio.0020212.
- ↑ Dark-Eyed Junco
- ↑ American Goldfinch
- ↑ Chernetsov N., Berthold P., Querner U. (2004). "Migratory orientation of first-year white storks (Ciconia ciconia): inherited information and social interactions". Journal of Experimental Biology 207: 937–943. doi:10.1242/jeb.00853.
- ↑ Sutherland W. J. (1998). "Evidence for flexibility and constraint in migration systems". Journal of Avian Biology 29: 441–446. doi:10.2307/3677163.
- ↑ Berthold P., Helbig A. J., Mohr G., Querner U. (1992). "Rapid microevolution of migratory behaviour in a wild bird species". Nature 360: 668–670. doi:10.1038/360668a0.
- ↑ Bairlein F., Norris D. R., Nagel R., Bulte M., Voigt C. C., Fox J. W., Hussell D. J. T.; et al. (2012). "Cross-hemisphere migration of a 25 g songbird". Biology Letters 8: 505–507. PMC 3391447. doi:10.1098/rsbl.2011.1223.
- ↑ Green, Martin (1999). "The Riddle of the White Sea". Geese.org. Arquivado dende o orixinal o 29 de xullo de 2020. Consultado o 10 de abril de 2014.
- ↑ "Species factsheet: Dunlin Calidris alpina". BirdLife International. 2014. Arquivado dende o orixinal o 12 de agosto de 2014. Consultado o 19 de xuño de 2014.
- ↑ "Species factsheet: Western Sandpiper Calidris mauri". BirdLife International. 2014. Arquivado dende o orixinal o 12 de agosto de 2014. Consultado o 19 de xuño de 2014.
- ↑ "Species factsheet: Semipalmated Sandpiper Calidris pusilla". BirdLife International. 2014. Arquivado dende o orixinal o 12 de agosto de 2014. Consultado o 19 de xuño de 2014.
- ↑ Sprague, A. J.; D. J. Hamilton & A. W. Diamond (2008). "Site Safety and Food Affect Movements of Semipalmated Sandpipers (Calidris pusilla) Migrating Through the Upper Bay of Fundy" (PDF). Avian Conservation and Ecology 3 (2).
- ↑ Kathleen E. Clark, Lawrence J. Niles and Joanna Burger (1993). "Abundance and Distribution of Migrant Shorebirds in Delaware Bay" (PDF). The Condor 95 (3): 694–705. JSTOR 1369612. doi:10.2307/1369612.
- ↑ Gill, Robert E. Jr.; Theunis Piersma; Gary Hufford; Rene Servranckx; Adrian Riegen (2005). "Crossing the ultimate ecological barrier: evidence for an 11,000 km-long nonstop flight from Alaska to New Zealand and Eastern Australia by Bar-tailed Godwits". The Condor 107 (1): 1–20. doi:10.1650/7613.
- ↑ Cramp, S., ed. (1985). Birds of the Western Palearctic. pp. 87–100. ISBN 0-19-857507-6.
- ↑ Pyle, Peter (2001). "Seabirds" (PDF). Circular 1198. USGS. p. 154. Consultado o 19 de xuño de 2014.
- ↑ Anon (18 de abril de 2002). "Oldest bird clocks 5 million miles". CNN.com. Consultado o 31 de marzo de 2013.
- ↑ Corso, Andrea. "European Birding Hot Spot: The Strait of Messina, southern Italy".[Ligazón morta]
- ↑ Maanen, E. van; Goradze, I.; Gavashelishvili, A.; Goradze, R. (2001). "Opinion: Trapping and hunting of migratory raptors in western Georgia". Bird Conservation International 11 (2): 77–92. doi:10.1017/S095927090100017X.
- ↑ Gensbol, B; (1984) Collins Guide to the Birds of Prey of Britain and Europe, p.28
- ↑ Panuccio, M.; Martín, B.; Onrubia, A.; Ferrer, M. (2017). "Long-term changes in autumn migration dates at the Strait of Gibraltar reflect population trends of soaring birds". Ibis 159 (1): 55–65. doi:10.1111/ibi.12420.
- ↑ "British Wildlife Recordings: Chaffinch". British Library. Arquivado dende o orixinal o 13 de abril de 2014. Consultado o 10 de abril de 2014.
- ↑ Cocker, 2005. p. 378
- ↑ 64,0 64,1 Cocker, 2005. p. 326
- ↑ Boyle, W. A.; Conway, C. J.; Bronstein, J. L. (2011). "Why do some, but not all, tropical birds migrate? A comparative study of diet breadth and fruit preference" (PDF). Evolutionary Ecology 25: 219–236. doi:10.1007/s10682-010-9403-4.
- ↑ Kreft, Stefan (23 de xuño de 2004). "The Fourth Dimension: An Overview of Altitudinal Migration" (PDF). 25th Annual Bonn Convention, Berlin. Consultado o 27 de marzo de 2013.
- ↑ Rohan., Clarke, (2014). Finding Australian Birds : a Field Guide to Birding Locations. CSIRO Publishing. p. xiv. ISBN 978-1-4863-0084-6. OCLC 880410149.
- ↑ Cocker, 2005. p. 455
- ↑ Somveille M, Manica A, Butchart SH, Rodrigues AS (2013). "Mapping Global Diversity Patterns for Migratory Birds". PLoS ONE 8 (8): e70907. PMC 3737225. PMID 23951037. doi:10.1371/journal.pone.0070907.
- ↑ Newton, Ian (2010). "13. Large-Scale Movement Patterns". The Migration Ecology of Birds. Academic Press. pp. 396, and throughout. ISBN 978-0-08-055483-9.
- ↑ Helm B, Gwinner E (2006). "Migratory Restlessness in an Equatorial Nonmigratory Bird". PLoS Biol. 4 (4): e110. PMC 1420642. PMID 16555925. doi:10.1371/journal.pbio.0040110.
- ↑ Fusani, L.; Cardinale, L.; Carere, C.; Goymann, W. (2009). "Stopover decision during migration: physiological conditions predict nocturnal restlessness in wild passerines". Biology Letters 5 (3): 302–305. PMC 2679912. PMID 19324648. doi:10.1098/rsbl.2008.0755.
- ↑ Nievergelt, F.; Liechti, F.; Bruderer, B. (1999). "MIGRATORY DIRECTIONS OF FREE-FLYING BIRDS VERSUS ORIENTATION IN REGISTRATION CAGES" (PDF). Journal of Experimental Biology 202 (16): 2225–2231.
- ↑ Diego Rubolini; Fernando Spina & Nicola Saino (2004). "Protandry and sexual dimorphism in trans-Saharan migratory birds". Behavioral Ecology 15 (4): 592–601. doi:10.1093/beheco/arh048.
- ↑ Edwards, Darryl B.; Forbes, Mark R. (2007). "Absence of protandry in the spring migration of a population of Song Sparrows Melospiza melodia". Ibis 149 (4): 715–720. doi:10.1111/j.1474-919X.2007.00692.x.
- ↑ Walraff, H. G. (2005). Avian Navigation: Pigeon Homing as a Paradigm. Springer.
- ↑ Ketterson, E.D. & V. Nolan Jr. (1990). "Site attachment and site fidelity in migratory birds: experimental evidence from the field and analogies from neurobiology.". En E. Gwinner. Bird Migration (PDF). Springer Verlag. pp. 117–129. Arquivado dende o orixinal (PDF) o 10 de febreiro de 2010. Consultado o 21 de agosto de 2018.
- ↑ Thorup, Kasper; Thomas Alerstam; Mikael Hake; Nils Kjelle (2003). "Bird orientation: compensation for wind drift in migrating raptors is age dependent". Proceedings of the Royal Society B 270 (Suppl 1): S8–S11. PMC 1698035. PMID 12952622. doi:10.1098/rsbl.2003.0014.
- ↑ Wiltschko, W.; U. Munro; H. Ford; R. Wiltschko (2006). "Bird navigation: what type of information does the magnetite-based receptor provide?". Proceedings of the Royal Society B 273 (1603): 2815–20. PMC 1664630. PMID 17015316. doi:10.1098/rspb.2006.3651.
- ↑ Heyers D, Manns M, Luksch H, Güntürkün O, Mouritsen H (2007). Iwaniuk A, ed. "A Visual Pathway Links Brain Structures Active during Magnetic Compass Orientation in Migratory Birds". PLoS ONE 2 (9): e937. PMC 1976598. PMID 17895978. doi:10.1371/journal.pone.0000937.
- ↑ Deutschlander, ME; Phillips, JB; Borland, SC (1999). "The case for light-dependent magnetic orientation in animals" (PDF). J. Exp. Biol. 202 (8): 891–908. PMID 10085262.
- ↑ Thorup, Kasper (2004). "Reverse migration as a cause of vagrancy" (PDF). Bird Study 51 (3): 228–238. doi:10.1080/00063650409461358. Arquivado dende o orixinal (PDF) o 25 de maio de 2017. Consultado o 21 de agosto de 2018.
- ↑ Kasper Thorup; Thomas Alerstam; Mikael Hake; Nils Kjellén (2003). "Bird orientation: compensation for wind drift in migrating raptors is age dependent". Proc. Royal Soc. Lond. B 270 (Suppl 1): S8–S11. PMC 1698035. PMID 12952622. doi:10.1098/rsbl.2003.0014.
- ↑ Guillemain, M.; Sadoul, N.; Simon, G. (2005). "European flyway permeability and abmigration in Teal Anas crecca, an analysis based on ringing recoveries". Ibis 147: 688–696. doi:10.1111/j.1474-919X.2005.00446.x.
- ↑ "Operation migration". Arquivado dende o orixinal o 29 de abril de 2006. Consultado o 21 de agosto de 2018.
- ↑ "Wisconsin Whooping Crane Management Plan" (PDF). Wisconsin Department of Natural Resources. 6 de decembro de 2006.
- ↑ Rohwer S; Butler LK & DR Froehlich (2005). "Ecology and Demography of East-West Differences in Molt Scheduling of Neotropical Migrant Passerines". En Greenberg R & Marra PP. Birds of two worlds: the ecology and evolution of migration. Johns Hopkins University Press. p. 94. ISBN 0-8018-8107-2.
- ↑ Hedenström, A. (2008). "Adaptations to migration in birds: behavioural strategies, morphology and scaling effects". Philosophical Transactions of the Royal Society B 363 (1490): 287–299. PMC 2606751. PMID 17638691. doi:10.1098/rstb.2007.2140.
- ↑ Weber, Jean-Michel (2009). "The physiology of long-distance migration: extending the limits of endurance metabolism" (PDF). J. Exp. Biol. 212 (Pt 5): 593–597. PMID 19218508. doi:10.1242/jeb.015024.
- ↑ Pulido, F. (2007). "The genetics and evolution of avian migration". BioScience 57 (2): 165–174. doi:10.1641/b570211.
- ↑ J. Rappole; B. Helm; M. Ramos (2003). "An integrative framework for understanding the origin and evolution of avian migration". Journal of Avian Biology 34: 125. doi:10.1034/j.1600-048x.2003.03170.x.
- ↑ B. Mila; T. Smith; R. Wayne. (2006). "Postglacial population expansion drives the evolution of long-distance avian migration in a songbird". Evolution 60 (11): 2403–2409. PMID 17236431. doi:10.1111/j.0014-3820.2006.tb01875.x.
- ↑ Alerstam, Thomas (2001). "Detours in bird migration" (PDF). Journal of Theoretical Biology 209 (3): 319–331. PMID 11312592. doi:10.1006/jtbi.2001.2266. Arquivado dende o orixinal (PDF) o 02 de maio de 2015. Consultado o 21 de agosto de 2018.
- ↑ Jenni L. & Kery M. (2003). "Timing of autumn bird migration under climate change: advances in long-distance migrants, delays in short-distance migrants". Proceedings of the Royal Society B 270 (1523): 1467–1471. PMC 1691393. PMID 12965011. doi:10.1098/rspb.2003.2394.
- ↑ Both, Christiaan; Sandra Bouwhuis; C. M. Lessells; Marcel E. Visser (2006-05-04). "Climate change and population declines in a long-distance migratory bird". Nature 441 (7089): 81–83. ISSN 0028-0836. PMID 16672969. doi:10.1038/nature04539.
- ↑ Wormworth, J.; Mallon, K. (2006). Bird Species and Climate Change: The Global Status Report version 1.0. WWF.
- ↑ Smith RP Jr; Rand PW; Lacombe EH; Morris SR; Holmes DW; Caporale DA (1996). "Role of bird migration in the long-distance dispersal of Ixodes dammini, the vector of Lyme disease". J. Infect. Dis. 174 (1): 221–4. PMID 8656000. doi:10.1093/infdis/174.1.221.
- ↑ Rappole, J.H.; Hubálek, Zdenek (2006). "Birds and Influenza H5N1 Virus Movement to and within North America". Emerging Infectious Diseases 12 (10): 10. doi:10.3201/eid1210.051577. hdl:10088/875.
- ↑ Rappole, J.H.; Derrickson, S.R.; Hubalek, Z. (2000). "Migratory birds and spread of West Nile virus in the Western Hemisphere". Emerging Infectious Diseases 6 (4): 319–328. PMC 2640881. PMID 10905964. doi:10.3201/eid0604.000401. hdl:10088/364.
- ↑ Figuerola, O.; Green, A.J. (2002). "Dispersal of aquatic organisms by waterbirds: a review of past research and priorities for future studies". Freshwater Biology 47 (3): 483–494. doi:10.1046/j.1365-2427.2002.00829.x.
- ↑ Cruden, R. W. (1966). "Birds as Agents of Long-Distance Dispersal for Disjunct Plant Groups of the Temperate Western Hemisphere". Evolution (Evolution, Vol. 20, No. 4) 20 (4): 517–532. JSTOR 2406587. doi:10.2307/2406587.
- ↑ Ydenberg, Ronald C.; Butler, Robert W.; Lank, David B.; Smith, Barry D.; Ireland, J. (2004). "Western sandpipers have altered migration tactics as peregrine falcon populations have recovered" (PDF). Proceedings of the Royal Society B 271 (1545): 1263–1269 1263. PMC 1691718. PMID 15306350. doi:10.1098/rspb.2004.2713.
- ↑ Greenwood, Jeremy J. D. (2007). "Citizens, science and bird conservation". J. Ornithol . 148 (Suppl 1): S77–S124. doi:10.1007/s10336-007-0239-9.
- ↑ Spencer, R. (1985) Marking. In: Campbell. B. & Lack, E. 1985. A dictionary of birds. British Ornithologists' Union. London, pp. 338–341.
- ↑ "Radar Ornithology: Introduction". Clemson University Radar Ornithology Laboratory. Arquivado dende o orixinal o 11 de xuño de 2015. Consultado o 15 de xuño de 2014.
- ↑ "Tracking Cuckoos to Africa ... and back again". British Trust for Ornithology. Consultado o 15 de xuño de 2014.
- ↑ Schmidt, M.; Aschwanden, J.; Liechti, F.; Wichmann, G.; Nemeth, E. (2017). "Comparison of visual bird migration counts with radar estimates". Ibis 159 (3): 491–497. doi:10.1111/ibi.12473.
- ↑ Keith Hobson; Leonard Wassenaar (1997). "Linking breeding and wintering grounds of neotropical migrant songbirds using stable hydrogen isotopic analysis of feathers". Oecologia 109: 142–148. doi:10.1007/s004420050068.
- ↑ Gabriel Bowen; Leonard Wassenaar; Keith Hobson (2005). "Global application of stable hydrogen and oxygen isotopes to wildlife forensics". Oecologia 143 (3): 337–348. PMID 15726429. doi:10.1007/s00442-004-1813-y.
- ↑ Farnsworth, A.; Gauthreaux, S.A.; and van Blaricom, D. (2004). "A comparison of nocturnal call counts of migrating birds and reflectivity measurements on Doppler radar" (PDF). Journal of Avian Biology 35 (4): 365–369. doi:10.1111/j.0908-8857.2004.03180.x. Arquivado dende o orixinal (PDF) o 25 de maio de 2017. Consultado o 21 de agosto de 2018.
- ↑ Liechti, F. (1996). Instructions to count nocturnal bird migration by watching the full moon. Schweizerische Vogelwarte, CH-6204 Sempach, Switzerland.
- ↑ Lowery, G.H. (1951). "A quantitative study of the nocturnal migration of birds". University Kan. Pub. Mus. Nat. Hist. 3: 361–472.
- ↑ Emlen, S. T. & Emlen, J. T. (1966). "A technique for recording migratory orientation of captive birds". Auk 83 (3): 361–367. JSTOR 4083048. doi:10.2307/4083048.
- ↑ Alerstam, 1993. p.352
- ↑ "Migratory bird Treaty 16 USC 703-711; 40 Stat. 755". Legal Information Institute (LII). Cornell Law School.
- ↑ "African-Eurasian Migratory Waterbird Agreement". Arquivado dende o orixinal o 16 de decembro de 2007. Consultado o 21 de agosto de 2018.
- ↑ "The Passenger Pigeon". Smithsonian Institution. Arquivado dende o orixinal o 13 de marzo de 2012. Consultado o 2013-05-24.
- ↑ Shimazaki, Hiroto; Masayuki Tamura & Hiroyoshi Higuchi (2004). "Migration routes and important stopover sites of endangered oriental white storks (Ciconia boyciana) as revealed by satellite tracking" (PDF). Mem Natl Inst. Polar Res., Spec. Issue 58: 162–178. Arquivado dende o orixinal (PDF) o 21 de agosto de 2018. Consultado o 21 de agosto de 2018.
- ↑ Catry, P., Encarnacao, V., Araujo, A., Fearon, P., Fearon, A., Armelin, M. & Delaloye, P. (2004). "Are long-distance migrant passerines faithful to their stopover sites?" (PDF). Journal of Avian Biology 35 (2): 170–181. doi:10.1111/j.0908-8857.2004.03112.x.
- ↑ "Siberian Crane fact sheet". Arquivado dende o orixinal o 10 de novembro de 2007. Consultado o 21 de agosto de 2018.
- ↑ "Fish and Wildlife Service- Bird Mortality Fact sheet" (PDF). Arquivado dende o orixinal (PDF) o 23 de febreiro de 2021. Consultado o 21 de agosto de 2018.
- ↑ Mayntz, Melissa. "Threats to Migrating Birds". About.com Birding. Arquivado dende o orixinal o 12 de xullo de 2014. Consultado o 19 de xuño de 2014.
- ↑ Murray, N. J.; Clemens, R. S.; Phinn, S. R.; Possingham, H. P.; Fuller, R. A. (2014). "Tracking the rapid loss of tidal wetlands in the Yellow Sea". Frontiers in Ecology and the Environment 12: 267–72. doi:10.1890/130260.
- ↑ MacKinnon, J.; Verkuil, Y.I.; Murray, N.J. (2012). IUCN situation analysis on East and Southeast Asian intertidal habitats, with particular reference to the Yellow Sea (including the Bohai Sea). Occasional Paper of the IUCN Species Survival Commission No. 47. Gland, Switzerland and Cambridge, UK: IUCN. p. 70. ISBN 978-2-8317-1255-0.
Véxase tamén
[editar | editar a fonte]Bibliografía
[editar | editar a fonte]- Alerstam, Thomas (2001). "Detours in bird migration" (PDF). Journal of Theoretical Biology 209 (3): 319–331. PMID 11312592. doi:10.1006/jtbi.2001.2266. Arquivado dende o orixinal (PDF) o 02 de maio de 2015. Consultado o 21 de agosto de 2018.
- Alerstam, Thomas (1993). Bird Migration. Cambridge University Press. ISBN 0-521-44822-0. (publicado primeiramente en 1982 como Fågelflyttning, Bokförlaget Signum)
- Berthold, Peter (2001). Bird Migration: A General Survey (2nd ed.). Oxford University Press. ISBN 0-19-850787-9.
- Bewick, Thomas (1797–1804). History of British Birds (1847 ed.). Newcastle: Beilby and Bewick.
- Dingle, Hugh (1996). Migration: The Biology of Life on The Move. Oxford University Press.
- Hobson, Keith; Wassenaar, Leonard (2008). Tracking Animal Migration with Stable Isotopes. Academic Press. ISBN 978-0-12-373867-7.
- Weidensaul, Scott (1999). Living On the Wind: Across the Hemisphere With Migratory Birds. Douglas & McIntyre.
- White, Gilbert (1898) [1789]. The Natural History of Selborne. Walter Scott.
Ligazóns externas
[editar | editar a fonte]- Lindsay, Bethany (3 de abril de 2006). "The compasses of birds". The Science Creative Quarterly. (en inglés)
- Bontadina, F.; Arlettaz, R. (2003). "A heap of feathers does not make a bat’s diet" (PDF). Functional Ecology (17): 141–145. Predación dos morcegos sobre aves migrantes. (en inglés)
- McNamara, John M; Houston, Alasdair I (2008-01-27). "Introduction. Adaptation to the annual cycle" [Introdución.Adaptación ao ciclo anual]. Philosophical Transactions of the Royal Society B: Biological Sciences (en inglés) 363 (1490): 209–210. ISSN 0962-8436. PMC 2042533. PMID 17681915. doi:10.1098/rstb.2007.2192. (en inglés)
- Rura da vía migratoria do leste de Asiay Arquivado 14 de xuño de 2013 en Wayback Machine. en Olango Wildlife Sanctuary, estación de "repostaxe" das aves migratorias.
- Grupo de investigación sobre a migración das aves da Universidade de Lund, Suecia(en inglés)
- Rede de coordinación da migración canadense (coordina as estacións de monitorización da migración do Canadá) (en inglés)
- Investigación das Aves en Science Daily- con varios artigos sobre migración das aves (en inglés)
- The Nature Conservancy's Migratory Bird Program (en inglés)
- BBC Supergoose – gansos rastreados por satélite (en inglés)
- Recensión do libro deGessner, David (2007). Soaring with Fidel : an osprey odyssey from Cape Cod to Cuba and beyond. Boston: Beacon Press. ISBN 9780807085783. OCLC 71581899, que segue a migración anual de aves desde Cabo Cod a Cuba e Venezuela (en inglés)
- Birder's Journal: A Morning With MigrantsNationalgeographic.com
- Global Register of Migratory Species – características de aves e outros vertebrados migratorios (en inglés)
- eBird.com Occurrence Maps – Mapas da migración de varias especies nos Estados Unidos (en inglés)
- Smithsonian Migratory Bird Center Arquivado 04 de febreiro de 2013 en Wayback Machine. (en inglés)
Bases de datos on line
[editar | editar a fonte]- Trektellen.org – Rexistros de contaxes e anelamentos de aves migrantes de todo o mundo.
- Migraction.net – Base de datos interactiva con información en tempo real sobre migración de aves (Francia)