Протеин
- Белтък пренасочва насам. За частта на яйцето вижте Яйчен белтък.

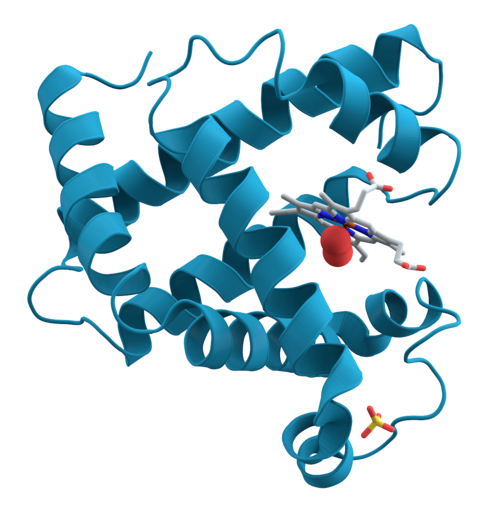
Протеинът, още белтък или белтъчина, е събирателен термин за биологически важни макромолекули, изградени чрез поликондензирането на отделни аминокиселини.
Белтъците са един от основните градивни компоненти на живите клетки, както и на вирусните частици. Притежават сложна пространствена структура и изпълняват разнообразни биологични функции – от типично структурни, защитни (активна и пасивна), транспортни и съкратителни до каталитични и регулаторни.
Цялата информация, необходима за изграждането на белтъчната молекула, е кодирана в ДНК. Посредством процес на точно презаписване на информацията в РНК (транскрипция) и превеждане от полинуклеотидна в аминокиселинна последователност (транслация) е възможно синтезирането на определен белтък, притежаващ конкретна биологична роля.
Номенклатура и синоними
[редактиране | редактиране на кода]Българските наименования „белтък“ и „белтъчина“ произлизат от традиционното наименование на яйчния белтък, който съдържа белтъчини за развитието на зародиша. Като синоним се използва и чуждицата протеин (protein), срещана в много западноевропейски езици.
История
[редактиране | редактиране на кода]
За първи път през 1789 г. френският химик Антоан Фуркроа прави разграничение между белтъците и другите известни органични субстанции. Класифицира познатите дотогава албумин, фибрин и желатин като самостоятелни съединения с животински произход. От тази начална стъпка до формулирането на концепция за това, какво представляват белтъците, е извървян дълъг път, минаващ през грубо характеризиране на химичния състав, опити за клинично приложение на белтъчни екстракти, до все по-прецизното дефиниране на структурата и функциите на всеки отделен белтък.
Самото понятие е въведено през 1838 година, когато в писмо от 10 юли, изпратено от Йонс Якоб Берцелиус до Герардус Йоханес Мулдер, за първи път е употребено названието „протеин“ с акцент върху първостепенното и първичното (от гръцки πρωτειοξ – първична нишка)[1][2]. Точният цитат от писмото е:
- „Le nom protéine que je vous propose pour l’oxyde organique de la fibrine et de l’albumine, je voulais le dériver de πρωτειοξ, parce qu’il paraît être la substance primitive ou principale de la nutrition animale.“[3]
В приблизителен превод цитатът гласи:
- „Наименованието протеин, което Ви предлагам за органичния оксид на фибрина и албумина, бих искал да произлиза от πρωτειοξ, защото изглежда, че това са основни или главни за храненето на животните вещества.“
С това изречение се дефинира за първи път понятието протеин, което се използва като синоним на „белтък“.
Биохимия на протеините
[редактиране | редактиране на кода]
Протеините представляват линейни полимери (поликондензати), изградени от поредица от до 20 различни вида L-α-аминокиселини. Всички аминокиселини притежават общи структурни характеристики, включително α-въглерод, който е свързан с аминогрупа, карбоксилна група, и странична верига, различна за всеки вид аминокиселина. Само пролин се различава от тази основна структура, тъй като тя съдържа необичаен пръстен в N-амино-групата, която поставя CO-NH амида във фиксиран конформация.[4] Страничните вериги на претогенните (канонични) аминокиселини, имат голямо разнообразие на химическите структури и свойства. Комбинираният ефект на всички странични вериги на аминокиселините в един протеин в крайна сметка определя неговата триизмерна структура, химическа реактивност и биологична функция.[5] Аминокиселините се свързват в полипептидна верига посредством пептидна връзка. Веднъж свързана във веригата на протеин, индивидуалната аминокиселина се нарича остатък, а поредицата от въглеродни, азотни и кислородни атоми е известна като основна верига или гръбнак на протеина.[6]

Пептидната връзка има две резонансни форми, за които способстват няколко двойни връзки и възпрепятстват въртенето около оста ѝ, така че α-въглеродните атоми са приблизително в една равнина. Другите два ъгъла в полипептидната верига, между α-въглеродните атоми и двете съседни пептидни групи (ъгли на Рамачандран), определят локалната форма на протеиновия гръбнак.[6] Краят на протеина със свободна карбоксилна група е познат като C-края или карбокси край, а краят със свободна аминогрупа е известен като N-края или аминокрай. Термините протеин, полипептид и пептид са малко двусмислени и могат да се застъпват по смисъл. Протеин се използва обикновено за биологична молекула в стабилна конформация, докато пептид обикновено се отнася за кратки аминокиселинни олигомери, при които често липсва стабилна триизмерна структура. Въпреки това, границата между двете не е добре дефинирана и обикновено се намира в близост до 20 – 30 аминокиселинни остатъка.[7] Полипептид може да се отнася до всяка една линейна верига от аминокиселини, обикновено независимо от дължината, но често предполага липсата на определена конформация.
Синтез
[редактиране | редактиране на кода]Протеините се изграждат от аминокиселини, използвайки информацията, кодирана в гените. Всеки протеин има своя собствена уникална аминокиселинна последователност, която се определя от нуклеотидната последователност на гена кодиращ този протеин. Генетичният код е набор от триплетни комплекти, наречени кодони и всяка комбинация от три нуклеотида определя аминокиселина, например триплетът AUG (аденин-урацил-гуанин) е кодон за метионин. Тъй като ДНК съдържа четири различни нуклеотида, общият брой на възможните кодони (триплети) е 64, следователно, има известен излишък в генетичния код. Някои аминокиселини се кодират от повече от един кодон.[8] Гените, кодирани в ДНК, първо се транскрибират в пре-иРНК от протеини като РНК-полимеразата. Повечето организми след това обработват пре-иРНКите (известни като първичен транскрипт), използвайки различни форми на посттранскрипционни модификации, за да формират зряла иРНК, която след това се използва като матрица за синтеза на протеини от рибозомата. При прокариотите иРНК може да се транслира веднага след началото на транскрипцията или да бъде свързана с рибозома, след като се отдалечи от бактериалната хромозома. За разлика от тях, еукариотите синтезират иРНК в ядрото на клетката и след това преминават през ядрената мембрана в цитоплазмата, където се извършва синтезът на протеини. Скоростта на синтеза на протеини е по-висок в прокариоти отколкото при еукариоти и може да достигне до 20 аминокиселини в секунда.[9]
Процесът на синтезиране на белтък от иРНК матрица е известен като транслация. иРНК се товарят на рибозомата и се четат по три нуклеотида в даден момент чрез разпознаване на всеки кодон от иРНК с антикодон от тРНК, която носи аминокиселината, съответстваща на кодона, когото тя разпознава. Ензимът аминоацил-тРНК синтетаза свързва тРНК молекулите с правилните аминокиселини. Растящият полипептид често се нарича зараждащата се верига. Протеините винаги се синтезират от N-края към C-края.[8]
Размерът на синтезирания протеин може да се измерва с броя на аминокиселините, които съдържа, и от общата му молекулна маса, която обикновено се отчитат в далтони (синоним с атомни единици за маса), или производната килодалтон (kDa). Дрождевите протеини са със средно 466 аминокиселини дълго и маса 53 kDa.[7] Най-големите известни протеини са титините, компонент на мускулния саркомер, с молекулна маса от почти 3000 kDa и обща дължина от почти 27 000 аминокиселини.[10]
Химичен синтез
[редактиране | редактиране на кода]Къси протеини могат да бъдат синтезирани химически чрез съвкупност от методи известни като пептиден синтез, които използват органични техники за синтез като химическо легиране, за да се произвеждат пептиди с висока ефективност.[11] Химичният синтез дава възможност за въвеждане на не-естествени аминокиселини в полипептидните вериги, като такива съдържащи флуоресцентни сонди като страничен радикал на аминокиселина.[12] Тези методи са полезни в лабораторната практика в биохимията и клетъчната биология, но като цяло не се използват за търговски приложения. Химичният синтез е неефективен за полипептиди по-дълги от около 300 аминокиселини, а синтезираните протеини не могат лесно да приемат естествената си, функционално активна третична структура. Повечето химични методи за синтез, изграждат полипептидната верига от C-края на N-края, в противовес на биохимичната реакция.[13]
Пост-транслационни модификации
[редактиране | редактиране на кода]
След завършване на транслацията и освобождаването на протеина от рибозомата, аминокиселините в полипептидните верига са изложени на разнообразни химични изменения, известни като пост-транслационни модификации. Те биват многообразни по своята химична природа и в огромна степен начертават биологичната съдба на протеина, която обаче, е предопределена на първо място от неговата първична структура. Пост-транслационните модификации биват:
- Добавяне на различни функционални групи (ацетилни, метилни, фосфатни и други групи)
- Присъединяване на липиден или въглехидратен компонент
- Замяна на стандартни аминокиселини с нестандартни (образуване на цитрулина)
- Формиране на структурни промени (образуване на дисулфидни мостове между цистеиновите остатъци);
- Премахване от протеина на различни сигнални последователности или частична протеолоза на пробелтъка във функционално активен (например триспиноген и трипсин);
- Добавяне на малки протеини, които влияят на разграждането на протеина (убиквитинилиране).
Тези модификации може да бъдат универсални (добавяне на вериги, състоящи се от мономери на убиквитин, сигнал за влошаване на този протеин и насочване към протеазома), както и специфични за протеина.[14].
В същото време, един и същ протеин може да претърпи много и различни пост-транслационни модификации. Например, хистоните (протеини, които са част от хроматин и в еукариоти) в различни условия може да имат до 150 различни модификации.[15].
Белтъчно сортиране
[редактиране | редактиране на кода]Синтезираните от рибозоми в цитоплазмата протеини трябва да попадат в различни отделения на клетката – ядро, митохондрии, ендоплазмен ретикулум, както и апарат на Голджи, лизозоми, и други; както и някои протеини, трябва се интегрират в клетъчната мембрана или да бъдат насочени към извънклетъчната среда. За да се насочи към определен компартмент, протеинът трябва да има специална сигнална последователност. В повечето случаи, този последователност е част от веригата на аминокиселините на протеините. В някои случаи, тези сигнали са добавени посттранскационно към протеина и представляват олигозахариди компоненти. Транспортът на протеини към ендоплазмения ретикулум се извършва паралелно с техния синтез, като сигналната последователност се разпознава от транслокационен комплекс изграден от РНК и протеинени, който свързва рибозомата и я придвижва до ЕПР. От ЕПР протеините могат да преминат в апарата Голджи и след това до лизозоми или към външната мембрана или екстрацелуларната среда постредством везикуларния транспорт. Протеините предназначени да функционират в клетъчното ядро имат сигнална последователност NLS (nuclear localisation signal) за насочване към ядрото, като се въвеждат през ядрените пори. В митохондриите и хлоропластите протеините попадат при наличие на сигнални последователности, които ги насочват към специфични протеини пори-транслокатори с участието на шаперони.
Структура на белтъците
[редактиране | редактиране на кода]
Основният компонент на всички белтъци са аминокиселини, подредени последователно в дълги полипептидни вериги. Броят на каноничните α-аминокиселини в живата природа е 20, макар че в някои белтъци се срещат и други α-аминолиселини като 3- и 4-хидроксипролин, 5-хидроксилизин и други, така също при определени условия аминокиселината селеноцистеин се кодира от стоп кодона UAG.
Аминокиселините се комбинират в строго определен ред във всяка белтъчна молекула, като броят им варира между 30 до 3000 (понякога и много повече) аминокиселини в протеин. Връзката между отделните аминокиселини е амидна, но поради някои свои особености е наречена със специфично име – пептидна връзка. Затова и самите полимерни вериги се означават като полипептидни. Организацията в пространството на полипептидните вериги е изключително сложна и характерна за всеки отделен белтък. За прегледност и по-лесна ориентация, пространствената структура на белтъците е разделена на отделни нива:
Първично ниво
[редактиране | редактиране на кода]Първично ниво (примерна структура) – представлява точната последователност на свързване на отделните аминокиселинни остатъци, кодирана от полинуклеотидната последователност на ДНК. В природата белтъците не се срещат в първична структура. Те могат да я заемат временно.
Към първичната структура на белтъците се отнася и така наречения дисулфиден мост. Извън пептидната връзка дисулфидният мост е единствената друга ковалентна връзка в белтъчната молекула. Той се формира при свързването на две сулфхидрилни (тиолови) групи на цистеинови остатъци. Свързването се осъществява чрез отделянето на два атома водород (дехидрогениране). Свързаните два цистеинови остатъка образуват един общ цистин. Дисулфидният мост може да бъде както между две отделни полипептидни вериги (междуверижен), така и от два цистеинови остатъка принадлежащи на една верига (вътрешноверижен).[16]
Първичната структура определя останалите равнища на организация и е ключова за свойствата и функциите на белтъка.

Вторично ниво
[редактиране | редактиране на кода]Вторично ниво (секундерна структура) – това са локални нагъвания на основния скелет на полипептидната верига, притежаващи известна периодичност. Всяка възпроизводима и характерна форма се означава като отделен тип вторична структура, като например алфа-спирала, бета-листовидна структура, бета- и гама-завой, както и липса на порядък.[16]
- α-спирала – α-спиралата се получава при възникването на водородни връзки между всяка пептидна група и третата следваща след нея. Това води до огъване на полипептидната верига на мястото на a-въглеродния атом. Един пълен оборот на такава спирала включва 3,6 аминокиселинни остатъка, а ходът и е 0,54 nm. α-спиралата е възможно най-компактната форма на полипептидната верига. Обикновено природните α-спирали са дясно завити. α-спирали могат да се получат и при възникване на водородни мостове между всяка пептидна група и втората или четвъртата след нея, но се срещат сравнително рядко (напр. колаген).[16]

- β-лист – β-лист се формира, когато две сравнително опънати полипептидни вериги се намират близо една до друга и техните пептидни групи образуват междуверижни (вътрешноверижни) водородни мостове. Това е сравнително най-опънатата форма на полипептидна верига. Гръбнакът на полипептидната верига се извива зигзагообразно и наподобява хармоника, в чийто краища се намират a-въглеродните атоми. β-структура се формира в рамките на една полипептидна верига, когато гръбнакът на веригата се извие в обратна посока при наличие на пролинова инверсия. За да формират β-лист няколко вериги те имат антипаралелен ход, тоест ако в дадена посока едната започва с азотния си край, то съседната трябва да започне с въглеродния си край. Макар и рядко срещат и природни β-листи с паралелни вериги.[16]
Третично ниво
[редактиране | редактиране на кода]Третично ниво (терциерна структура) – пълното нагъване на полипептидната верига в пространството, с всички възможни близки и далечни взаимодействия. Това ниво дава представа за цялостната форма на белтъчната молекула, както и за връзките и отношенията между отделните вторични структури. Третичната структура не е резултат от действието на случайни сили, а е строго определена от аминокиселинната последователност (първичната структура) на полипептидната верига, тоест тя е генетично детерминирана. Триизмерната форма, която белтъчната молекула приема на ниво третична структура се означава като конформация. Във физиологични условия конформацията търпи неголеми колебания, възникнали при взаимодействия с други молекули, свързани с функционалните прояви на белтъците. Формата на белтъчната молекула се поддържа от множество слаби нековалентни взаимодействия:
- Водородни връзки – Водородните връзки възникват, когато водороден атом е привлечен от два електоотрицателни атома, като с единия от тях е свързан предварително. При това положение водородния атом е силно привлечен от тях и застава помежду им като мост (връзка). В белтъчните молекули се създават няколко вида водородни връзки. Между азотния атом на една пептидна група и кислородния от друга; фенолната група на тирозина с карбоксилна група (аспаргинова киселина, глутаминова киселина) или карбонилна група от пептидна връзка. Водородните връзки са десетки пъти по-слаби от ковалентните връзки в полипептидната верига, но поради своето изобилие значително способстват за поддържането на конформацията. Енергия на връзката 4,2 – 25 kJ/mol.[16]
- Хидрофобни взаимодействия – Те са сили на привличане възникващи между ненатоватени групи, които при своето доближаване претърпяват преходна поляризация. Максимумът им на действие е при разстояние 0,32 – 0,37 nm, като при по-близки разстояния водят до отблъскване. Поради това, че при нагъване на полипептидната верига на малко разстояние се събират значително количество на полярни групи и силите на привличане са адитивни, те достигат висока стойност и са от съществено значение за поддържането на белтъчната конформация. Енергия на връзката 4,2 kJ/mol.[16]
- Йонни връзки – Това са електростатични сили на привличане между разноименно натоварени атомни групи и такива на отблъскване при едноименни. Тъй като броят и вида на натоварените групи е функция на pH, то и йонните връзки в белтъчната молекула са функция на pH. При значително повишаване или понижаване на pH дисоциацията на отрицателно, респективно положително натоварените групи се увеличава, което води до дисоциация на белтъчните комплекси. Йонните сили на привличане са най-значителни около изоелектричната точка на белтъка. Енергия на връзката 21 kJ/mol.[16]
- Взаимодействия с p-връзки – Получават се, когато ароматни пръстени на фенилаланин, тирозин и триптофан се окажат паралелно разположени между техните p-електрони се зараждат слаби взаимодействия.[16]
- Дипол-диполни взаимодействия – Нямат пряко отношение към поддържането на конформацията, но са важни са взаимодействията с други молекули. Белтъците имат много висок диполен момент: 310 D (дебай) за инсулин, 1100 D за g-глобулин. За сравнение водата има диполен момент 1,8 D.[16]
Четвъртично ниво
[редактиране | редактиране на кода]Четвъртично ниво (кватернерна структура) – това е отново цялостна пространствена форма, но на асоциирани нековалентно белтъчни молекули. Тя се изразява в образуването на олигомерни белтъци, но не всеки белтък се характеризира с това четвъртично равнище. Всяка отделна белтъчна молекула, притежаваща характерна, завършена, третична структура се означава като субединица, а цялостният конгломерат – като олигомер. Не всички белтъци притежават четвъртична структура, тъй като повечето са изградени от една полипептидна верига, а четвъртично ниво предполага наличието на поне две. Типичен пример за белтък с четвъртична структура е хемоглобинът.[16]
Белтъците в разтвор, както нормално съществуват в живите клетки, търпят вариации в структурата си, тъй като са подложени на редица въздействия от различни химични елементи, хормони и други биохимични вещества.
Пространствената структура на белтъка се нарича конформация, а всички изменения, независимо дали са съществени или не, се означават като конформационни промени. Благодарение на пространствената си структура белтъците имат свой активен център и множество алостерични центрове. Тези центрове представляват части от молекулата които са пригодени за връзка с точно определени вещества от клетката (например хормони и т.н.).[16]
Денатурация
[редактиране | редактиране на кода]
Белтъците са молекули много чувствителни към факторите на заобикалящата ги среда. На тяхната функция и структура могат да повлияят множество фактори като например промяна в pH на средата, температурата, налягането, концентрацията на определени вещества (тежки метали, детергенти и други), йонизиращи лъчения. Когато даден протеин бъде подложен на подобна промяна следва денатурация. Това е процес на разкъсване на връзките поддържащи четвъртичната, третичната и вторичната структура. По-податливи на денатурация са глобуларните белтъци, защото техните свойства са много зависими от вторичната и особено третичната им структура. При продължително въздействие на вредния фактор и неговото естество процесът е необратим. При възстановяване на хомеостазата следва обратният процес – ренатурация, при който се възстановява нативната физиологично активна конформация на белтъка. В клетката този процес се спомага от други протеини известни като шаперони и шаперонини. При дълго и продължително въздействие на денатуриращия фактор може да се достигне до необратима денатурация, при което настъпва и утаяване на белтъците, процес известен като коагулация. Коагулацията и денатурацията, обаче не са тъждествени процеси, така например нативни белтъци могат да се утаят (при процес известен като изсолване), както и не всички денатурирали белтъци задължително се утаяват. Прозаичен пример е термичната обработка на кокоше яйце, при което консистенцията се променя. Но в този случай не става въпрос единствено за денатурация, тъй като системата е многокомпонентна.
При денатурация белтъчната молекула преминава от по-подредено към по-безпорядъчно състояние, с нарастваща ентропия и следователно привидно по-изгодно от термодинамична гледна точка. При процеса на ренатурация няма противоречие със законите на термодинамиката, ако белтъчната молекула не се разглежда сама за себе си, а в система с обкръжаващата я водна фаза.
Денатурацията може да има практическо приложение, като част от имунната защита. Така например при вирусно заболяване в организма се разпространяват множество вирусни частици, в болшинството от случаите изградени и от протеини. Неспецифичната имунната реакция на организма се изразява с покачване на постоянната температура на организма. По този начин белтъците изграждащи вирусните частици денатурират, вследствие на което вирусите се разпадат. Затова не е полезно температура от 37 до 38 да се сваля чрез медикаменти веднага.
Структура на белтъците в живите организми
[редактиране | редактиране на кода]Способността на протеините да възстановяват правилната триизмерна структура след денатурация позволява да се предложи хипотезата, че цялата информация за крайната структура на протеините, се съдържа в аминокиселинната последователност. Сега е широко приетата теория, че в резултат на еволюцията стабилната структура на протеина има минимална свободна енергия в сравнение с други възможни конформации на полипептидната верига.[17].
Въпреки това, в клетките има група протеини, чиято функция е да се гарантира връщането на протеиновата структура след повреда, както и създаването и разпадането на протеиновите комплекси. Тези протеини се наричат шаперони и шаперонини. Концентрацията на много шаперони в клетката се увеличава рязко с покачване на температурата на околната среда. Те се отнасят към групата на HSP (Heat shock proteins, топлинно шокови протеини).[18] Значението на шапероните за нормалното функциониране на организма може да се илюстрира с α-кристалина, в лещата на човешкото око. Мутациите в този протеин водят до помътняване на лещата дължащо се на натрупването на протеини и в резултат на това до катаракта.[19].
Прости и комплексни протеини
[редактиране | редактиране на кода]В структурата на много протеини в допълнение към пептидни вериги са включени и неаминокиселинни компоненти. По този критерий белтъците се разделя на две големи групи – прости и комплексни протеини (протеиди). Простите протеини, съдържат само аминокиселинна верига, а комплексните протеини съдържат различни небелтъчни фрагменти. Тези фрагменти не са с протеинова природа и са наречени „простетични групи“. В зависимост от химическата природа на простетичните групи комплексните протеини се делят на следните класове:
- Гликопротеини, съдържащи ковалентно свързани въглехидратни остатъци и техния подклас – протеогликани свързани с мукополизахариди. Въглехидратните остатъци обикновено са свързани в хидроксилни групи на серин и треонин (О гликозилиране) или аспаргин (N гликозилиране). По-голямата част от екстрацелуларната протеини са гликопротеини. Въглехидрати част от протеогликани, основен компонент на извънклетъчната матрица, е около 95%.
- Липопротеини, съдържащи като небелтъчна част нековалентно свързани липиди. Липопротеините и техните липидни комплекси аполипопротеинте изпълняват функция на липиден транспорт.
- Металопротеини, съдържщи не-хемово координативно свързани метални йони. Сред металопротеините са протеини, които извършват депониране и транспорт на метали (например, феритин и трансферин за желязо) и ензими (например, цинк, съдържащата карбоанхидраза; супероксид дисмутаза съдържаща в активните си центрове на йоните на мед, манган, желязо и/или други метали)
- Нуклеопротеини, съдържащи ковалентно свързани ДНК или РНК, по-специално, хроматин.
- Фосфопротеиди, съдържащи ковалентно свързани остатъци от фосфорна киселина. В образуването на естерната връзка с фосфата са включени хидроксилните групи на серин или треонин. Фосфопротеид е казеина в мляко.
- Хромоптотеини – колективното име на комплексни протеини с цветни групи от различен химичен характер. Те включват много протеини съдържащи порфиринова група, изпълнявайки различни функции – хемопротеини (протеини, съдържащи хем група – хемоглобин, миоглобин, цитохроми и т.н.), хлорофил, флавопротеини с флавонова група и т.н.
Функция на белтъците
[редактиране | редактиране на кода]Всеки белтък притежава специфична функция, като практически няма процес в живите организми, който да не зависи от конкретен протеин. Функциите могат да са най-разнообразни, но формално могат да се обединят в няколко основни:
Трябва да се отбележи, че класификацията на протеините в съответствие с тяхната функция е по-скоро условно, тъй като в еукариотите един и същ протеин може да изпълнява няколко функции. Добре проучен пример за този гъвкавост е lysyl-tRNA синтетазата – ензим, от класа на aminoacyl-tRNA синтетазите, който не само добавя лизин към tRNA, но и регулира транскрипцията на някои гени [20]. Много протеини имат функция, която се осъществява чрез ензимната им активност. Такива са например двигателния протеин миозин, регулаторните протеини протеинкинази, транспортният протеин натриево-калиева помпа и други.
Каталитична функция
[редактиране | редактиране на кода]
Най-добре позната роля на протеините в тялото е тази на катализа на различни биохимични реакции. Ензимите са група протеини, притежаващи специфични каталитични свойства, което означава, че всеки ензим катализира една или няколко подобни реакции. Ензимите катализират разграждането на сложни молекули (катаболизъм) и синтеза им (анаболизъм), както и репликацията и репарацията на ДНК, синтеза на РНК. Има няколко хиляди ензими и около 4000 реакции, катализирани от протеини. [21] Ускоряване на реакция в резултат на ензимна катализа понякога е огромна: например, реакция катализирана от ензима оротат-карбоксилаза, се осъществява 1017 пъти по-бързо от некатализираната (78 милиона години без ензим, 18 милисекунди с ензим) [22]. Молекулите, които се свързват към ензима и се променят в ход на реакцията се наричат субстрати.
Въпреки че, ензимите обикновено са съставени от стотици аминокиселини, само една малка част от тях си взаимодейства със субстрата – средно 3 – 4 аминокиселини, често разположени далеч една от друга в основната последователност на аминокиселините – са пряко ангажирани в катализата [23]. Частта от ензима, в която се осъществява каталитичния акт се нарича активен център на ензима.
Структурна функция
[редактиране | редактиране на кода]
Структурни протеини са тези на цитоскелета, които като вид арматура придават форма на клетките и органелите и са отговорни за промените на формата на клетка. Повечето структурни протеини са фибриларни, така например мономерите на актина и туболина са глобуларни, разтворими протеини, но след полимеризация, образуват дълги нишки, които формират цитоскелета, което позволява на клетките да поддържат форма [24]. Структурни протеини са и колагена и еластина – основните компоненти на междуклетъчното вещество на съединителната тъкан (напр. хрущял) и кератина участващ в състава на коса, нокти, пера, а някои черупки.
Защитна функция
[редактиране | редактиране на кода]
Има няколко типа защитни функции на протеини:
Физическа защита. Например колаген – протеин, който е в основата на междуклетъчното вещество на съединителната тъкан (включително кости, хрущяли, сухожилията и дълбоките слоеве на кожата (дермата)); кератин, които стои в основата на роговия слой, косми, пера, рога, и други производни на епидермиса. Обикновено, тези протеини се разглеждат като протеини със структурна функция. Други примери в тази група са протеините фибриноген и тромбин [25], участващи в кръвосъсирването.
Химическа защита. Свързването на протеини с токсини може да доведе до детоксикация на последните. Особено важна роля за детоксикация в хората имат чернодробните ензими, които трансформират отровите или да ги превръщат в разтворима форма, което допринася за бързото им отделяне от тялото. [26] Имунната защита. Протеините на кръвта и другите телесни течности са включени в защитна реакция на организма от повреди или от атака на патогенните микроорганизми. Протеините на системата на комплемента и антителата (имуноглобулини) неутрализират бактерии, вируси или чужди протеини. Антителата, принадлежащи към адаптивни имунната система, се свързват към чужди за организма вещества, антигени и по този начин ги неутрализират, синтезират се от специализирани B-клетки наречени плазмоцити. [27] Докато ензимите са ограничени от афинитета към субстрата, то антитяло може да се синтезира към почти всяка молекула или молекулен комплекс. [28].
Регулаторна функция
[редактиране | редактиране на кода]Много процеси в клетките, се регулират от протеинови молекули. Тези протеини регулират транскрипцията, транслациятата, сплайсинга и дейността на другите протеини. Белтъците осъществяват регулаторната си функция чрез ензимна активност (напр. протеинкинази) или чрез специфично свързване с други молекули, обикновено засягащи взаимодействието на тези молекули с ензими.
По този начин, генната транскрипция се регулира чрез добавяне на транскрипция фактори – протеини активатори или протеини супресори – към регулаторните последователности на гените. На нивото на иРНК четенето също се регулира чрез добавяне на протеинови фактори [29], а разграждане на РНК и протеините също се осъществява от специализирани протеинови комплекси [30]. Решаваща роля в регулацията на вътреклетъчните процеси играят протеинкиназите – ензими, които активират или потискат активността на други протеини чрез свързване на фосфатни групи към тях.
Сигнална функция
[редактиране | редактиране на кода]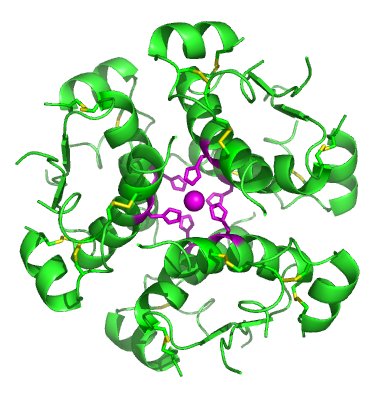
Протеините служат като сигнализиращи вещества, пренасящи сигнали между клетките, тъканите, органите и различните организми. Често сигналната функция е комбинирана с регулаторна, тъй като много вътреклетъчни регулаторните протеини също така участват в сигналната трансдукция.
Сигнална функция се извършва чрез белтъци – хормони, цитокини, растежни фактори и т.н.
Хормоните са пренасяни от кръвта. Повечето от хормоните на животните – са протеини или пептиди. Свързването на хормона към рецептора е сигнал, който води до отговор на клетката, промяна в клетъчния метаболизъм или променен профил на генна експресия. Хормоните регулират концентрацията на вещества в кръвта и клетките, растежа, възпроизводството и други процеси. Примери за такива протеини е на инсулина, който регулира нивата на кръвната захар.
Клетките взаимодействат една с друга с помощта на сигнални протеини преминаващи през междуклетъчно вещество. Тези протеини включват цитокини и растежни фактори.
Цитокините са малки пептидни молекули, служещи за комуникация между клетките. Те регулират взаимодействието между клетките, определят тяхното оцеляване, стимулират или потискат растежа, диференциацията, функционална активност и апоптозата; осигуряват съгласуваност в имунната, ендокринната и нервната система. За пример може да послужи цитокина тумор-некротизиращ фактор, който предава сигнали между клетките в тялото при възпаление и може да активира апоптоза. [31]
Транспортна функция
[редактиране | редактиране на кода]
Разтворимите протеини, участват в транспорта на малки молекули, към които имат висок афинитет и свързват, когато се намират във висока концентрация, както и лесно освобождават в места с ниска концентрация. Пример за транспортните протеини е хемоглобина, който пренася кислорода от белите дробове до други тъкани и въглероден диоксид от тъканите към белите дробове. Съществуват редица хомоложни протеини пренасящи кислород, които присъства във всички еукариотни организми. [32]
Транспортна функция осъществяват и някои мембранни протеини, които участват в транспорта на малки молекули през клетъчната мембрана, чрез промяната в пропускливостта ѝ. Липидния компонент на мембраната е водоустойчив (хидрофобен), което предотвратява преминаването на полярните и заредени молекули и йони. Мембранни транспортни белтъци биват белтъчни-канали и белтъци-преносители. Протеиновите канали съдържат вътрешни водни пори, които позволяват преминаването на йони (чрез йонни канали) или на водните молекули (чрез протеини аквапорини) през мембраната. Много йонни каналчета са специализирани в превоз на само един вид йони, например калиевите и натриевите канали правят разлика между тези подобни йони и позволяват преминаването на само един от тях. [33] Протеините преносители се свързват, подобно на ензимите, за една определена молекула или йон и за разлика от каналите могат да извършват активен транспорт, като използват енергията от хидролизата АТФ. Такъв протеин е калиево-натриевата помпа. „Електроценталата на клетката“ – АТФ синтазата, която извършва синтеза на АТФ задвижвана от протонния градиент от двете страни на вътрешната митохондриална мембрана също могат да бъде отнесена към протеините за мембранен транспорт. [34]
Рецепторна функция
[редактиране | редактиране на кода]
Протеините рецептори могат да бъдат едновременно в цитоплазмата и/или включени в клетъчната мембрана. Една част от рецептора молекула получава сигнал, който често може да е химично вещество, а в някои случаи – светлина, механични ефекти (напр., разтягане) и други стимули. Под влияние на сигнала в определена част на молекулата (протеинов рецептор) нейната конформация се изменя. В резултат на това изменение друга част на молекула предава сигнала към други клетъчни компоненти. Има няколко механизма на предаване на сигнала. Някои рецептори катализира химична реакция, докато други са йонни канали, които под влияние на сигнала се отварят или затварят, третата група свързват специален вътреклетъчен молекулен медиатор (посредник). При мембранните рецептори част от молекулата, които се свързват сигнала се намира на повърхността на клетката, а домена, който изпраща сигнал – в клетката. [35]
Двигателна функция
[редактиране | редактиране на кода]
Класа на моторните протеини осигурява движение на тялото, като мускулно съкращение, (миозин), движението на клетките в организма (например, амебовидното движение на левкоцитите), движението на ресничките и камщичетата, както и активния вътреклетъчнен транспорт (кинезин, динеин). Кинезин и динеин осъществяват превоз на молекулите по микротубулите използвайки АТФ хидролиза като източник на енергия. Динеинът придвижва молекули и органели от периферните части на клетката към центрозомата, а кинезина – в обратна посока. [36][37] Динеина е отговорен и за движението на ресничките и камшичетата на еукариотите. Цитоплазматични вариантина миозина могат да участват в транспортирането на молекули и органели чрез микрофиламентите.
Резервна функция
[редактиране | редактиране на кода]Специализираните в резервна функция белтъци се означават като резерви протеини. Те се синтезират като източник на енергия и „строителен материал“ в растителните семена и яйцата при животните. Такива са протеините от белтъка на яйцата (овалбумин) и основният млечен протеин (казеин). Няколко други протеини се използвани в тялото като източник на аминокиселини, които от своя страна са предшественици на биологично активни вещества, които регулират обмяната на веществата. Рядко, при продължително гладуване и след изчерпване на въглехидратните и липидните запаси, белтъците започват да се разграждат, като енергията от тях се използва за поддържане на жизнените функции.
Значение
[редактиране | редактиране на кода]Белтъците участват в голяма част от процесите, протичащи в живия организъм. Структурата на клетката, протичането на метаболитните процеси, регулацията на хомеостазата и дори защитата на организма се дължи преди всичко на конкретни белтъци. Ролята им в съхраняването и реализирането на генетичната информация, ги нарежда сред най-важните компоненти на живата клетка, наред с нуклеиновите киселини, въглехидратите и липидите.
Представители
[редактиране | редактиране на кода]Източници
[редактиране | редактиране на кода]- ↑ Oxford Dictionary of English[неработеща препратка]
- ↑ Nature journal
- ↑ THE ORIGIN OF THE WORD PROTEIN
- ↑ Nelson DL, Cox MM. Lehninger's Principles of Biochemistry. 4th. New York, New York, W. H. Freeman and Company, 2005.
- ↑ Gutteridge A, Thornton JM. Understanding nature's catalytic toolkit // Trends in Biochemical Sciences 30 (11). 2005. DOI:10.1016/j.tibs.2005.09.006. с. 622 – 29.
- ↑ а б Murray RF, Harper HW, Granner DK, Mayes PA, Rodwell VW. Harper's Illustrated Biochemistry. New York, Lange Medical Books/McGraw-Hill, 2006. ISBN 0-07-146197-3.
- ↑ а б Lodish H, Berk A, Matsudaira P, Kaiser CA, Krieger M, Scott MP, Zipurksy SL, Darnell J. Molecular Cell Biology. 5th. New York, New York, WH Freeman and Company, 2004.
- ↑ а б Van Holde KE, Mathews CK. Biochemistry. Menlo Park, California, Benjamin/Cummings Pub. Co., Inc, 1996. ISBN 0-8053-3931-0.
- ↑ Dobson CM. The nature and significance of protein folding // Mechanisms of Protein Folding. Oxford, Oxfordshire, Oxford University Press, 2000. ISBN 0-19-963789-X. с. 1 – 28.
- ↑ Fulton A, Isaacs W. Titin, a huge, elastic sarcomeric protein with a probable role in morphogenesis // Bioessays 13 (4). 1991. DOI:10.1002/bies.950130403. с. 157 – 61.
- ↑ Bruckdorfer T, Marder O, Albericio F. From production of peptides in milligram amounts for research to multi-tons quantities for drugs of the future // Current Pharmaceutical Biotechnology 5 (1). 2004. DOI:10.2174/1389201043489620. с. 29 – 43.
- ↑ Schwarzer D, Cole P. Protein semisynthesis and expressed protein ligation: chasing a protein's tail // Current Opinions in Chemical Biology 9 (6). 2005. DOI:10.1016/j.cbpa.2005.09.018. с. 561 – 69.
- ↑ Kent SB. Total chemical synthesis of proteins // Chemical Society Reviews 38 (2). 2009. DOI:10.1039/b700141j. с. 338 – 51.
- ↑ Demartino GN, Gillette TG. Proteasomes: machines for all reasons. Cell. 2007 May 18. 129(4):659 – 662
- ↑ Bronner C, Chataigneau T, Schini-Kerth VB, Landry Y. The „Epigenetic Code Replication Machinery“, ECREM: a promising drugable target of the epigenetic cell memory. Curr Med Chem. 2007;14(25):2629 – 2641
- ↑ а б в г д е ж з и к л Ангелов А., Гачев Е., Николов Т. Биохимия за медици и стоматолози. София, Университетско издателство „Св. Климент Охридски“, 1996.
- ↑ Anfinsen C. Principles that Govern the Folding of Protein Chains // Science 181. 1973. с. 223 – 229.
- ↑ Ellis RJ, van der Vies SM. Molecular chaperones // Annu. Rev. Biochem. 60. 1991. DOI:10.1146/annurev.bi.60.070191.001541. с. 321 – 347.
- ↑ Sun Y, MacRae TH. The small heat shock proteins and their role in human disease // FEBS J. 60. 2005. с. 2613 – 2627.
- ↑ Yannay-Cohen N, Razin E. Translation and transcription: the dual functionality of LysRS in mast cells // Mol Cells. 22. 2000. PMID 17085962. с. 127 – 132. Архивиран от оригинала на 2008-06-18.
- ↑ Bairoch A. The ENZYME database in 2000 // Nucleic Acids Res 28. 2000. PMID 10592255. с. 304 – 305.
- ↑ Radzicka A, Wolfenden R. A proficient enzyme // Science 6 (267). 1995. PMID 7809611. с. 90 – 93.
- ↑ The Catalytic Site Atlas at The European Bioinformatics Institute
- ↑ Erickson HP. Evolution of the cytoskeleton. Bioessays. 2007:668 – 677
- ↑ Wolberg AS (2007). „Thrombin generation and fibrin clot structure“. Blood Rev. 21(3): 131 – 142. PMID 17208341.
- ↑ Я. Кольман, К.-Г. Рем. Наглядная биохимия. М.: Мир, 2000, с. 308 – 309.
- ↑ J. Li, D. R. Barreda, Y.-A. Zhang, H. Boshra, A. E. Gelman, S. LaPatra, L. Tort & J. O. Sunyer (2006). „B lymphocytes from early vertebrates have potent phagocytic and microbicidal abilities“. Nature Immunology 7: 1116 – 1124. PMID 16980980.
- ↑ Felix NJ, Allen PM. Specificity of T-cell alloreactivity. Rev Immunol. 2007 Dec; 7(12):942 – 953
- ↑ Hinnebusch AG. Translational regulation of GCN4 and the general amino acid control of yeast. Annu Rev Microbiol. 2005;59:407 – 450
- ↑ Anderson P, Kedersha N. RNA granules. Cell Biol. 2006:172(6):803—808
- ↑ Повещенко А. Ф., Абрамов В. В., Козлов В. В. Цитокины – факторы нейроэндокринной регуляции. Успехи физиологических наук. 2007 – 38(3):40 – 46
- ↑ Wittenberg JB. On optima: the case of myoglobin-facilitated oxygen diffusion. Gene. 2007 Aug 15. 398(1 – 2):156 – 161.
- ↑ Driessen AJ, Nouwen N. Protein Translocation Across the Bacterial Cytoplasmic Membrane. Annu Rev Biochem. 2007 Dec 13 [Epub ahead of print]
- ↑ Drory O, Nelson N. The emerging structure of vacuolar ATPases // Physiology (Bethesda). 21. 2006. PMID 16990452. с. 317 – 325. Архивиран от оригинала на 2008-11-21.
- ↑ Dupré DJ, Hébert TE. Biosynthesis and trafficking of seven transmembrane receptor signalling complexes. Cell Signal. 2006;18(10):1549 – 1559
- ↑ Karp G. Cell and Molecular Biology: Concepts and Experiments, Fourth ed, pp. 346 – 358. John Wiley and Sons, Hoboken, NJ. 2005.
- ↑ Schroer, Trina A. Dynactin. Annual Review of Cell and Developmental Biology. 2004 20, 759 – 779. PMID 15473859