Astrocit
| Astrocit | |
|---|---|
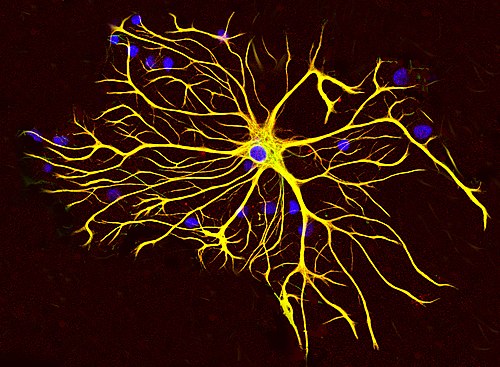 Astrocitna ćelija stanica moždanog tkiva pacova, uzgojena u kulturi i obojena antitijelima za GFAP (crveno) i vimentin (zeleno). Oba proteina su prisutna u velikim količinama u intermedijarnim vlaknima ove ćelije, tako da se ćelija vidi kao žuta. Plavi materijal pokazuje DNK koja je vizuelizirana bojom DAPI, a otkriva jedro astrocita i ostale ćelije. Slika je priložena ljubaznošću EnCor Biotechnology Inc. | |
| Detalji | |
| Identifikatori | |
| Latinski | 'Astrocytus' |
| MeSH | D001253 |
| NeuroLex ID | sao1394521419 |
| TH | TH H2.00.06.2.01008 {{{2}}}.html HH2.00.06.2.00002, H2.00.06.2.01008 .{{{2}}}.{{{3}}} |
| FMA | 54537 |
| Anatomska terminologija | |
Astrociti grč. astron = zvijezda + "kyttaron" = ćelija, šupljina) – poznata i kao astroglija – je karakteristična zvjezdasta ćelija glije, u mozgu i kičmenoj moždini.[1] [2][3] Relativni udio astrocita u mozgu nije dobro definiran. Ovisno o primijenjenoj tehnici brojanja, studije su otkrile da taj udio varira od regije do regije, a kreće se od 20% do 40% ukupne glije.[4] One obavljaju mnoge funkcije, uključujući i biohemijsku podršku endotelnim ćelijama za formiranje krvno-moždane barijere, dobavljanje hranjivih tvari do nervnog tkiva, održavanje vanćelijske ionske ravnoteže, kao i ulogu u procesa popravke ožiljaka mozga i kičmene moždine nakon traumatskih povreda.
Istraživanja od sredine 1990-ih su pokazala da astrociti propagiraju međućelijste talase Ca2+ over lona duge distance u odgovoru na stimulaciju, i, slično neuronima, otpoštaju neurotransmitere (zvane gliotransmiteri), na način koji oisi o Ca2+. Podaci također ukazuju da astrociti također daju signale neuronima tokom otpuštanja glutamata koji zavisi od Ca2+.[5] Takva otkrića su napravili istraživanja na astrocitim važnim područjem neuronauke.

Struktura
[uredi | uredi izvor]Astrociti su pod-tip ćelija glije i centralnog nervnog sistema. Također su poznati i kao astrocitne glijne ćelije. Kako su u obliku zvijezde, njihovi mnogi nastavci i ovojnice preve sinapse sa neuronima. Astrociti klasično sr identificiraju pomoću histološke analize; mnoge od ovih ćelija ispoljavaju intermedijerni filament glijinih vlaknastih kiselih proteina (GFAP).[6] U centralnom nervnom sistemu postoji ekoliko oblika astrocita, uključujući vlaknaste (bijela masa), protoplazmatske (siva masa), i radijalne. Vlaknasta glija obično se nalazi u bijeloj masi, sa relativno malo organela, a imauju duge nerazgranate ćelijske nastavke. Ovaj tip često ima "vaskularne noge", koje fizički povezuju ćelije sa vanjske strane kapilarnih zidova kada su u njihovoj blizini. Ćelije protoplazmatske glije su najrasprostranjenije, a nalaze se u tkivu sive mase, posjeduju veću količinu organela i imuju kratke i vrlo razgranate tercijarne nastavke. Radijalne ćelije glije su smještene u ravninama koje su okomite na osima komora. Jedan od njihovih nastavaka oslanja se na pia mater, a drugi je duboko zaronjen i sivu masu. Ćelije radijalne glije su uglavnom prisutne tokom razvoja, kada imaju ulogu u migracijama neurona. Müllerove ćelije u mrežnjači i ćelije Bergmannove glije moždane kora predstavljaju iznimku, jer su prisutne još i u odrasloj dobi. Kada su u blizini ovojnice pia mater, sva tri oblika astrocita imaju nastavke za formiranje membrane pija-glije.
Razvoj
[uredi | uredi izvor]
Astrociti su makroglijske ćelije centralnog nervnog sistema. Izvedeni su iz heterogene skupine progenitornih ćelija u neuroepitelu u razvijajućem centralnom nervnom sistemu. Nedavni radovi, sažeti u komentaru Rowitcha i Kriegsteina,[7] ukazuju na to da postoji izuzetna sličnost između poznatih genetičkih mehanizama koji preciziraju loze različitih neuronskih podtipova makroglijinih ćelija. Baš kao i kod specifikacije neuronskih ćelija, faktori kanonske signalizacije, kao što su sonični jež (SHH), hepatocitni faktor rasta (FGFs), Wnt i koštani morfogenetski proteini (BMP), pružaju pozicijske informacije za razvoj tokom morfogenih gradienata u smjerovima osovina dorzalno–ventralo, naprijed–natrag i medijalno–lateralno. Rezultanta se ispoljava u segmentaciji neuroepitela u progenitorne domene (p0, p1 p2, p3 and pMN) za razne tipove neurona i razvoju kičmene moždine. Na osnovu nekoliko studija, sada se vjeruje da se ćelija makroglije nastaju tokom specijalizacije ćelija. Studije koje je sa suradnicima objavio Hoechst, pokazale su da iz astrocita proizilaze tri različite skupine domena p1, p2 i p3.[8] Podtipovi asterocita mogu biti identificirani na osnovu ispoljavanja različitih transcripcijskih faktora (PAX6, NKX6.1) i markera ćelijske površine (reelin i SLIT1). Identificirane su tri podtipske skupine:
- 1) dorzalna, na eritricitima VA1, izvedenim iz domena p1, koja ispoljava PAX6 i reelin,
- 2) ventralni eritrociti VA3, izvedeni iz p3, koji ispoljavaju NKX6.1 i SLIT1 i
- 3) intermedjarni bijele mase, VA2 astrociti, izvedeni iz domena p2, koji ispoljavaju PAX6, NKX6.1, reelin i SLIT1.[9] Nakon pojave specijalizacije astrocita u razvoju CNS-a, vjeruje se da njihovi prekursori migriraju do konačne pozicije unutar živčanog sistema prije završnogh procesa diferencijacije.
Funkcija
[uredi | uredi izvor]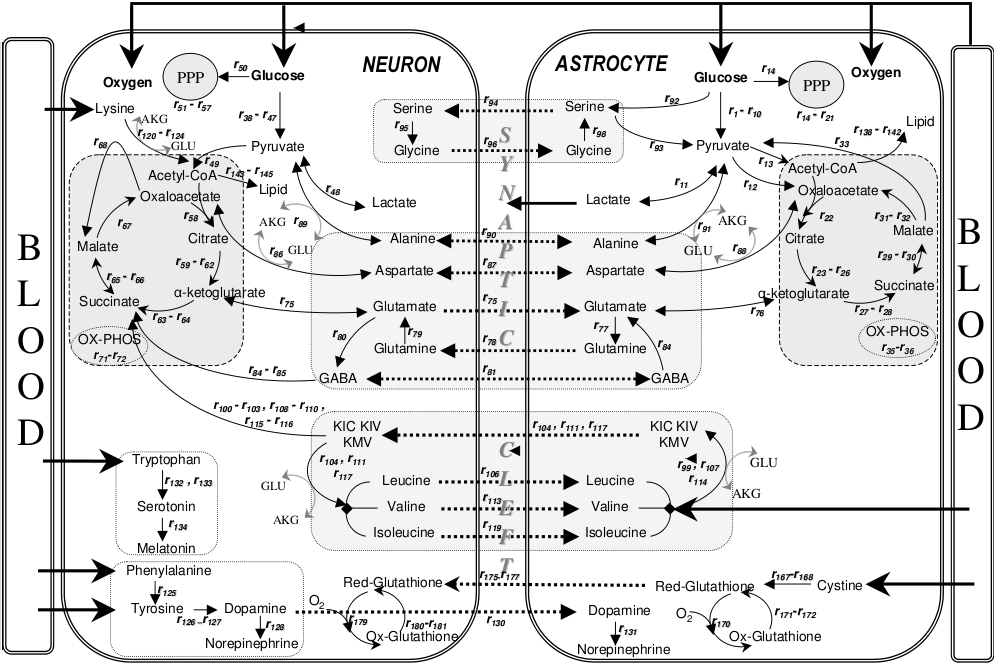
Ranije je u medicinskoj nauci, tvorba neuronske mreže smatrala se jedinom važnom funkcijom astrocita, a oni su viđeni kao punila jaza. U novije vrijeme, ponovo je razmatrana funkcija astrocita, pa se za njih sada mislili da imaju brojne aktivnr uloge u mozgu, uključujući i lučenje ili apsorpciju neuronskih predajnika i održavanje krvno-moždane barijere.[11] Nadovezujući se na ovu ideju predložen je koncept "tripartitne sinapse", pozivajući se na uske veze koje se javljaju na sinapsi među presinapsnog, postsinapsnog i glijinog elementa.[12]
- Strukturne: Astrociti su uključeni u fizičko strukturiranje mozga, gdje su "u obliku zvijezde". Najzastupljenije su glija-ćelije u mozgu. koje su usko povezane sa neuronims sinapse. Reguliraju prijenos električnih impulsa u mozgu.
- Rezervni pufer rezervi glikogenskog goriva: Astrociti sadrže glikogen i sposobni su za glikogenezu. Astrociti slijede neurone u čeonoj kori i hipokampusu, gdje pohranjuju i otpuštaju glikogen. Dakle, astrociti mogu podstaknuti živčane ćelije s glukozom tokom razdoblja visoke stope potrošnje glukoze i nedostatka glukoze. Nedavna istraživanja pokazuju da možda postoji veza između ove aktivnosti i vježbanja.[13]
- Podrška metabolizma: Astrociti snabdijevaju neurone hranljivim tvarima kao što je laktat.
- Krvno-moždana barijera: Astrociti na kraju nastavaka okružuju endotelne ćelije, gdje (mislilo se) pomažu u održavanju krvno-moždane barijere, ali nedavna istraživanja pokazuju da oni tu nemaju značajnu ulogu. Umjesto toga, oni su uski spojevi i bazne lamine moždanih endotelneđih ćelija koje imaju najznačajnu ulogu u održavanju barijere.[14] Međutim, nedavno je pokazano da je aktivnost astrocita vezana za protok krvi u mozgu i da je to ono što se zapravo meri u snimcimas MRI.[15][16]
- Preuzimanje i otpuštanje transmitera: Astrociti imaju plazmamembranske transportere, kao što je glutamatni transporter, za nekoliko neurotransmitera, uključujući i glutamat ATP i GABA. U novije vrijeme, astrociti su pokazali da oslobađaju glutamat ili vezikule adenozin-trifosfata (ATP), na način koji zavisi od Ca2+.[17] (This has been disputed for hippocampal astrocytes.)[18]
- Regulacija kalijevih iona u vanćelijskom prostoru: Astrociti imaku kalijeve kanale visoke gustoće. Kada su neuroni aktivni, oni oslobađaju kalij, povećavajući njegovu lokalnu vanćelijsku koncentraciju. Pošto su astrociti vrlo propusni za kalij, brzo čiste višak nakupina u vanćelijski prostor.[19]
Ako je ova funkcija ometana, vanćelijska koncentracija kalija će rasti, što dovodi do depolarizacije neurona, prema Goldmanovoj jednadžbi. Za abnormalno nakupljanje vanćelijskog kalija je dobro znano da rezultira epilepsijskim aktivnošću neurona.[20]
- Modulacija sinapsne transmisije: U nadočnom jezgru hipotalamusa, javljaju se brze promjene morfologije astrocita, što izaziva heterosinapsni prijenos impulsa između neorona.[21] U hipokampusu, astroiti potiskuju sinapsni prijenos, otpuštanjem ATP, koji se hidrolizira pomoću ektonukliotidaza, dajući adenozin. Adenozin djeluje na nervne receptore adenozina i inhibira transmisiju sinapse, čime se povećava dinamički raspon dostupan za LTP.[22]
- Vazomodulacija: Astrociti mogu pomagati kao posrednici u nervnoj regulaciji protoka krvi.[23]
- Podsticanje mijelinacijske aktivnosti oligodendrocita: Električna aktivnost u neuronima uzrokuje da otpuštaju ATP, koji služi kao važan podsticaj za formiranje mijelina. Međutim, ATP ne djeluje direktno na oligodendrocite. Umjesto toga, uzrokuje da astrociti luče inhibitorni faktor citokina leukemije (LIF), regulatorni protein koji podstiče mijelinizirajuću aktivnost oligodendrocita. To sugerira da astrociti u mozgu imaju izvršno-koordinirajuću ulogu.[24]
- Popravke nervnog sistema: Nakon ozljede živčanih ćelija u centralnom živčanom sistemu, astrociti popunjavaju prostor za formiranje ožiljka glije, popravjeno područje transformira u neurone.[25]
- Dugoročna potencijacija: Naučnici su dugo raspravljati o tome da li astrociti integriraju učenje i pamćenje i hipokampusu. Nedavno je pokazano da ubacivanje progeneatora ljudskih glija predačkih ćelijama u mozak novorođrnih miše podstiče ćelije da se diferenciraju u astrocite. Nakon diferencijacija ove ćelije povećavaju LTP i poboljšavaju performanse memorije u miševa.[26]
Astrociti su povezani putem jaznih međućelijskih veza, stvarajući električno spojeni (funkcijski) sincicij.[27] Zbog takve sposobnosti astrociti da komuniciraju sa svojim susjedima, promjene u njihovoj aktivnosti mogu imati posljedice na aktivnosti drugih koje su prilično udaljeni od originalnog astrocita.
Priliv Ca2+ iona u astrocite je bitna promjena koja u konačnici generira talse kalcija. Budući da je ovaj priliv direktno izazvan povećanjem protoka krvi u mozak, evi talasi su, kao neka vrsta hemodinamskog funkcijskog odgovora. Povećanje koncentracije unutarćelijskog kalcija se može širiti prema van duž tog funkcijskog sincicija. Mehanizmi širenja talasa kalcija su difuzija iona kalcija i IP3 preko jaznog spoja i vanćelijskog ATP signala.[28] Podizanje razine kalcija je primarna poznata osa aktivacije astrocita, a neophodno je i dovoljno za neke vrste astrocitnog oslobađanja glutamata.[29]
S obzirom na važnost kalcija u signalizaciju astrocita, razvijeni su uski regulatorni mehanizmi za napredovanje u prostorno-vremenskoj signalizaciji kalcija. Preko matematičke analize pokazalo se da lokaliziran priliv iona Ca2+ doprinosi lokaliziranoj pojavi citosolne koncentracije iona Ca2+.[30] Šta više, akumulacija citoplazmatskog Ca2+ je nezavisna od unutarćelijskog protoka kalcija, a zavisi od razmjene Ca2+ preko membrane, citosolne difuzije kalcija, ćelijske geometrijel, kolebanja vanćelijskog kalcija i njegovih početnih koncentracija.[30]
Klasifikacija
[uredi | uredi izvor]Postoji nekoliko različitih mačina klsifikacije astrocita.
Vezanje i antigenski fenotip
[uredi | uredi izvor]Oni su zasnovani na klasičnom djelu Raffa i suradnika, početkom 1980-ih, proučavanjem živaca pacova.
- Tip 1: Antigeni Ran2+, GFAP+, FGFR3+, A2B5−, nalik je na "tip 1 astrocita" 7. dana postporođajnog vidnog živca pacova. Može nastati iz glijinog ograničenog prekursora tropotencijskih ćelija (GRP), ali ne i iz bipotencijskih O2A/OPC (oligodendrocit, tip 2 astrocitnih prekursora, koji se nazivaju i oligodendrocitne pro stagenmitorne ćelije).
- Tip 2: Antigeni A2B5+, GFAP+, FGFR3−, Ran 2−. Ove ćelije razvijaju se in vitro i od tripotencijskih GRP (vjerovatno preko faze O2A) ili od bipotencijskih ćelija O2A (za koje neki misle da su izvedene iz GRP) ili in vivo, ove progenitorske ćelije transplantiraju u oštećena mjest (ali vjerovatno ne u normalnom razvoju, najmanje ne u vidnom živcu). Ovaj tip astrocita je glavna astrocitna komponenta u kulturi postnatalnih ćelija očnog živca, koje se generiraju rastom O2A ćelija u prisustvu goveđeg fetusnog seruma, ali ne tokom razvoja in vivo.[31]
Anatomska klasifikacija
[uredi | uredi izvor]- Protoplazmatsi astrociti nalaze se u sivoj masi i imaju mnogo granatih nastavaka koji se ugrađuju u sinapse. Newki se generiraju iz multipotentne supventrikulske zone progenitorskih ćelija.[32][33]
- Gömöri-pozitivni astrociti su podskup protoplazmatskih astrocita koji sadrže brojne citoplalmatske inkluzije, ili granule, koje se oboje Gömörijevom bojom zvanom hrom-alum hematoksilin. Sada je poznato da su ove granule formirane od ostataka degeneriranih mitohondrija zahvaćenih u lizosomima.[34] Jedan tip odgovoran je za oštećenja mitohondrija, što je u tim specijaliziranim astrocitima neka vrsta oksidativnog stresa. Gömöri-pozitivni astrociti su mnogo više zastupljeni u lučnom jezgru hipotalamusa i hipokampusu nego u ostalim dijelovima mozga. Imaju ulogu u regulaciji odgovora hipotalamusa na glukozu.[35][36]
- Vlaknasti astrociti nađeni su u bijeloj masi i imaju duge tanke nastavke, koji na krajevima utastaju u Ranvijeove čvorove. Neki od njih su generalizirani kao radijalna glija.[37][38][39][40][41]
Klasifikacija transporter/receptor
[uredi | uredi izvor]- GluT tip ispoljva se kao glutamatski transporteri (EAAT1/SLC1A3 i EAAT2/SLC1A2), a odgovoran je za sinaosno otpuštanje glutamata pomoću postojećeg transportera. Funkcija i dostupnost EAAT2 su modulirani putem TAAR1 – unutarćelijskog receptora u ljudkim ćelijama.[42]
Prekomjerna ekspresija TAAR1 značajno smanjuje razinu EAAT-2 i glutamata koji su dodatno reducirani metioninom (Meth). Uzevši zajedno, raspolooživi podaci pokazuju da Meth tretman aktivira TAAR1 dovodeći do pojave međućelijskog cAMP-a u ljudskim astrocitima i moduliranih sposobnosti klirensa glutamata. Nadalje, molekulske promjene razina TAAR1 u astrocitima odgovaraju na promjene u razinama astrocitnih EAAT-2 i funkcija.
- GluR tip se ispoljava kao glutamatni receptori (pretežno mGluR i AMPA tip) i odgovara za otpuštanje glutamata u sinapsama što je posredovano kanalima sinapsi i IP3-zavisnim Ca2+ medijatorima.
Također pogledajte
[uredi | uredi izvor]Reference
[uredi | uredi izvor]- ^ Sofradžija A., Šoljan D., Hadžiselimović R. (2004). Biologija 1. Svjetlost, Sarajevo. ISBN 9958-10-686-8.CS1 održavanje: više imena: authors list (link)
- ^ Korene Z., Hadžiselimović R., Maslić E. (2001). Biologija 8. Sarajevo: Svjetlost. ISBN 9958-10-396-6.CS1 održavanje: upotreba parametra authors (link)
- ^ Međedović S., Maslić E., Hadžiselimović R. (2002). Biologija 2. Svjetlost, Sarajevo. ISBN 9958-10-222-6.CS1 održavanje: više imena: authors list (link)
- ^ Verkhratsky, A.; Butt, A.M. (2013). "Numbers: how many glial cells are in the brain?". Glial Physiology and Pathophysiology. John Wiley and Sons. str. 93–96. ISBN 978-0-470-97853-5.
- ^ Fiacco TA, Agulhon C, McCarthy KD (oktobar 2008). "Sorting out Astrocyte Physiology from Pharmacology". Annu. Rev. Pharmacol. Toxicol. 49 (1): 151–74. doi:10.1146/annurev.pharmtox.011008.145602. PMID 18834310.CS1 održavanje: upotreba parametra authors (link)
- ^ Sarma, PotukuchiVenkata Gurunadha Krishna; Srikanth, Lokanathan; Vengamma, Bhuma; Venkatesh, Katari; Chandrasekhar, Chodimella; Mouleshwara Prasad, BodapatiChandra; Sanjeevkumar, Akula (2013). "In vitro differentiation of cultured human CD34+ cells into astrocytes". Neurology India. 61 (4): 383–8. doi:10.4103/0028-3886.117615. PMID 24005729.
- ^ Rowitch, David H.; Kriegstein, Arnold R. (2010). "Developmental genetics of vertebrate glial–cell specification". Nature. 468 (7321): 214–22. doi:10.1038/nature09611. PMID 21068830.
- ^ Muroyama, Y; Fujiwara, Y; Orkin, SH; Rowitch, DH (2005). "Specification of astrocytes by bHLH protein SCL in a restricted region of the neural tube". Nature. 438 (7066): 360–363. doi:10.1038/nature04139. PMID 16292311.
- ^ Hochstim, C; Deneen, B; Lukaszewicz, A; Zhou, Q; Anderson, DJ (2008). "Identification of positionally distinct astrocyte subtypes whose identities are specified by a homeodomain code". Cell. 133 (3): 510–522. doi:10.1016/j.cell.2008.02.046. PMC 2394859. PMID 18455991.
- ^ Çakιr, Tunahan; Alsan, Selma; Saybaşιlι, Hale; Akιn, Ata; Ülgen, Kutlu Ö (2007). "Reconstruction and flux analysis of coupling between metabolic pathways of astrocytes and neurons: application to cerebral hypoxia". Theoretical Biology and Medical Modelling. 4 (1): 48. doi:10.1186/1742-4682-4-48. PMC 2246127. PMID 18070347.
- ^ Kolb, Brian and Whishaw, Ian Q. (2008) Fundamentals of Human Neuropsychology. Worth Publishers. 6th ed. ISBN 0716795868
- ^ Araque A, Parpura V, Sanzgiri RP, Haydon PG (1999). "Tripartite synapses: glia, the unacknowledged partner". Trends in Neuroscience. 22 (5): 208–215. doi:10.1016/S0166-2236(98)01349-6. PMID 10322493.CS1 održavanje: upotreba parametra authors (link)
- ^ Reynolds, Gretchen (22. 2. 2012). "How Exercise Fuels the Brain". New York Times.
- ^ Kimelberg HK, Jalonen T, Walz W (1993). "Regulation of the brain microenvironment:transmitters and ions.". u Murphy, S (ured.). Astrocytes: pharmacology and function. San Diego, CA: Academic Press. str. 193–222. ISBN 0125113706.CS1 održavanje: upotreba parametra authors (link)
- ^ Swaminathan N (2008). "Brain-scan mystery solved". Scientific American Mind: 7.
- ^ Figley CR, Stroman PW (2011). "The role(s) of astrocytes and astrocyte activity in neurometabolism, neurovascular coupling, and the production of functional neuroimaging signals". European Journal of Neuroscience. 33 (4): 577–588. doi:10.1111/j.1460-9568.2010.07584.x. Nepoznati parametar
|last-author-amp=zanemaren (prijedlog zamjene:|name-list-style=) (pomoć)CS1 održavanje: upotreba parametra authors (link) - ^ Santello M, Volterra A (2008). "Synaptic modulation by astrocytes via Ca2+-dependent glutamate release". Neuroscience. Mar. 158 (1): 253–9. doi:10.1016/j.neuroscience.2008.03.039. PMID 18455880.CS1 održavanje: upotreba parametra authors (link)
- ^ Agulhon C, Fiacco T, McCarthy K (2010). "Hippocampal short- and long-term plasticity are not modulated by astrocyte Ca2+ signaling". Science. 327 (5970): 1250–1257. doi:10.1126/science.1184821. PMID 20203048.CS1 održavanje: upotreba parametra authors (link)
- ^ Walz W (2000). "Role of astrocytes in the clearance of excess extracellular potassium". Neurochemistry International. 36 (4–5): 291–300. doi:10.1016/S0197-0186(99)00137-0.
- ^ Gabriel S, Njunting M, Pomper JK, Merschhemke M, Sanabria ER, Eilers A, Kivi A, Zeller M, Meencke HJ, Cavalheiro EA, Heinemann U, Lehmann TN (2004). "Stimulus and Potassium-Induced Epileptiform Activity in the Human Dentate Gyrus from Patients with and without Hippocampal Sclerosis". The Journal of Neuroscience. 24 (46): 10416–10430. doi:10.1523/JNEUROSCI.2074-04.2004. PMID 15548657.CS1 održavanje: upotreba parametra authors (link)
- ^ Piet R, Vargová L, Syková E, Poulain D, Oliet S (2004). "Physiological contribution of the astrocytic environment of neurons to intersynaptic crosstalk". Proceedings of the National Academy of Sciences of the United States of America. 101 (7): 2151–5. doi:10.1073/pnas.0308408100. PMC 357067. PMID 14766975.CS1 održavanje: upotreba parametra authors (link)
- ^ Pascual O, Casper KB, Kubera C, Zhang J, Revilla-Sanchez R, Sul JY, Takano H, Moss SJ, McCarthy K, Haydon PG (2005). "Astrocytic purinergic signaling coordinates synaptic networks". Science. 310 (5745): 113–6. doi:10.1126/science.1116916. PMID 16210541.CS1 održavanje: upotreba parametra authors (link)
- ^ Parri R, Crunelli V (2003). "An astrocyte bridge from synapse to blood flow". Nature Neuroscience. 6 (1): 5–6. doi:10.1038/nn0103-5. PMID 12494240.CS1 održavanje: upotreba parametra authors (link)
- ^ Ishibashi T, Dakin K, Stevens B, Lee P, Kozlov S, Stewart C, Fields R (2006). "Astrocytes Promote Myelination in Response to Electrical Impulses". Neuron. 49 (6): 823–32. doi:10.1016/j.neuron.2006.02.006. PMC 1474838. PMID 16543131.CS1 održavanje: upotreba parametra authors (link)
- ^ http://lenta.ru/news/2014/10/12/cvi/
- ^ Han, Xiaoning; Chen, Michael; Wang, Fushun; Windrem, Martha; Wang, Su; Shanz, Steven; Xu, Qiwu; Oberheim, Nancy Ann; Bekar, Lane (3. 7. 2013). "Forebrain Engraftment by Human Glial Progenitor Cells Enhances Synaptic Plasticity and Learning in Adult Mice". Cell Stem Cell (jezik: English). 12 (3): 342–353. doi:10.1016/j.stem.2012.12.015. ISSN 1934-5909. PMC 3700554. PMID 23472873.CS1 održavanje: nepoznati jezik (link)
- ^ Bennett M, Contreras J, Bukauskas F, Sáez J (2003). "New roles for astrocytes: gap junction hemichannels have something to communicate". Trends Neurosci. 26 (11): 610–7. doi:10.1016/j.tins.2003.09.008. PMID 14585601.CS1 održavanje: upotreba parametra authors (link)
- ^ Newman, E. A. (2001). "Propagation of intercellular calcium waves in retinal astrocytes and Müller cells". The Journal of neuroscience : the official journal of the Society for Neuroscience. 21 (7): 2215–23. PMC 2409971. PMID 11264297.
- ^ Parpura V, Haydon P (2000). "Physiological astrocytic calcium levels stimulate glutamate release to modulate adjacent neurons". Proceedings of the National Academy of Sciences of the United States of America. 97 (15): 8629–34. doi:10.1073/pnas.97.15.8629. PMC 26999. PMID 10900020.CS1 održavanje: upotreba parametra authors (link)
- ^ a b Lopez-Caamal, F.; Oyarzun, D.A.; Middleton, R.H.; Garcia, M.R. (maj 2014). "Spatial Quantification of Cytosolic Ca2+ Accumulation in Nonexcitable Cells:An Analytical Study". IEEE/ACM Transactions on Computational Biology and Bioinformatics. 11 (3): 592–603. doi:10.1109/TCBB.2014.2316010.
- ^ Fulton, B. P.; Burne, J. F.; Raff, M. C. (1992). "Visualization of O-2A progenitor cells in developing and adult rat optic nerve by quisqualate-stimulated cobalt uptake". The Journal of neuroscience : the official journal of the Society for Neuroscience. 12 (12): 4816–33. PMID 1281496.
- ^ Levison SW, Goldman JE (1993). "Both oligodendrocytes and astrocytes develop from progenitors in the subventricular zone of postnatal rat forebrain". Neuron. 10 (2): 201–12. doi:10.1016/0896-6273(93)90311-E. PMID 8439409.CS1 održavanje: upotreba parametra authors (link)
- ^ Zerlin M, Levison SW, Goldman JE (1995). "Early patterns of migration, morphogenesis, and intermediate filament expression of subventricular zone cells in the postnatal rat forebrain". J. Neurosci. 15 (11): 7238–49. PMID 7472478.CS1 održavanje: upotreba parametra authors (link)
- ^ Brawer JR; Stein, Robert; Small, Lorne; Cissé, Soriba; Schipper, Hyman M. (1994). "Composition of Gomori-positive inclusions in astrocytes of the hypothalamic arcuate nucleus". Anatomical Record. 240 (3): 407–415. doi:10.1002/ar.1092400313. PMID 7825737.
- ^ Young, J. K.; McKenzie, J. C. (2004). "GLUT2 immunoreactivity in Gomori-positive astrocytes of the hypothalamus". Journal of Histochemistry and Cytochemistry. 52 (11): 1519–24. doi:10.1369/jhc.4A6375.2004. PMC 3957823. PMID 15505347.
- ^ Marty N (2005). "Regulation of glucagon secretion by glucose transporter type 2 (glut2) and astrocyte-dependent glucose sensors". J. Clinical Investigation. 115 (12): 3545–3553. doi:10.1172/jci26309.
- ^ Choi BH, Lapham LW (1978). "Radial glia in the human fetal cerebrum: a combined Golgi, immunofluorescent and electron microscopic study". Brain Res. 148 (2): 295–311. doi:10.1016/0006-8993(78)90721-7. PMID 77708.CS1 održavanje: upotreba parametra authors (link)
- ^ Schmechel DE, Rakic P (1979). "A Golgi study of radial glial cells in developing monkey telencephalon: morphogenesis and transformation into astrocytes". Anat. Embryol. 156 (2): 115–52. doi:10.1007/BF00300010. PMID 111580.CS1 održavanje: upotreba parametra authors (link)
- ^ Misson JP, Edwards MA, Yamamoto M, Caviness VS (novembar 1988). "Identification of radial glial cells within the developing murine central nervous system: studies based upon a new immunohistochemical marker". Brain Res. Dev. Brain Res. 44 (1): 95–108. doi:10.1016/0165-3806(88)90121-6. PMID 3069243.CS1 održavanje: upotreba parametra authors (link)
- ^ Voigt T (novembar 1989). "Development of glial cells in the cerebral wall of ferrets: direct tracing of their transformation from radial glia into astrocytes". J. Comp. Neurol. 289 (1): 74–88. doi:10.1002/cne.902890106. PMID 2808761.
- ^ Goldman SA, Zukhar A, Barami K, Mikawa T, Niedzwiecki D (august 1996). "Ependymal/subependymal zone cells of postnatal and adult songbird brain generate both neurons and nonneuronal siblings in vitro and in vivo". J. Neurobiol. 30 (4): 505–20. doi:10.1002/(SICI)1097-4695(199608)30:4<505::AID-NEU6>3.0.CO;2-7. PMID 8844514.CS1 održavanje: upotreba parametra authors (link)
- ^ Cisneros IE, Ghorpade A (oktobar 2014). "Methamphetamine and HIV-1-induced neurotoxicity: role of trace amine associated receptor 1 cAMP signaling in astrocytes". Neuropharmacology. 85: 499–507. doi:10.1016/j.neuropharm.2014.06.011. PMID 24950453.
Dopunska literatura
[uredi | uredi izvor]- White, F.A.; Jung, H.; Miller, R.J. (2007). "Chemokines and the pathophysiology of neuropathic pain". Proceedings of the National Academy of Sciences of the United States of America. 104 (51): 20151–20158. doi:10.1073/pnas.0709250104. PMC 2154400. PMID 18083844. Nepoznati parametar
|last-author-amp=zanemaren (prijedlog zamjene:|name-list-style=) (pomoć) - Milligan, E.D.; Watson, L.R. (2009). "Pathological and protective roles of glia in chronic pain". Neuron-Glia Interactions. 10 (1): 23–36. doi:10.1038/nrn2533. PMC 2752436. PMID 19096368. Nepoznati parametar
|last-author-amp=zanemaren (prijedlog zamjene:|name-list-style=) (pomoć) - Watkins, L.R.; Milligan, E.D.; Maier, S.F. (2001). "Glial activation: a driving force for pathological pain". Trends in Neurosci. 24 (8): 450–455. doi:10.1016/S0166-2236(00)01854-3. PMID 11476884. Nepoznati parametar
|last-author-amp=zanemaren (prijedlog zamjene:|name-list-style=) (pomoć) - Freeman, M. R. (2010). "Specification and Morphogenesis of Astrocytes". Science. 330 (6005): 774–8. doi:10.1126/science.1190928. PMID 21051628.
- Verkhratsky, A.; Butt, A.M. (2013). "Numbers: how many glial cells are in the brain?". Glial Physiology and Pathophysiology. John Wiley and Sons. str. 93–96. ISBN 978-0-470-97853-5.
Vanjski linkovi
[uredi | uredi izvor]- Cell Centered Database – Astrocyte
- "Astrocytes" Arhivirano 21. 11. 2012. na Wayback Machine at Society for Neuroscience
- NIF Search – Astrocyte via the Neuroscience Information Framework