Gliazelle
Gliazelle (kurz Glia) ist ein Sammelbegriff für Zellen im Nervengewebe, die sich strukturell und funktionell von den Nervenzellen (Neuronen) abgrenzen lassen.
Nach heutigen Erkenntnissen bilden Gliazellen nicht nur ein Stützgerüst für Nervenzellen, sondern sorgen auch durch ihre Umhüllung für deren elektrische Isolation. Weiterhin sind Gliazellen maßgeblich an Stofftransport und Flüssigkeitsaustausch sowie an der Aufrechterhaltung der Homöostase im Gehirn beteiligt. Darüber hinaus wirken sie auch im Prozess der Informationsverarbeitung, -speicherung und -weiterleitung mit, wobei die Menge des von ihnen gebildeten Myelins entscheidend ist.[1]
Etwa die Hälfte der Zellen im menschlichen Gehirn sind Gliazellen, ähnlich wie bei anderen Primaten.[2] Gliazellen sind meist kleiner als die Nervenzellen, aber im Unterschied zu diesen variiert ihre durchschnittliche Zellmasse im Nervengewebe nur gering bei verschiedenen Säugetierspezies. In deren Hirnstrukturen hängt das jeweilige Verhältnis von Glia zu Neuronen nach Anzahl und Volumen hauptsächlich von der durchschnittlichen Neuronengröße ab.[3]
Fast alle Gliazellen stammen (wie die Nervenzellen) vom ektodermalen Keimblatt ab, genauer vom Neuroektoderm (Neuroglia); lediglich die Mikroglia (Mesoglia) sind mesodermalen Ursprungs.
Geschichte
[Bearbeiten | Quelltext bearbeiten]Der Entdecker der Gliazellen war Mitte des 19. Jahrhunderts Rudolf Virchow. Er vermutete eine Stütz- und Haltefunktion und gab den Zellen deshalb den Namen Gliazellen, abgeleitet aus dem griechischen Wort glia für „Leim“.
Ende des 19. Jahrhunderts gelang es Santiago Ramón y Cajal, Pío del Río Hortega und Camillo Golgi, sie mittels unterschiedlicher Silberimprägnation (Golgi-Färbung) noch detaillierter zu klassifizieren.
Gliazelltypen
[Bearbeiten | Quelltext bearbeiten]Es werden verschiedene Typen unterschieden. Im Zentralnervensystem finden sich:
- Neuroglia, weiter unterteilt in:
- Astroglia (Astrozyten)
- Oligodendroglia (Oligodendrozyten)
- Ependymzellen
- Plexusepithelzellen
- Radialglia
- Mikroglia (Mesoglia, Hortega-Zellen, residentielle Makrophagen)
- Müllerzellen in der Netzhaut
Im peripheren Nervensystem finden sich:
- Schwann-Zellen (Periphere Glia)
- Mantelzellen (Satellitenzellen, Amphizyten)
- Teloglia der motorischen Endplatte
Darüber hinaus werden den Gliazellen folgende Zellen zugerechnet:
- Stützzellen des Sinnesepithels
- Pituizyten sind Gliazellen, die nur in der Neurohypophyse zu finden sind (sie beeinflussen Transport, Speicherung und Freigabe der Hormone in den Nervenfasern)
Astroglia
[Bearbeiten | Quelltext bearbeiten]Die Astroglia (Astrozyten) bilden die Mehrheit der Gliazellen im zentralen Nervensystem von Säugetieren. Es sind sternförmig verzweigte Zellen, deren Fortsätze Grenzmembranen zur Gehirnoberfläche (bzw. Pia mater) und zu den Blutgefäßen bilden.
Es sind zwei Typen von Astrozyten bekannt:
- Faserglia (Astrocytus fibrosus – auch: Langstrahler), fibrillenreich, vor allem in der weißen Substanz. Im Elektronenmikroskop durch zahlreiche Mikrotubuli und intrazelluläre Faserstrukturen charakterisiert
- Protoplasmatische Glia (Astrocytus protoplasmaticus – auch: Kurzstrahler) vor allem in der grauen Substanz
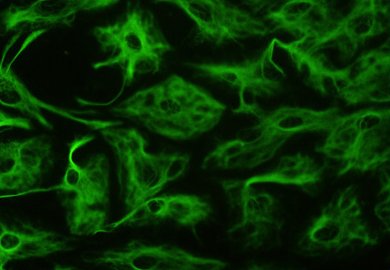
Astrozyten sind maßgeblich an der Flüssigkeitsregulation im Gehirn beteiligt und sorgen für die Aufrechterhaltung des Kalium-Haushaltes. Die während der Erregungsleitung in Nervenzellen frei werdenden Kalium-Ionen werden vor allem durch eine hohe Kalium-Leitfähigkeit und zum Teil auch durch K+- und Cl−-Kotransporter in die Gliazellen aufgenommen. Damit regulieren sie auch den extrazellulären pH-Haushalt im Gehirn.
Astrozyten sind tragendes Element des Mikrokreislaufs zur Abfallentsorgung in Gehirn und Rückenmark (ZNS) des 2012 entdeckten glymphatischen Systems. Liquor, der über den Virchow-Robin-Raum rund um die Arterien in alle Bereiche des ZNS gelangt, wird über Endfüßchen der Astrozyten direkt vom Virchow-Robin-Raum aufgenommen, im Zellzwischenraum verteilt und am Ende – unter Mitnahme von Abfallstoffen – entlang der Außenwände der Venen wieder aus dem ZNS ausgeschwemmt.[4][5]
Astrozyten nehmen an der Informationsverarbeitung am Gehirn teil. Sie enthalten in Vesikeln Glutamat, das bei seiner exozytotischen Freisetzung benachbarte Neurone aktiviert.
Astrozyten bilden nach Durchtrennung der Axone von Nervenzellen „Glianarben“, die maßgeblich daran beteiligt sind, das neuerliche Auswachsen der Axone zu verhindern. Dies stellt ein zentrales Problem für Patienten mit Querschnittlähmung dar.
In Astrozyten kommt als Marker das Intermediärfilament GFAP (glial fibrillary acidic protein, „saures Gliafaserprotein“) vor, welches somit zum Nachweis von zentralnervösem Gewebe z. B. in Fleischprodukten verwendet werden kann, was insbesondere in Hinblick auf BSE an Bedeutung gewonnen hat. Die Bildung des Proteins wird durch krankhafte Veränderungen im Hirngewebe verstärkt.
Eine spezielle Form von Astrozyten ist die Radialglia, deren Hauptausläufer z. B. die Molekularschicht in etwa parallel durchziehen und in Endfüßen an der Pia mater enden. Sie spielen eine wichtige Rolle als Leitstrukturen in der frühen Hirnentwicklung von Vertebraten (Wirbeltieren). Im ausgereiften (Säuger-)Gehirn sind sie nur noch im Kleinhirn (Bergmannglia) und in der Retina (Müllerglia) vorhanden.
Oligodendroglia
[Bearbeiten | Quelltext bearbeiten]Oligodendroglia (Oligodendrozyten) bilden das Myelin, die elektrische Isolation der Axone der Nervenzellen im Gehirn bzw. Zentralnervensystem. Sie entsprechen damit den Schwannschen Zellen im peripheren Nervensystem. Allerdings unterscheiden sie sich prinzipiell, denn ein Oligodendrozyt kann Axonabschnitte mehrerer Nervenzellen umwickeln, während eine myelinbildende Schwannsche Zelle immer nur ein neuronales Axon umwickelt. Die Evolution von Oligodendrozyten wird als Voraussetzung der Großhirnentwicklung bei Chordatieren angesehen.
Astroglia und Oligodendroglia werden von manchen Autoren auch unter Makroglia zusammengefasst in Abgrenzung zur:
Mikroglia
[Bearbeiten | Quelltext bearbeiten]Mikrogliazellen (auch als Hortega-Zellen oder Mesoglia bezeichnet) machen ca. 20 % aller Gliazellen aus. Sie sind der einzige Zelltyp des Parenchyms des zentralen Nervensystems (ZNS), bei dem es sich weder um eine neuronale, noch um eine vaskuläre Zelle handelt, vielmehr stellen die mikroglialen Zellen die residenten (ansässigen) inflammatorischen Zellen des ZNS dar. Eine Besonderheit der Mikrogliazellen ist, dass es sich bei ihnen sowohl um Gliazellen des ZNS als auch um einen einzigartigen Typ mononukleärer Phagozyten handelt. Dabei fungieren sie nicht nur als Fresszellen der Immunabwehr im ZNS, sondern sorgen auch für die richtige Anzahl neuronaler Vorläuferzellen während der Entwicklung des ZNS.[6][7]
Da Antikörper die Blut-Hirn-Schranke nicht passieren können, stehen Mikrogliazellen für die Hauptform der aktiven Immunabwehr im ZNS. Ihre Aufgabe besteht in der Erkennung und Beseitigung potenzieller pathogen wirkender Substanzen. Sie schützen so in erster Linie die nicht regenerierbaren Neuronen des ZNS vor irreversiblen Schäden. Durch die Vermittlung inflammatorischer Immunantworten unterstützen sie darüber hinaus die Nervenzellen bei der Regeneration nach einer Verletzung.[6] Sie haben mithin eine ähnliche Funktion wie Makrophagen in anderen Geweben, da sie Zellreste abgestorbener Nervenzellen und Oligodendrozyten durch Phagozytose beseitigen. Mikroglia entstehen in der Embryonalentwicklung aus Vorläuferzellen im Dottersack und nicht wie die restlichen Zellen des Nervensystems aus der Neuralleiste und dem Neuralrohr, also dem Ektoderm.[8] Zu den Antigen-präsentierenden Zellen gehörend, ist für ihre Funktion eine molekulare Aktivierung notwendig. So hat man beispielsweise eine Aktivierung nach Schädeltraumata, bei Erkrankungen wie der multiplen Sklerose oder bei erblichen Leukodystrophien beobachtet. Auffällig ist u. a. das Territorialverhalten der Mikroglia: Zwischen zwei Zellen ist immer ein gewisser Abstand vorhanden.[9]
Auch Schizophreniepatienten haben deutlich mehr aktivierte Mikrogliazellen im Gehirn als Gesunde.[10][11]
Morphologie
[Bearbeiten | Quelltext bearbeiten]Ein Hinweis auf die Zugehörigkeit der Mikroglia zum monozytären Phagozytensystem sind ihr Vorkommen als ruhende und aktive Zellen. Dieses Verhalten wird bei Makrophagen ebenfalls beobachtet.
- Ruhende Mikroglia weisen heterochromatinreiche Zellkerne auf sowie elektronendichtes Zytoplasma. Hier befinden sich neben typischen Organellen vor allem Lysosomen und Vimentinfilamente als Bestandteile des Zytoskeletts. Die Zellform ist geprägt durch feinen, unregelmäßigen Fortsatzreichtum.
- Aktive Mikroglia reagieren auf Verletzungen des ZNS mit Hypertrophie und Proliferation. Sie unterscheiden sich von den inaktiven Formen durch stärker entwickelte Fortsätze.
Funktion
[Bearbeiten | Quelltext bearbeiten]Reaktive Mikroglia weisen charakteristische Verhaltensweisen auf. Nach ihrer Aktivierung kommt es zur Ansammlung der Zellen am Ort der Läsion, was durch die Fähigkeit zur amöboiden Fortbewegung ermöglicht wird. Anschließend werden durch Phagozytose bzw. Exozytose zytotoxischer Stoffe wie Wasserstoffperoxid oder Stickstoffmonoxid abgestorbene Zellsubstanzen und Fremdkörper beseitigt. Nach Abbau von defekten körpereigenen und fremden Bestandteilen geben sie spezifische Zytokine (Interleukin-1, Tumor-Nekrose-Faktor α, Interferon-γ) in den Extrazellularraum ab, was die Astrozytenproliferation und die Bildung von Glianarben initiiert, wodurch weitere Immunreaktionen inhibiert werden.
Ependym
[Bearbeiten | Quelltext bearbeiten]Ependymzellen bilden die einschichtige Auskleidung des Hohlraumsystems im Zentralnervensystem.
Erkrankungen
[Bearbeiten | Quelltext bearbeiten]Durch Störung der Entwässerungsfunktion der Astrozyten oder Schädigung der Blut-Hirn-Schranke kann ein Hirnödem entstehen. Häufigere Tumoren sind Gliome wie das Astrozytom, das Oligodendrogliom und das Glioblastom.
Siehe auch
[Bearbeiten | Quelltext bearbeiten]Literatur
[Bearbeiten | Quelltext bearbeiten]- Helmut Kettenmann, B. R. Ransom: Neuroglia. Oxford University Press, New York 2012, ISBN 978-0-19-979459-1.
Weblinks
[Bearbeiten | Quelltext bearbeiten]- Network Glia Eine Homepage mit vielen Informationen zu Neuroglia
- Mikrogliazellen auf die Finger geschaut, Max-Planck-Gesellschaft (www.mpg.de)
- Neue Einblicke ins Gehirn – Beobachtung von Gliazellen in der intakten Hirnrinde, Max-Planck-Gesellschaft (www.mpg.de)
- Gliazellen bei der Bildung von Synapsen entscheidend, Pressetext Austria (www.pressetext.at)
- Briefträger, Botenstoffe und der unterschätzte Klebstoff, Telepolis (www.heise.de) – Artikel von Telepolis
- Unterschätzter Nervenkitt
- Astrozyten steuern Neurone
- Spezifische Zellfunktionen der sogenannten Astroglia aufgedeckt
Einzelnachweise
[Bearbeiten | Quelltext bearbeiten]- ↑ Gliazellen entscheidend für die zeitliche Verarbeitung akustischer Signale im Gehirn. In: Ärzteblatt News vom 20. Dezember 2020.
- ↑ F. Azevedo, L. Carvalho, L. Grinberg, J. Farfel, R. Ferretti, R. Leite, W. Filho, R. Lent, S. Herculano-Houzel: Equal numbers of neuronal and nonneuronal cells make the human brain an isometrically scaled-up primate brain. In: J Comparative Neurology. Band 513. Jahrgang, Nr. 5, April 2009, S. 532–541, doi:10.1002/cne.21974, PMID 19226510.
- ↑ Bruno Mota, Suzana Herculano-Houzel: All brains are made of this: a fundamental building block of brain matter with matching neuronal and glial masses. In: Frontiers in Neuroanatomy. 8. Jahrgang, Nr. 127, November 2014, PMC 4228857 (freier Volltext).
- ↑ N. A. Jessen, A. S. Munk, I. Lundgaard, M. Nedergaard: The Glymphatic System: A Beginner's Guide. In: Neurochemical research. Band 40, Nummer 12, Dezember 2015, S. 2583–2599, doi:10.1007/s11064-015-1581-6, PMID 25947369, PMC 4636982 (freier Volltext) (Review).
- ↑ D. Raper, A. Louveau, J. Kipnis: How Do Meningeal Lymphatic Vessels Drain the CNS? In: Trends in neurosciences. Band 39, Nummer 9, September 2016, S. 581–586, doi:10.1016/j.tins.2016.07.001, PMID 27460561, PMC 5002390 (freier Volltext) (Review).
- ↑ a b R. M. Ransohoff, A. E. Cardona: The myeloid cells of the central nervous system parenchyma. In: Nature. Band 468, Nr. 7321, 2010, S. 253–262, PMID 21068834.
- ↑ C. L. Cunningham, V. Martínez-Cerdeño, S. C. Noctor: Microglia regulate the number of neural precursor cells in the developing cerebral cortex. In: The Journal of neuroscience : the official journal of the Society for Neuroscience. Band 33, Nummer 10, März 2013, S. 4216–4233, ISSN 1529-2401. doi:10.1523/JNEUROSCI.3441-12.2013. PMID 23467340.
- ↑ Florent Ginhoux, Shawn Lim, Guillaume Hoeffel, Donovan Low, Tara Huber: Origin and differentiation of microglia. In: Frontiers in Cellular Neuroscience. Band 7, 2013, ISSN 1662-5102, doi:10.3389/fncel.2013.00045.
- ↑ H. Lassmann, F. Zimprich, K. Vass, W. F. Hickey: Microglial cells are a component of the perivascular glia limitans. In: J. Neurosci. Res. 28, 1991, S. 236–243. PMID 2033652.
- ↑ Georg Juckel, Marie Pierre Manitz, Martin Brüne, Astrid Friebe, Michael T. Heneka, Rainer J. Wolf: Microglial activation in a neuroinflammational animal model of schizophrenia — a pilot study. In: Schizophrenia Research. Band 131, Nr. 1–3, 2011, S. 96–100, ISSN 0920-9964, doi:10.1016/j.schres.2011.06.018, PMID 21752601.
- ↑ Meike Drießen: Psychisch krank durch Dauerstress: Der Zusammenhang zwischen Stress, Immunsystem und psychischen Erkrankungen. In: Rubin Wissenschaftsmagazin. Ruhr Universität Bochum, 3. November 2014, abgerufen am 22. November 2014.