Xantofyll
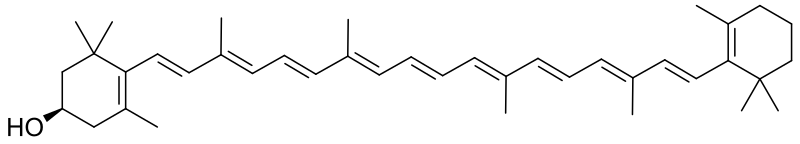


Xantofyller är gula växtpigment som är en av de två huvudgrupperna av karotenoider (tetraterpenoider med 40 kolatomer). Namnet, som kommer från grekiska ξανθός xanthos ("gul") och φύλλον phyllon ("blad"), har de fått på grund av det gula band de bildar vid kromatografi på ett extrakt av bladpigment. Deras molekylstruktur liknar karotenerna, som är den andra huvudgruppen av karotenoider, men xantofyllerna innehåller syre till skillnad från karotenerna som är rena kolväten. Xantofyllernas syre förekommer som en hydroxylgrupp och/eller som en syrebrygga (epoxid) som substituerar två väteatomer. På grund av dessa syreatomer är de mera polära än karotenerna, och det är denna skillnad som tillåter separation genom många typer av kromatografi. Karotener är i allmänhet mera orange än de vanligen gulare xantofyllerna.
Liksom andra karotenoider förekommer xantofyllerna talrikast i bladen hos de flesta gröna växter där de modererar den mängd ljusenergi som är tillgänglig för fotosyntes och hanterar (genom "icke-fotokemisk energiupptagning") tripletter (en excitationsform) av klorofyll[1][2] som överproduceras i starkt ljus. De xantofyller som finns i djurens kroppar och i djurprodukter kommer ursprungligen från dieten och den gula färgen hos äggula, fett och skinn kommer från xantofyller i födan (huvudsakligen lutein, som ofta tillförs hönsfoder för att ge gulan kraftigare färg).
Färgen hos den gula fläcken i ögats näthinna kommer från det lutein och zeaxantin den innehåller, och båda dessa xantofyller har alltså kommit via födan. Dessa skyddar ögat genom att absorbera joniserande blått ljus.[3] Dessa xantofyller är inte verksamma i synprocessen, eftersom de inte kan omvandlas till retinal (vitamin A-aldehyd) som karotenoiderna.
Xantofyllerna innefattar bland annat lutein, zeaxantin, anteraxantin, violaxantin, neoxantin, fukoxantin samt α- och β-kryptoxantin. Den senare är den enda kända xantofyllen som innehåller en β-jononring och ger därför en vitamin A om den klyvs på mitten. Det är därmed den enda xantofyllen som fungerar som provitamin A hos däggdjur.[4] Det fungerar dock endast som provitamin för växtätare som har ett enzym för att omvandla karotenoider som innehåller β-jonon till retinal. Hos andra djur än däggdjur kan vissa xantofyller omvandlas till hydroxylerade retinal-analoger som fungerar direkt som synpigment. Exempelvis använder många insekter, med undantag från bland andra några flugor och fjärilar, den från xantofyll deriverade R-isomeren av 3-hydroxiretinal som synpigment, vilket innebär att β-kryptoxantin och andra xantofyller (som lutein och zeaxantin) kan fungera som ett "A-vitamin" för dem, medan karotener (som β-karoten) inte gör det.[5]
Xantofyllcykeln
[redigera | redigera wikitext]
Genom att skapa eller avlägsna epoxidbryggor kan man bilda andra xantofyller av befintliga. Dessa processer kallas xantofyllcykler.[6] Den viktigaste av dessa cykler, och den man normalt avser med "xantofyllcykeln", är den som går från violaxantin (två epoxidbryggor) via anteraxantin (en brygga) till zeaxantin (inga bryggor) och åter, och som ofta kallas "VAZ" efter initialerna i de tre ämnena eller violaxantincykeln för att skilja den från de andra.
En annan sådan cykel finns hos kromalveolater (som brunalger, kiselalger, dinoflagellater och fästalger) och innefattar diadinoxantin (en epoxidbrygga) som kan omvandlas till diatoxantin (ingen brygga).[7][8]
Den tredje xantofyllcykeln innefattar de-epoxidering av luteinepoxid till lutein och den omvända epoxideringen.[9]
Dessa tre cykler spelar en nyckelroll för antennpigmentens så kallade "icke-fotokemiska energiupptagning" (non-photochemical quenching, NPQ), det vill säga ljusabsorption som ombildas till värme i stället för att ledas via klorofyll till fotosyntesens reaktionscentra, vilket skyddar klorofyllmolekylerna vid hög ljusintensitet och är ett av de viktigaste sätten att förhindra fotoinhibering.[10] Vid hög ljusintensitet bildas, med hjälp av enzymet violaxantin-de-epeoxidas (VDE), zeaxantin från violaxantin via anteroxantin. Violaxantin fungerar som ett antennpigment och kan således leverera uppfångad ljusenergi vidare till fotosyntesen, medan zeaxantin kan ta emot energi från klorofyll och omvandla det till värme.[6] När ljuset minskas går reaktionen (långsammare) åt motsatta hållet med hjälp av zeaxantin-epoxidas (ZE).[11][12] I de två andra cyklerna spelar diadinoxantin och luteinepoxid rollen som ljussamlande antennpigment, medan diatoxantin och lutein spelar samma skyddande roll som zeaxantin.[9]
Biosyntes
[redigera | redigera wikitext]Levande organismer syntetiserar xantofyller genom oxidation av karotener. Lutein har α-karoten som prekursor, medan de flesta övriga xantofyllerna syntetiseras från β-karoten, och prekursorn till båda dessa karotener är lykopen (som i sin tur fås från två geranylgeranylpyrofosfatmolekyler via fytoen).
Från β-karoten får man kantaxantin (via echienon) eller zeaxantin (via β-kryptoxantin) och från dessa båda kan man få astaxantin via antingen fenikoxantin (från kantaxantin) eller adonixantin (från zeaxantin). Från zeaxantin får man, som nämnts under xantofyllcykeln ovan, violaxantin via anteraxantin och från violaxantin kan troligen neoxantin och diadinoxantin syntetiseras. Dessa båda senare kan troligen ombildas till fukoxantin och, som ovan nämnts under xantofyllcykeln, kan diadinoxantin i sin tur ombildas till diatoxantin.[13]
Från lutein kan man erhålla loroxantin[14] och , som nämnts under xantofyllcykeln, luteinepoxid.
Xantofyllkällor i födan
[redigera | redigera wikitext]Xantofyller som lutein, zeaxantin och kantaxantin förekommer huvudsakligen i gröna blad och gröna, gula och orange frukter.[15] De två xantofyller som är viktiga för människans (och andra vertebraters) syn, zeaxantin och lutein, måste tillföras genom födan. Zeaxantin finns i rik mängd i majs, orange paprika, salladslök och äggula medan lutein finns rikligt i bladgrönsaker (särskilt spenat, grönkål, romansallad, koriander och persilja - som alla också innehåller mycket β-karoten, d.v.s. provitamin A).[16] I näthinnan förekommer huvudsakligen trans-xantofyller som förekommer rikligast i obehandlade grönsaker. Tillagade eller på annat sätt behandlade (som exempelvis "frukostflingor") födoämnen innehåller mer av cis-isomerer, troligen på grund av isomeriseringsprocesser under behandlingen.[16]
Referenser
[redigera | redigera wikitext]- Demmig-Adams, B & W. W. Adams, 2006. Photoprotection in an ecological context: the remarkable complexity of thermal energy dissipation, New Phytologist, 172: 11–21.
- ^ Ballottari, Matteo; Mozzo, Milena; Girardon, Julien. ”Chlorophyll Triplet Quenching and Photoprotection in the Higher Plant Monomeric Antenna Protein Lhcb5”. The Journal of Physical Chemistry B 117 (38): sid. 11337-11348. doi:. ISSN 1520-6106. http://dx.doi.org/10.1021/jp402977y. Läst 20 juli 2015.
- ^ Linda Holmer, 2013, Solning på låg höjd - fotosyntesen hos grön svavelbakterie. Arkiverad 21 december 2014 hämtat från the Wayback Machine., sid. 2.
- ^ Jane Higdon, Victoria J. Drake, Elizabeth J. Johnson, 2009, Carotenoids på Linus Pauling Institute Micronutrient Information Centre, Oregon State University.
- ^ Liu, Y.-Q.; Davis, C. R.; Schmaelzle, S. T.. ”β-Cryptoxanthin biofortified maize (Zea mays) increases β-cryptoxanthin concentration and enhances the color of chicken egg yolk” (på engelska). Poultry Science 91 (2): sid. 432-438. doi:. ISSN 0032-5791. http://ps.oxfordjournals.org/content/91/2/432. Läst 20 juli 2015.
- ^ Klaus Vogt, 1984, The Chromophore of the Visual Pigment in Som e Insect Orders, Zeitschrift für Naturforschung C - A Journal of Biosciences, 39 c, 196-197 (1984).
- ^ [a b] Xantofyllcykeln i Nationalencyklopedin.
- ^ Lohr, Martin; Wilhelm, Christian. ”Algae displaying the diadinoxanthin cycle also possess the violaxanthin cycle” (på engelska). Proceedings of the National Academy of Sciences 96 (15): sid. 8784-8789. doi:. ISSN 0027-8424. http://www.pnas.org/content/96/15/8784. Läst 20 juli 2015.
- ^ J.D. Hedley, P.J. Mumby, Remote sensing of Pigmentation in Coral Reef Organisms i Alan J. Southward, P.A. Tyler, Craig M. Young, Lee A. Fuiman, 2002, Advances in Marine Biology, Volym 43, ISBN 0-12-026143-X, sid.285.
- ^ [a b] Martin Lohr, Carotenoid Metabolism in Phytoplankton i Suzanne Roy, Carole A. Llewellyn, Einar Skarstad Egeland, Geir Johnsen (eds.), 2011, Phytoplankton Pigments: Characterization, Chemotaxonomy and Applications in Oceanography, ISBN 978-1-107-00066-7, sid. 68.
- ^ Bilger, Wolfgang; Björkman, Olle. ”Role of the xanthophyll cycle in photoprotection elucidated by measurements of light-induced absorbance changes, fluorescence and photosynthesis in leaves of Hedera canariensis” (på engelska). Photosynthesis Research 25 (3): sid. 173-185. doi:. ISSN 0166-8595. https://link.springer.com/article/10.1007/BF00033159. Läst 20 juli 2015.
- ^ Marie Eskling, Anna Emanuelsson, Hans-Erik Åkerlund, 2001, Enzymes and mechanisms for violaxanthin-zeaxanthin conversion, i Advances in Photosynthesis and Respiration vol. 11, 2001, sid. 433-452.
- ^ Eskling M, Arvidsson P och Åkerlund H, 1997. The xanthophyll cycle, its regulation and components. Physiologia Plantarum 100, 806-816
- ^ Schema över syntesvägarna för astaxantin, lutein och fukoxantin från lykopen. i Chonglong Wang, Jung-Hun Kim, Seon-Won Kim, 2014, Synthetic Biology and Metabolic Engineering for Marine Carotenoids: New Opportunities and Future Prospects i Marine Drugs, Sep 2014; 12(9): 4810–4832.
- ^ Eonseon Jin, Juergen Polle et. al, 2003,Xanthophylls in Microalgae: From Biosynthesis to Biotechnological Mass Production and Application i J. Microbiol. Biotechnol. (2003), 13(2), 165–174. Denna artikel ger också strukturformler för de ovannämnda föreningarna.
- ^ Zaripheh, Susan; Erdman, John W.. ”Factors That Influence the Bioavailablity of Xanthophylls” (på engelska). The Journal of Nutrition 132 (3): sid. 531S-534S. ISSN 0022-3166. http://jn.nutrition.org/content/132/3/531S. Läst 20 juli 2015.
- ^ [a b] Perry, Alisa; Rasmussen, Helen; Johnson, Elizabeth J.. ”Xanthophyll (lutein, zeaxanthin) content in fruits, vegetables and corn and egg products”. Journal of Food Composition and Analysis 22 (1): sid. 9-15. doi:. http://www.sciencedirect.com/science/article/pii/S0889157508001336. Läst 20 juli 2015.