Elasmozaur
| Elasmosaurus | |
| Cope, 1868 | |
| Okres istnienia: 80,5 mln lat temu | |
 Elasmosaurus platyurus | |
| Systematyka | |
| Domena | |
|---|---|
| Królestwo | |
| Typ | |
| Podtyp | |
| Gromada | |
| Nadrząd | |
| Rząd | |
| Podrząd | |
| Rodzina | |
| Rodzaj |
Elasmosaurus |
Elasmozaur (Elasmosaurus) – rodzaj plezjozaura żyjącego w Ameryce Północnej w kampanie (kreda późna), około 80,5 miliona lat temu. Pierwszy okaz odkryto w 1867 w Kansas blisko Fort Wallace, wysłano go do amerykańskiego paleontologa Cope’a, który na jego podstawie wyróżnił nowy rodzaj i gatunek, ten ostatni nazwał E. platyurus w 1868. Nazwa rodzajowa oznacza „cienkopłytowy jaszczur”, natomiast epitet gatunkowy znaczy „płaskoogonowy”. Cope pierwotnie zrekonstruował szkielet zwierzęcia, umieszczając głowę na ogonie, który to błąd nagłośnił paleontolog Marsh, rywal Cope’a w wojnie o kości. Odkryto tylko jeden niekompletny szkielet elasmozaura. Składa się on z niepełnej czaszki, kręgosłupa, obręczy kończyn piersiowej i miednicznej. Obecnie wyróżnia się pojedynczy gatunek, inne zaliczane dawniej do omawianego rodzaju uznaje się za niepewne bądź też przeniesiono je do innych rodzajów.
Elasmozaur miał 10,3 m długości. Cechowało go opływowe ciało o wiosłowatych kończynach, krótki ogon, niewielka głowa na końcu niezwykle długiej szyi, osiągającej 7,1 m długości. Wraz ze swym krewnym Albertonectes należał do najbardziej długoszyich zwierząt, jakie kiedykolwiek żyły, miał też najwięcej kręgów szyjnych: 72. Czaszka była smukła, trójkątna, na przedzie miała duże, przypominające kły zęby, z tyłu zęby były mniejsze. Każda kość przedszczękowa miała przypuszczalnie po 6 zębów, kość szczękowa 14, natomiast kość zębowa – 19. Większość kręgów szyjnych cechowało spłaszczenie boczne, a wzdłuż boków przebiegał podłużny grzebień bądź kil.
Od nazwy elasmozaura wywodzi się nazwa rodziny elasmozaurów. Elasmozaur był pierwszym rozpoznanym członkiem tej grupy długoszyich plezjozaurów. Członkowie wspomnianej rodziny dobrze zaadaptowali się do wodnego trybu życia. Pływali za pomocą płetw. Wbrew wcześniejszym wyobrażeniom nie mieli zbyt giętkiej szyi, nie potrafili wystawiać jej do góry ponad powierzchnię wody. Nie wiadomo do czego służyły im tak długie szyje – być może spełniały jakąś funkcję związaną ze zdobywaniem pokarmu. Elasmozaur prawdopodobnie żywił się niewielkimi rybami i morskimi bezkręgowcami, chwytając je długimi zębami. Przedstawiciele rodziny mogli wykorzystywać gastrolity, pomagające im w trawieniu. Elasmosaurus pochodzi z formacji Pierre Shale, reprezentującej osady morskie Morza Środkowego Zachodu.
Budowa
[edytuj | edytuj kod]
Jedyny znany okaz elasmozaura (ANSP 10081) jest fragmentaryczny, brakuje weń wielu elementów. Spokrewnione z nim elasmozaury wskazują, że powinien on mieć zwarte, opływowe ciało, płetwowate kończyny, krótki ogon, proporcjonalnie małą głowę i niezwykle długą szyję. Jej długość szacuje się na 7,1 m[1]. Zalicza to elasmozaura i jego bliskiego krewnego, rodzaj Albertonectes, do zwierząt o najdłuższej szyi, jakie kiedykolwiek żyły oraz o największej liczbie kręgów szyjnych wśród wszystkich kręgowców[2][3]. Jednak pomimo tak licznych kręgów szyjnych szyje elasmozaurów nie były nawet w połowie tak długie, jak szyje zauropodów[1]. W 1869 amerykański paleontolog Edward Drinker Cope oszacował długość kompletnego szkieletu zwierzęcia przez zsumowanie długości kręgów szyjnych i szacunki brakujących części, na 13,1 m. Za życia zwierzę byłoby jeszcze nieco dłuższe z powodu chrząstek pomiędzy trzonami kręgów i wedle szacunków Cope’a mierzyło 13,7 m[4]. W 1952 amerykański paleontolog Samuel Welles oszacował długość ciała tego zwierzęcia na 10,3 m[5].

Jak inne elasmozaury, Elasmosaurus miał smukłą, trójkątną czaszkę. Pysk był zaokrąglony, widziany z góry prawie tworzył półokrąg. Kości przedszczękowe miały pośrodkowo niski kil. Z powodu niekompletności zachowanych szczątków nie jest jasna liczba zębów tego zwierzęcia. Każda kość przedszczękowa miała prawdopodobnie 6 zębów, które przypominały duże kły. Liczba ta odróżniała elasmozaura od prymitywnych Plesiosauroidea i większości innych elasmozaurów, które na analogicznych kościach miały mniej zębów. Dwa zęby na przedzie ustępowały rozmiarami kolejnym, umiejscowione były pomiędzy pierwszymi dwoma zębami kości zębowych. Znane zęby przedniej części żuchwy przypominały duże kły. Elasmozaury cechowało uzębienie heterodontyczne, nieregularne. Zęby od przodu ku tyłowi stawały się coraz mniejsze. Kości szczękowe miały 14 zębów, zaś kości zębowe miały ich od 17 do 19. Zęby zachodziły na siebie. Ich korony były smukłe, zaokrąglone w przekroju. Spojenie żuchwy, gdzie łączyły się obie jej połowy, było mocno skostniałe, o niedostrzegalnym szwie[2][6].
Obręcze kończyn piersiowych i miednicznych holotypu w 1906 uznano za zaginione, jednak znane są z oryginalnych opisów i ilustracji z końca XIX wieku. Łopatki były połączone, stykając się w płaszczyźnie pośrodkowej, jednak bez śladu pośrodkowej belki. Miały szerokie wyrostki oraz długie szyjki. Z kolei obręcz miedniczna nosiła pośrodkowo belkę, była to prawdopodobnie zaawansowana ewolucyjnie cecha, nieobecna u młodocianych plezjozaurów. Kości kulszowe również łączyły się w płaszczyźnie pośrodkowej, wobec czego wspomniana wyżej pośrodkowa belka przebiegała wzdłuż długości miednicy. Cechy takiej zazwyczaj nie spotyka się u plezjozaurów[6]. Jak i inne elasmozaury (i plezjozaury w ogólności), Elasmosaurus powinien mieć duże, wiosłowate kończyny zakończone bardzo długimi palcami. Płetwy przednie (piersiowe) były dłuższe od tylnych (miednicznych)[2].
Kręgi
[edytuj | edytuj kod]
W przeciwieństwie do wielu innych zwierząt o długiej szyi poszczególne kręgi szyjne elasmozaura nie były szczególnie wydłużone. Niezwykłą długość szyja osiągała raczej dzięki zwiększonej ich liczbie[1]. Elasmosaurus wyróżniał się wśród plezjozaurów 72 kręgami szyjnymi. Niewykluczone, że było ich jeszcze więcej, ale część uległa erozji bądź zniszczeniu po wydobyciu. Tylko Albertonectes posiadał większą liczbą kręgów szyjnych: 75. Są to jedyne dwa rodzaje plezjozaurów mające powyżej 70 kręgów szyjnych, przy czym już obecność powyżej 60 stanowi cechę ewolucyjnie zaawansowaną u tej grupy[3][6].
Zespół dźwigacza i obrotnika, a więc dwóch pierwszych kręgów szyjnych, połączony stawowo z tylną częścią czaszki, był długi, niski i horyzontalnie prostokątny w widoku z boku. Trzony tych kręgów łączył u holotypowego okazu kościozrost, co wskazuje, że było to dorosłe zwierzę. Łuki kręgowe były bardzo cienkie i raczej wysokie, tworzyły one przebiegający w środku kanał kręgowy o trójkątnym, patrząc z tyłu, obrysie. Dolna część kanału kręgowego była wąska ku tyłowi obrotnika, osiągając tam połowę szerokości trzonu. Ku przodowi robił się szerszy, prawie osiągając szerokość trzonu dźwigacza. Jego łuki są także potężniejsze niż u obrotnika. Kanał kręgowy jest wyższy. Wyrostek kolczysty był niski, kierował się do góry i do tyłu. Trzony atlasu i obrotnika mają wspólne cechy: podobną długość i kwadratowy kształt widziany z boku. Powierzchnie stawowe dla kolejnego kręgu są owalne w obrysie, na górnym brzegu pośrodku leży zagłębienie dla kanału kręgowego. Wydatny kil biegnie dołem pośrodkowo wzdłuż dźwigacza i obrotnika[6].

Większość kręgów szyjnych była spłaszczona bocznie, zwłaszcza pośrodku szyi. Grzebień czy też kil biegł podłużnie wzdłuż boku kręgów szyjnych, co stanowi cechę typową dla Elasmosauridae. Widać to od trzeciego do pięćdziesiątego piątego kręgu, w tylnej części szyi. Grzebień leżał pośrodku trzonu w kręgach przednich (do osiemnastego włącznie), a w następnych kręgach znajdował się w górnej połówce trzonu. Służył on jako miejsce przyczepu mięśni szyi. Trzony różniły się kształtem zależnie od pozycji w szyi. Trzon trzeciego kręgu był prawie tak długi, jak szeroki. Od czwartego kręgu trzony stawały się dłuższe niż szersze. Trzony stawały się bardziej wydłużone w środkowym odcinku szyi, a w tylnym były coraz krótsze. Ich długość zrównywała się z szerokością około sześćdziesiątego pierwszego kręgu, a najbardziej dystalne były szersze niż dłuższe. Powierzchnie stawowe kręgów z przodu szyi były szerokie, owalne i umiarkowanie wklęsłe, z zaokrąglonymi, pogrubionymi brzegami, z zagłębieniem lub jamą po górnej i dolnej stronie. Dalej w tył w proksymalnej części szyi, około dwudziestego piątego kręgu, dolny brzeg powierzchni stawowej stawał się bardziej wklęsły, a sama powierzchnia przybierała kształt kwadratowy o zaokrąglonych brzegach. Do kręgu sześćdziesiątego trzeciego powierzchnia stawowa była kwadratowa, o zaokrąglonych brzegach, podczas gdy trzony najbardziej dystalnych kręgów cechowały się szerokim, owalnym obrysem[2][3][6].
Łuki kręgowe kręgów szyjnych były dobrze umocowane do trzonów. Nie widać szwów. Kanał kręgowy był wąski w przednich kręgach, stając się wydatniejszy w tylnych, gdzie był tak szeroki, jak wysoki i prawie okrągły. Górne i dolne wyrostki stawowe kręgów szyjnych posiadały powierzchnie stawowe dla sąsiednich kręgów, pasujące do siebie, podobnej wielkości. Poprzedni osiągał całkowicie poziom trzonu, podczas gdy następny osiągał jedynie tylną połowę. Wyrostki kolczyste są niskie i od dwudziestego kręgu prawie półkoliste. Powierzchnie stawowe dla żeber szyjnych leżały na dolnych bokach trzonu, tylko na trzech ostatnich kręgach wyżej, osiągając środek boku trzonu. Żebra szyjne były w widoku bocznym półokrągłe do kwadratowych, kierowały się raczej prosto do dołu. Na dole każdego z kręgów szyjnego widniała para otworów odżywczych, przegrodzonych grzbietem kostnym, coraz wydatniejszym i grubszym u kolejnych kręgów, idąc ku tyłowi szyi[6].

Kręgi przejściowe pomiędzy szyjnymi i grzbietowymi w okolicy piersiowej plezjozaurów, blisko przedniego brzegu obręczy kończyny piersiowej, są często zwane kręgami piersiowymi (pectoral vertebrae). Elasmosaurus miał trzy takie kręgi, co stanowi częstą cechę rodziny elazmozaurów. Powierzchnie stawowe dla żeber kręgów piersiowych były trójkątnego kształtu i leżały na wyrostkach poprzecznych. Natomiast trzony miały po parze otworów odżywczych pośrodku dolnej części boku. Kręgi grzbietowe miały powierzchnie stawowe dla żeber na poziomie kanału kręgowego. Przednią i tylną część ich wyrostków poprzecznych zdobiły na brzegach wydatne grzbiety. Tutaj powierzchnie stawowe dla żeber lokowały się powyżej wyrostków poprzecznych, rozdzielając je. Były owalne do prostokątnych w obrysie. Wyrostki stawowe przednie nie dorównywały długością ich odpowiednikom odcinka szyjnego czy piersiowego, sięgały bowiem zaledwie poziomu trzonu jedną trzecią długości. Tylne z kolei osiągały więcej niż poziom trzonu tylną połowę swej długości. Kręgi grzbietowe nie nadają się do rozróżniania Elasmosauridae, gdyż nie przedstawiają cech diagnostycznych na poziomie rodzaju[3][6].
Elasmosaurus miał cztery kręgi krzyżowe, połączone razem w kość krzyżową wchodzącą w skład miednicy. Liczba ta stanowi wartość typową dla rodziny. Wyrostki poprzeczne tego odcinka kręgosłupa były bardzo krótkie, a wymiary powierzchni stawowych dla żeber rosły od pierwszego do czwartego kręgu krzyżowego. Wzdłuż szczytu tychże kręgów biegł grzbiet. Dolne boki kręgów były zaokrąglone. Widniały na nich pary otworów odżywczych, przedzielone niskimi grzbietami. Pierwszy kręg ogonowy wyróżnia się mniejszymi niż u poprzedzających go kręgów krzyżowych powierzchniami stawowymi dla żeber oraz ich ulokowaniem w dolnej połowie trzonu. Są to kręgi prawie okrągłe w kształcie. Pierwsze dwa mają wąskie kile pośrodku górnego boku. Powierzchnie stawowe żebrowe kręgów ogonowych położone były u dołu trzonów, a ich owalny kształt zwiększał swój rozmiar i robił się coraz szerszy od trzeciego kręgu, malejąc znowu od czternastego. Wyrostki stawowe przednie osiągały znowu poziom trzonów na większości długości, tylne zaś jedynie na połowie swej długości. Dolna część trzonów była zaokrąglona od pierwszego do trzeciego kręgu, wklęsła od czwartego do osiemnastego. Przedstawiciele rodziny zazwyczaj mieli 30 kręgów ogonowych[6]. Jako że ostatnie ich kręgi łączyły się w strukturę przypominającą pygostyl ptaków, istnieje możliwość, że wspierał on płetwę ogonową, jednak jej ewentualny kształt pozostaje tajemnicą[2].
Historia odkryć
[edytuj | edytuj kod]
Wiosną 1867 chirurg amerykańskiej armii Theophilus Hunt Turner i zwiadowca William Comstock eksplorowali skały w okolicy Fort Wallace w Kansas, gdzie stacjonowali podczas budowy Union Pacific Railway. Około 23 km na północny wschód od Fort Wallace, blisko McAllaster, Turner odkrył kości dużego kopalnego gada w wąwozie, w skałach formacji Pierre Shale. W czerwcu Turner dał trzy skamieniałe kręgi pochodzące z tego odkrycia amerykańskiemu naukowcowi nazwiskiem John LeConte. W grudniu LeConte dostarczył część kręgów do Cope’a do Academy of Natural Sciences of Philadelphia (ANSP, znana od 2011 jako Academy of Natural Sciences of Drexel University). Rozpoznając w nich szczątki plezjozaura, większego niż jakiekolwiek widziane dotychczas w Europie, Cope napisał do Turnera z prośbą o dostarczenie reszty okazu na koszt ANSP[6][7][8].
W grudniu 1867 Turner i współpracownicy z Fort Wallace wrócili na miejsce znaleziska i odkryli większość kręgosłupa, jak też konkrecje zawierające inne kości. Materiał ważył w sumie 360 kg. Skamieniałości zostały wydobyte ze względnie miękkiego łupka ilastego, po czym załadowano je na wóz konny i przetransportowano do Fort Wallace. Cope wysłał instrukcje, jak należy zapakować kości. Wysłano je później wagonem wojskowym w wyściełanych sianem pojemnikach. Okaz dotarł koleją do Filadelfii w marcu 1868. Cope pośpiesznie je zbadał. Mówił o nich na marcowym spotkaniu ANSP, podczas którego określał je nazwą Elasmosaurus platyurus. Nazwa rodzajowa Elasmosaurus oznacza „cienkopłytowego jaszczura”, co nawiązuje do płytowatych kości okolic mostkowej i miednicznej. Epitet gatunkowy platyurus znaczy „płaskoogonowy” i odnosi się do spłaszczonego ogona i blaszek kręgów ogonowych[4][7][9][10][11].

Cope poprosił Turnera o poszukanie innych pozostałości okazu Elasmosaurus. Więcej skamieniałości wysłano mu w sierpniu i wrześniu 1868. ANSP w grudniu 1868 podziękował Turnerowi za te okazy, a Turner odwiedził muzeum wiosną, kiedy nie było w nim Cope’a. Turner zmarł niespodziewanie w Fort Wallace 22 lipca 1869, nie widząc zakończenia rozpoczętej przez siebie pracy, jednak Cope w dalszym ciągu pisał do niego, nieświadom jego zgonu do 1870. Okoliczności odkrycia przez Turnera okazu typowego nie zostały uwzględnione w opisie Cope’a i pozostawały nieznane aż do publikacji listów Turnera w 1987. Elasmosaurus był pierwszą większą skamieniałością znalezioną w Kansas (i największą, jaką kiedykolwiek znaleziono w tym stanie). Jego odkrycie zapoczątkowało na tym terenie intensywne poszukiwania przez zbieraczy, w efekcie czego tysiące skamieniałości z tego stanu wysłano do czołowych muzeów amerykańskiego wschodniego wybrzeża[7]. Elasmosaurus był jednym z kilku plezjozaurów znanych z Nowego Świata w tych czasach i pierwszym znanym członkiem długoszyjej rodziny Elasmosauridae[6].

W 1869 Cope opisał naukowo i narysował kości elasmozaura. Preprint zawierał rekonstrukcję szkieletu, którą badacz wcześniej zaprezentował na spotkaniu ANSP we wrześniu 1868. Przedstawiała ona Elasmosaurus z krótką szyją i długim ogonem, inaczej, niż inne plezjozaury. Autor ów nie miał też pewności, gdzie należało umieścić kończyny tylne. Półtora roku później na spotkaniu ANSP (marzec 1870) amerykański paleontolog Joseph Leidy, mentor Cope’a, zauważył, że w owej rekonstrukcji czaszkę umieszczono na złym końcu kręgosłupa elasmozaura: zamiast na szyi zamontowana została na końcu ogona. Cope widocznie przyjął, że kręgi, przy których znaleziono szczękę i żuchwę muszą reprezentować odcinek szyjny, nie zauważając, iż obrotnik i atlas są na badanym okazie na drugim końcu kręgosłupa i nie biorąc pod uwagę, że szczęki mogły być przemieszczone pośmiertnie. Leidy uznał także elasmozaura za identycznego z Discosaurus, którego to plezjozaura odkrył i opisał w 1851[4][7][12][13].

Chcąc ukryć pomyłkę, Cope próbował wycofać wszystkie egzemplarze preprintu i w 1870 wydrukował poprawioną wersję z nową rekonstrukcją szkieletu, w której głowę umieścił tym razem na szyi (choć odwrócił poszczególne kręgi), użył też odmiennych sformułowań. Odpowiadając Leidy’emu twierdził, że do pomyłki doprowadził go fakt ułożenia przez Leidy’ego kręgów Cimoliasaurus na odwrót w opisie tego rodzaju w 1851 i wskazał, że tę rekonstrukcję skorygowano. Odrzucił także identyczność Elasmosaurus i Discosaurus, zauważając, że ten ostatni i Cimoliasaurus nie mają żadnych cech wyróżniających. Mimo prób zniszczenia preprintów jedna z kopii trafiła w ręce amerykańskiego paleontologa Othniela Charlesa Marsha. Marsh twierdził, że natychmiast zauważył pomyłkę Cope’a i zawstydzał rywala, przez dekady przypominając mu o błędzie, co doprowadziło do antagonizmu między badaczami. Ich długoletni konflikt przyciągnął szeroką uwagę opinii publicznej i stał się znany w historii paleontologii, gdyż przekształcił się w tzw. wojnę o kości toczoną przez tych dwóch paleontologów[7][12][14][15][16][17].
W związku z reputacją znakomitego paleontologa, jaką cieszył się Cope, zastanawiano się, czemu popełnił tak oczywisty błąd anatomiczny. Sugerowano, że jako unikalny okaz w 1868 holotypowy Elasmosaurus sprawiał trudności w interpretacji na podstawie dostępnej wtedy wiedzy. Ponadto Cope sądził początkowo, że chodzi o dwa okazy różnych zwierząt – w napisanym w 1868 liście do LeConte’a odnosił się do zakładanego „mniejszego okazu” jako do Discosaurus carinatus. Cope kończył wtedy trzecią dekadę swego życia i nie miał jeszcze paleontologicznego doświadczenia, więc mógł być pod wpływem pomyłki Leidy’ego, który odwrócił kręgosłup Cimoliasaurus. W 2002 amerykańska historyczka sztuki Jane P. Davidson zwróciła uwagę, że wczesne odkrycie błędu Leidy’ego przez innych naukowców przeczy tej hipotezie, dodając, że Cope nie był przekonany, że zaszła pomyłka. Anatomia plezjozaurów była w tamtych czasach wystarczająco dobrze znana i Cope wedle Davidson nie powinien był popełnić takiego błędu[12]. Cope niewiele pracował z okazem od jego opisu w 1870, trzymał go w magazynie prawie 30 lat[7]. Powtórny, dokładny opis tego okazu wyszedł spod pióra niemieckiego paleontologa Svena Sachsa w 2005[6].
Znane i możliwe skamieniałości
[edytuj | edytuj kod]
Niekompletny okaz holotypowy, skatalogowany jako ANSP 10081, jest jedynym niewątpliwym okazem Elasmosaurus. Był długo wystawiany, natomiast obecnie jest przechowywany w zamknięciu wraz z innymi przypisanymi elasmozaurowi fragmentami. Okaz obejmuje kości przedszczękowe, część tylnego fragmentu prawej kości szczękowej, dwa fragmenty kości szczękowych z zębami, przednią część kości zębowych, jeszcze trzy fragmenty szczęki bądź żuchwy, 2 fragmenty czaszki bliżej nieustalonego pochodzenia, 72 kręgi szyjne, w tym atlas i obrotnik, 3 kręgi piersiowe, 6 grzbietowych, 4 krzyżowe, 18 ogonowych, fragmenty żeber[3][6]. W 2013 niekompletny trzon kręgu szyjnego holotypu wspomniany przez Cope’a, ale uznany za zaginiony, został powtórnie odkryty przez Sachsa, zwiększając liczbę kręgów szyjnych z 71 do 72[3]. W 1986 ukończono trójwymiarową rekonstrukcję holotypowego szkieletu, wystawianego jako ANSP. Rzeczony odlew skopiowała później Triebold Paleontology Incorporated, a repliki udostępniono innym muzeom. Jedna z nich mierzy 12,8 m długości[7].

Choć Cope w 1869 i 1875 opisał i zilustrował obręcze kończyny piersiowej i miednicznej Elasmosaurus, fragmenty te już w 1906 r. uznał za zaginione amerykański paleontolog Samuel Wendell Williston. Cope pożyczył je bowiem angielskiemu rzeźbiarzowi Hawkinsowi, by ten wypreparował je z konkrecji, w których częściowo tkwiły. Pełne odsłonięcie kości było potrzebne, gdyż Hawkins chciał stworzyć rekonstrukcję elasmozaura do swojej wystawy „Paleozoic Museum” w nowojorskim Central Parku, która to wystawa miała być amerykańskim odpowiednikiem londyńskiej ekspozycji pełnowymiarowych modeli wymarłych dużych kręgowców tzw. Crystal Palace Dinosaurs w parku Crystal Palace. W maju 1871 wiele eksponatów z pracowni Hawkinsa zniszczyli wandale i ich fragmenty zakopano. Możliwe, że elementy obręczy elasmozaura zostały wówczas zniszczone, choć nie wspominają o ich utracie ani Hawkins ani Cope. Skamieniałości mogące należeć do holotypu znalazł amerykański geolog Benjamin Franklin Mudge w 1871, ale też prawdopodobnie zaginęły[6][7][18].
Dodatkowe skamieniałości plezjozaura odkryto nieopodal pierwotnej lokalizacji okazu holotypowego w 1954, 1991, 1994 i 1998, zaliczają się doń kręgi, żebra (w tym żebra brzuszne, czyli gastralia) i gastrolity. Jako że żaden z tych fragmentów nie pokrywa się z elementami szkieletu holotypu, w 2005 amerykański paleontolog Michael Everhart doszedł do konkluzji, że są to brakujące elementy należące do osobnika holotypowego, którego część zwłok została oddzielona przed pogrzebaniem i zachowała się osobno. Zauważył też, że niewielki kamień zaklinowany w kanale kręgowym jednego z kręgów ogonowych holotypu może być gastrolitem, o czym świadczy to, że jest wypolerowany[19]. W 2007 Leslie Noè i Marcela Gómez-Pérez wyrazili wątpliwość co do przynależności tych dodatkowych znalezisk do holotypu, a nawet do Elasmosaurus z powodu braku dowodów. Wyjaśniali, że elementy brakujące w holotypie mogły ulec erozji bądź po prostu nie zostać zebrane, mogły też ulec zniszczeniu podczas transportu czy preparowania. Gastrolity mogły być potraktowane jako kamienie niezwiązane ze szkieletem i zostać pominięte podczas zbierania okazu holotypowego, jako że w tym czasie nie wiedziano, że mogą one występować u plezjozaurów (udokumentowano to dopiero 10 lat później u innego okazu plezjozaura)[20].
W 2017 Sachs i Joachim Ladwig zasugerowali, że fragmentaryczny szkielet Elasmosauridae z górnego kampanu z Kronsmoor w Szlezwik-Holsztyn w Niemczech, przechowywany w Naturkunde-Museum Bielefeld, może należeć do Elasmosaurus. Dodatkowe części tego szkieletu przechowuje Instytut Geologii Uniwersytetu w Hamburgu, jak też prywatne kolekcje. W sumie okaz obejmuje kręgi szyjne, grzbietowe i ogonowe, paliczki, zęby, 110 gastrolitów i niezidentyfikowane fragmenty[21].
Gatunki dawniej zaliczane do Elasmosaurus
[edytuj | edytuj kod]Przez kolejne lata liczne gatunki elasmozaura opisywali Cope, Williston i inni autorzy. Żaden z nich nie ostał się jednak w rodzaju Elasmosaurus do dnia dzisiejszego. Większość z nich przeniesiono do własnych rodzajów bądź uznano za wątpliwe, nomina dubia bez cech diagnostycznych[6][22][23].

Wraz z opisem E. platyurus w 1869 Cope ustanowił inny gatunek elasmozaura E. orientalis, opierając się na dwóch kręgach grzbietowych z New Jersey[24]. Odróżnił E. orientalis od E. platyurus dzięki silniej rozwiniętym wyrostkom poprzecznym kręgów, wedle autora bliższych Cimoliasaurus. Jednak przypisał on je rodzajowi Elasmosaurus, kierując się znacznym rozmiarem i pochylonym bokiem. Pierwszy z tych kręgów służył za próg w sklepie krawieckim, kolejny został znaleziony w wyrobisku kopalnianym przez Samuela Lockwooda, kuratora oświaty. Cope nadał nowemu gatunkowi epitet orientalis, uwzględniając prawdopodobnie bardziej wschodni zasięg występowania w stosunku do E. platyurus[4]. Leidy w następnym roku przeniósł E. orientalis do obecnie uznawanego za wątpliwy rodzaju Discosaurus[25]. W 1952 Welles uznał gatunek za nomen dubium z powodu fragmentaryczności szczątków[26].
W 1869 Cope opublikował także artykuł o skamieniałościach gadów z New Jersey, w którym opisał E. orientalis jako zwierzę o długiej szyi. Jednak na towarzyszącej tekstowi ilustracji przedstawił elasmozaura o krótkiej szyi w konfrontacji z dryptozaurem (zwanym wtedy Laelaps), obok przypominającego wyglądem plezjozaura mozazaura i innych zwierząt w tle. Wedle Davidsona nie ma pewności, który gatunek elasmozaura przedstawił autor, jeśli jednak chodzi o E. orientalis, krótka szyja pozostaje w sprzeczności z napisanym przez Cope’a tekstem, a jeśli E. platyurus, przedstawił on zwierzę o krótkiej szyi już po stwierdzeniu niepoprawności takiej interpretacji. Davidson zasugerował, że nawet jeśli Leidy wskazał błąd Cope’a w 1868, Cope nie musiał go uznać[12][27]. W odpowiedzi danej Leydy’emu w 1870 Cope twierdził, że przeniesienie E. orientalis do innego rodzaju jest wątpliwe, w związku z czym przedstawił go z krótką szyją zgodnie z poglądem na Cimoliasaurus. W sytuacji, gdy więcej szczątków wskazało na obecność u E. orientalis długiej szyi, stwierdził, że obrazek może przedstawiać raczej Cimoliasaurus[28].

W tej samej pracy z 1869, gdzie nazywał E. platyurus i E. orientalis, Cope przypisał rodzajowi jeszcze kolejny gatunek, E. constrictus[4], bazując na fragmentarycznym trzonie kręgu szyjnego znalezionym w turońskich osadach ilastych w Steyning w Sussex w Wielkiej Brytanii. Został on opisany przez brytyjskiego paleontologa Richarda Owena jako Plesiosaurus constrictus w 1850. Owen ukuł nazwę, zwracając uwagę na niezwykle wąski odstęp pomiędzy wyrostkami łączącymi się z żebrami. Uznał to częściowo za efekt wtórnej deformacji w trakcie procesu fosylizacji, nie potrafił jednak wyjaśnić, czemu kompresji uległa jedynie część środkowa, a nie stawowo łączące się końce trzonu[29]. Cope uznał to więc za stan naturalny, a P. constrictus za gatunek elasmozaura bądź jego krewnego[4]. W 1962 Welles uznał P. constrictus za nomen dubium z uwagi na fragmentaryczność[30][31]. Per Ove Persson utrzymał go jako pewny w 1963, zwracając uwagę na podłużny grzebień po bokach trzonu jako na cechę Elasmosauridae[32]. W 1995 Nathalie Bardet i Pascal Godefroit także rozpoznali w nim przedstawiciela rodziny elasmozaurów, choć nieokreślonego bliżej rodzaju[33].

Cope odkrył kolejny szkielet Elasmosauridae w 1876. Zaliczył go do nowego gatunku – E. serpentinus w 1877, odróżniając go po braku kompresji w tylnych kręgach szyjnych, obecności kilku specyficznie osadzonych żeber na pierwszych kręgach grzbietowych i obecności „słabych kątów” poniżej proksymalnych kręgów ogonowych. Odkrył on również inny duży szkielet wykazujący znaczne podobieństwo do znanych szczątków E. orientalis z czarnych łupków „kredowej warstwy nr 4". Wydobył go z pomocą George’a B. Cledenninga i kapitana Nicholasa Buesena[34]. W 1943 Welles przeniósł E. serpentinus z Elasmosaurus do nowego rodzaju Hydralmosaurus[35]. W 2016 wszystkie okazy Hydralmosaurus znalazły się w rodzaju Styxosaurus, czyniąc wcześniej stosowane miano nomen dubium[36]. Williston opublikował rysunek innego okazu E. serpentinus w 1914[37]; Elmer Riggs dokonał formalnego opisu w 1939[38]. Welles przeniósł okaz do nowego rodzaju i gatunku pod nazwą Alzadasaurus riggsi w 1943[35]. Kenneth Carpenter przypisał go do Thalassomedon haningtoni w 1999[22]. Sachs, Johan Lindgren i Benjamin Kear w 2016 zauważyli, że szczątki należały do młodocianego osobnika i były bardzo zdeformowane, wobec czego woleli uznać wcześniejsze oznaczenia tego okazu za nomen dubium[39].
Seria 19 kręgów szyjnych i grzbietowych z rejonu Big Bend Dam w Missouri, z formacji Pierre Shale, znaleziona została przez Johna H. Charlesa. Cope, otrzymawszy te kości w Academy of Natural Sciences, uznał ich przynależność do kolejnego gatunku elasmozaura. Wedle badacza rzeczone kręgi były najkrótsze wśród członków tego rodzaju (przypominające pod tym względem Cimoliasaurus), jednak wciąż zaliczał je do Elasmosaurus z powodu ich kompresji. W 1894 zaliczył je do E. intermedius[40], jednak w rewizji północnoamerykańskich plezjozaurów z 1906 przez Williston uznał wszystkie te kręgi za „bardziej lub mniej uszkodzone”. Badacz nie znalazł wyraźnych różnic między szczątkami E. intermedius i E. platyurus[18]. W 1952 Welles sądził, że jeśli E. intermedius jest pewny, to musiałby być zaliczony do pliozaurów[26]; jednak w 1962 wciąż określał owe szczątki jako nomen dubium[30]. Trzy krótsze kręgi znalezione wśród pozostałości E. intermedius Cope przypisał do nowego rodzaju i gatunku Embaphias circulosus[40], który Welles znów uznał za nomen dubium w 1962[30].
Williston nazwał liczne nowe gatunki elasmozaura w swej rewizji z 1906[41]. W 1874 wraz z Mudgem odkryli okaz w Plum Creek w Kansas[18], początkowo przypisując go w 1890 do nowego gatunku Cimoliasaurus, C. snowii[42]. Później badacz ów rozpoznał w nim charakterystyczną dla rodziny elasmozaurów naturę kości ramiennej i kruczej. Wobec tego nazwał znalezisko E. snowii. Drugi okaz, odkryty przez Eliasa Westa w 1890, również przypisany został przezeń do gatunku E. snowii[18]. W 1943 Welles przeniósł E. snowii do odrębnego rodzaju Styxosaurus[35], gdzie gatunek ten pozostał. Jednak okaz Westa Welles zaliczył w 1952 do Thalassiosaurus ischiadicus[26]. Carpenter przywrócił go do S. snowii w 1999[22][41]. Williston przeniósł też gatunek E. ischiadicus z rodzaju Polycotylus, gdzie został początkowo umieszczony przezeń przy nadawaniu nazwy w 1903. Szczątki typowe odkrył on w trakcie tej samej wyprawy w 1874 z Mudgem. Williston odnosił się też do innego okazu odkrytego przez Mudge’a i H.A. Brousa w 1876[18]. W 1943 oba okazy zaliczono do nowego rodzaju Thalassiosaurus, czego dokonał Welles[35], przypisując tego ostatniego później do Alzadasaurus kansasensis w 1952[26]. Glenn Storrs uznał oba za nieokreślone bliżej elasmozaurydy w 1999[43]. W tym samym roku Carpenter zaliczył oba do Styxosaurus snowii[22][41].

W 1889 Handel Martin znalazł okaz Elasmosauridae w hrabstwie Logan w Kansas. Williston nadał mu nową nazwę gatunkową E. (?) marshii. Zachował jednak rezerwę co do przypisania go do rodzaju, uznając możliwość, że należy do rodzaju innego, niż zaproponowany przezeń[18]. W 1943 przeniósł E. (?) marshii do własnego rodzaju Thalassonomosaurus[35]; jednak w 1999 Carpenter wrzucił T. marshii do Styxosaurus snowii[22]. Inny gatunek, E. nobilis, nazwany został przez Willistona dzięki bardzo dużym szczątkom odkrytym przez Mudge’a w 1874 w hrabstwie Jewell w Kansas[18]. Welles przeniósł E. nobilis do rodzaju Thalassonomosaurus, tworząc T. nobilis w 1943[35], określony z kolei jako S. snowii przez Carpentera[22]. W końcu Williston dwa wyjątkowo duże kręgi grzbietowe zebrane przez Charlesa Sternberga w 1895 nazwał E. sternbergii, a Storrs uznał je za nieokreślone[41][43]. Williston wymienił trzy dodatkowe gatunki Elasmosaurus, które zobrazował i opisał później[18]. Znów odniósł się do nowego gatunku elasmozaura z Kansas w 1908[44].
Do rodzaju elasmozaur przypisał też kilka gatunków bazujących na słabo zachowanych szczątkach kręgów z Rosji N.N. Bogolubow w 1911. Jednym z nich był E. helmerseni, opisany przez W. Kiprijanowa w 1882 z Maloje Serdoba, z obwodu saratowskiego, jako Plesiosaurus helmerseni. Materiał ze szwedzkiej Skanii przypisano E. helmerseni w 1885 piórem H. Schrödera[45]. Pozostałości kręgów i kończyn[46] z Kurska początkowo zaliczone przez Kiprijanowa do P. helmerseni zostały przeniesione przez Bogolubowa do nowego gatunku E. kurskensis, który uznany został za „identyczny bądź spokrewniony z elasmozaurem”. Wspomniany badacz nazwał też E. orskensis, opierając się o „bardzo duże” kręgi szyjne i ogonowe z Konopljanki w obwodzie orenburskim, i E. serdobensis, w oparciu o pojedynczy kręg szyjny z Maloje Serdoba[47]. Jednakże pewność wszystkich tych gatunków została zakwestionowana. Welles uznał E. kurskensis za nieokreślonego bliżej plezjozaura w 1962[30]. Persson zauważył w swym przeglądzie z 1959 omawiającym materiał szwedzkiego „E.” helmerseni, że o ile gatunek był prawdopodobnie bliskim krewnym właściwego Elasmosaurus, materiał jest zbyt fragmentaryczny dla oceny rozważanej hipotezy[45]. W 1963 zaznaczył w odniesieniu do trzech rozpatrywanych gatunków, że „ich określenie rodzajowe i gatunkowe jest dyskusyjne”, choć odmówił określenia ich jako niepewne, powołując się na to, że nie widział materiału kopalnego[32]. Podobnie w 1999 Jewgenij Perwuszow, Maksim Archangielskij i A.W. Iwanow uznali E. helmerseni za nieokreślonego bliżej elasmozauryda[48]. W 2000 Storrs, Archangielskij i Władimir Efimow zgodzili się z Wellesem co do E. kurskensis, jak również uznali E. orskensis i E. serdobensis za bliżej nieokreślone elasmozaurydy[49].
Dwa kolejne gatunki z Rosji zostały opisane przez innych autorów. A.N. Riabinin opisał pojedynczy paliczek płetwy w 1915 jako E. (?) sachalinensis. Nazwał gatunek, odwołując się do wyspy Sachalin, gdzie N.N. Tichonowicz znalazł tę kość w 1909[50]. Jednak okaz nie może być określony bliżej niż jako elasmozauryd, który to pogląd poparł Persson[32] oraz Pervwuszow i współpracownicy[48]. Storrs, Archangielskij i Efimow określili ten szczątek jako należący do niezidentyfikowanego bliżej plezjozaura[49], później zaś podobnie Aleksander Awerianow i W.K. Popow w 2005[50]. W 1916 P.A. Prawosławlew nazwał E. amalitskii z regionu Donu, opierając się na okazie obejmującym kręgi, obręcze kończyn i kości kończyn. Persson uznał go za pewny gatunek i względnie dużego przedstawiciela Elasmosauridae[32]. Jednak, jak w przypadku E. (?) sachalinensis, Perwuszow i inni uznali E. amalitskii za nieokreślonego bliżej elasmozauryda[48].

W 1918 w przeglądzie rozmieszczenia geograficznego i ewolucji Elasmosaurus, Prawosławlew prowizorycznie zaliczył 3 opisane wcześniej gatunki do elasmozaura[46]. Jego pogląd nie zdobył szerokiego poparcia. Do tych gatunków należał E. chilensis, oparty na chilijskim Plesiosaurus chilensis, pojedynczym kręgu ogonowym z Claude Gay z 1848[51]. Wilhelm Deecke przeniósł gatunek do rodzaju pliozaur w 1895[52], co uznał Prawosławlew. Edwin Colbert później, w 1949, przypisał typowy kręg do Pliosauroidea, inne szczątki uznając za należące do nieokreślonego bliżej elasmozauroida[53][54]. Typowy kręg potencjalnie należeć mógł do Aristonectes parvidens wedle publikacji José O’Gormana i współpracowników z 2013[55]. Innym był E. haasti, pierwotnie Mauisaurus haasti, nazwany przez Jamesa Hectora w 1874 na podstawie szczątków znalezionych w Nowej Zelandii. Choć uznawany za pewny przez znaczny czas, od 2017 uznaje się go za nomen dubium[56]. Prawosławlew rozpoznał kolejny gatunek z Nowej Zelandii, E. hoodii, nazwany przez Owena w 1870 Plesiosaurus hoodii w oparciu o kręg szyjny[57]. Welles uznał go za nomen dubium w 1962[30]; Joan Wiffen i William Moisley zgodzili się z nim w przeglądzie nowozelandzkich plezjozaurów z 1986[58].
W 1949 Welles nazwał nowy gatunek elasmozaura E. morgani dzięki dobrze zachowanemu szkieletowi z Hrabstwa Dallas w Teksasie[59]. Jednak część okazu została przypadkowo wyrzucona podczas relokacji kolekcji paleontologicznej Southern Methodist University[60]. Welles dostrzegł podobieństwo E. morgani do E. platyurus w jego obręczy kończyny piersiowej, utrzymał jednak oddzielne gatunki z powodu krótszej szyi i mocniejszej budowy dystalnych kręgów szyjnych[59]. W 1997 Carpenter ponownie przemyślał różnice między gatunkami i uznał za właściwe wydzielenie E. morgani do własnego rodzaju Libonectes[61]. Pomimo przenosin i utraty materiału L. morgani traktowany jest często jako archetypowy elasmozauryd. Dane oparte na utraconych fragmentach były uznawane bez zastrzeżeń w kolejnych analizach filogenetycznych, nim powtórny opis zachowanych fragmentów opublikowali Sachs i Benjamin Kear w 2015[60].
Persson zaliczył jeszcze inny gatunek do Elasmosaurus w swym opublikowanym w 1959 opisie szczątków „E.” helmerseni ze Szwecji, E. (?) gigas. Oparł się o Schröderowskiego Pliosaurus (?) gigas, nazwanego w 1885 dzięki dwóm kręgom grzbietowym, z których jeden znaleziono w Prusach, a drugi w Skanii. Były niekompletne, ale Persson uznał, że ich proporcje i kształt części stawowych różnią się znacznie od pliozauroidowych, a pasują do spotykanych u elasmozaurydów. Ponieważ w czasie publikacji tej pracy nie było argumentów przeciwko uznaniu ich za najbardziej zbliżone do Elasmosaurus, badacz z wahaniem umieścił je więc właśnie w tym rodzaju. Theodor Wagner wcześniej zaliczył gatunek do Plesiosaurus w 1914[45]. W 2013 kwestia ta pozostawała jeszcze nierozwiązana[62]. Inny gatunek z Rosji, E. antiquus, nazwali Dubejkowski i Oczew w 1967[49] z kamieniołomu fosforytów Kamsko-Wiatskoj, jednak Perwuszow i inni w 1999, a później Storrs i współpracownicy w 2000 uznali go za nieokreślonego bliżej elasmozauryda[48][49].
Systematyka
[edytuj | edytuj kod]
Choć Cope pierwotnie uznał Elasmosaurus za plezjozaura, w pracy z 1869 umieścił go wraz z Cimoliasaurus i Crymocetus w nowym rzędzie zauropterygów, nazwanym Streptosauria, „odwrócone jaszczury”, z powodu ułożenia ich kręgów, które zdawały mu się odwrócone w porównaniu z kręgami innych zwierząt morskich[17][63]. Następnie porzucił ten pogląd w swym opisie elasmozaura z 1869, gdzie przyznał, że oparł się na błędnej interpretacji Leidy’ego dotyczącej Cimoliasaurus. W tej pracy ukuł też bez komentarza nową rodzinę Elasmosauridae, włączając doń Elasmosaurus i Cimoliasaurus. W jej obrębie za cechę wyróżniającą tego pierwszego uznał dłuższą szyję o bardziej zbitych kręgach, drugiego zaś – krótszą szyję o wgłębionych, kwadratowych kręgach[4].

W kolejnych latach Elasmosauridae stały się jedną z trzech grup plezjozaurów, obok Pliosauridae i Plesiosauridae (niekiedy łączonych w jedną)[64]. Charles Andrews zajmował się różnicami pomiędzy elasmozaurydami a pliozaurydami w 1910 i 1913. Odróżniał elasmozaurydy po długiej szyi i niewielkiej głowie, jak też sztywnej i dobrze rozwiniętej łopatce (ale obojczykach i kości międzyobojczykowej w zaniku, niekiedy zupełnie nieobecnej) związanej z poruszaniem się dzięki kończynom przednim napędzającym ciało. Natomiast pliozaurydy miały krótkie szyje i duże głowy, napęd zapewniały im kończyn tylne[65][66]. Choć zaliczenie Elasmosaurus do Elasmosauridae nie budziło kontrowersji, opinie o pokrewieństwach rodziny różniły się od siebie w kolejnych dekadach. Williston stworzył zrewidowaną taksonomię plezjozaurów w 1925[67].
W 1940 Theodore White opublikował hipotezę dotyczącą pokrewieństwa pomiędzy różnymi rodzinami plezjozaurów. Uznał Elasmosauridae za najbliższe Pliosauridae, zwracając uwagę na ich względnie wąskie kości krucze, jak też brak obojczyków bądź kości międzyobojczykowych. Jego diagnoza Elasmosauridae zwracała też uwagę na umiarkowaną długość czaszki (czaszka mezocefaliczna), żebra szyjne o jednej bądź dwóch główkach, łopatkę i kość kruczą kontaktujące się w linii pośrodkowej, tępy tylny kąt łopatki i parę okien w kompleksie łopatkowo-kruczym rozdzielonych wałem kostnym węższym, niż obserwowany u pliozaurydów. Cytowana zmienność liczby główek żeber szyjnych powstaje na skutek włączenia przez badacza Simolestes do Elasmosauridae, jako że cechy czaszki i obręczy kończyny piersiowej pasowały autorowi bardziej do elasmozaura, niż do Pliosaurus lub Peloneustes. Uznał on Simolestes za możliwego przodka elasmozaura[68]. Oskar Kuhn przyjął podobną klasyfikację w 1961[32].
Welles podjął kwestię klasyfikacji White’a w rewizji plezjozaurów z 1943, zwracając uwagę, że na cechy zauważone przez owego autora wpływały i zachowanie szczątków, i ontogeneza. Podzielił zamiast tego plezjozaury na dwie nadrodziny, Plesiosauroidea i Pliosauroidea, bazując na długości szyi, wielkości głowy, długości kości kulszowej oraz smukłości kości ramiennej i udowej (propodialiów). Każdą z tych nadrodzin podzielił dalej dzięki liczbie główek żeber i proporcjom epipodialiów. Tak więc elasmozaurydy miały długie szyje, małe głowy, krótkie kości kulszowe, korpulentne propodialia, jednogłowe żebra i krótkie epipodialia[35]. Pierre de Saint-Seine w 1955 i Alfred Romer w 1956 adoptowali klasyfikację Wellesa[32]. On sam w 1962 dalej podzielił Elasmosauridae w oparciu o obecność belki kostnej w miednicy tworzonej przez połączenie kości kulszowych, łącząc Elasmosaurus i Brancasaurus w podrodzinę Elasmosaurinae na podstawie całkowicie zamkniętych belek[30].

Carpenter wykonał opublikowaną w 1997 analizę filogenetyczną plezjozaurów i zmienił tradycyjny podział plezjozaurów opierający się na długości szyi. O ile Polycotylidae zaliczano wcześniej do Pliosauroidea, Carpenter przeniósł je, uznając za grupę siostrzaną Elasmosauridae, bazując na podobieństwach między nimi, co oznacza, że Polycotylidae i Pliosauroidae wyewoluowały swe krótkie szyje niezależnie[61]. Zawartość Elasmosauridae również została dogłębniej zbadana. Od początkowego umieszczenia wśród elasmozaurydów obecność w tej rodzinie Brancasaurus uznawano za dobrze wspartą, co zachowała jaszcze analiza O’Keefe’a z 2004[69] i analiza Franziski Großmann z 2007[70]. Jednak analiza Ketchuma i Bensona włączyła go w Leptocleidia[71], co potwierdziły kolejne badania[72][73][36]. Przeniesiono także Muraenosaurus do Cryptoclididae, a Microcleidus i Occitanosaurus do Plesiosauridae[71]; Benson i Druckenmiller wyodrębnili dwa ostatnie w Microcleididae w 2014, uznając Occitanosaurus za gatunek Microcleidus[73]. Rodzaje te wcześniej były uznawane za elasmozaurydy przez Carpentera, Großmann i innych badaczy[22][70][74][75].
W obrębie Elasmosauridae Elasmosaurus uznawano za „wildcard taxon” o bardzo różnych proponowanych pokrewieństwach[76]. Analiza Carpentera z 1999 widziała go jako bardziej bazalnego od innych elasmozaurydów z wyjątkiem Libonectes[22]. W 2005 Sachs zasugerował jego bliskie pokrewieństwo ze Styxosaurus[6]. W 2008 Druckenmiller i Russell umieścili go w politomii z dwiema grupami, jedną obejmującą Libonectes i Terminonatator, drugą zawierającą Callawayasaurus i Hydrotherosaurus[77]. Ketchum i Benson w 2010 włączyli Elasmosaurus do pierwszej z tych grup[71]. Benson i Druckenmiller w 2013 (wynik ich analizy poniżej) usunęli z niej Terminonatator, umieszczając go jako bardziej zaawansowaną formę[72]. Rodrigo Otero w 2016 oparł swą analizę na modyfikacji tego samego zbioru danych (poniżej), ukazując elsmozaura jako najbliższego krewniaka Albertonectes, tworzącego Styxosaurinae ze Styxosaurus i Terminonatator[36]. Danielle Serratos, Druckenmiller i Benson nie mogli rozwikłać pozycji elasmozaura w 2017, zauważyli jednak, że Styxosaurinae byłoby synonimem Elasmosaurinae, jeśliby zaliczyć do nich Elasmosaurus[76].


|
Topologia A: Benson et al. (2013)[72]
|
Topologia B: Otero (2016)[36]
|
Paleobiologia
[edytuj | edytuj kod]
Elasmosauridae w pełni przystosowały się do życia w oceanie. Miały opływowy kształt ciała i długie płetwy, wskazujące, że chodzi o aktywnych pływaków[2]. Niezwykła budowa ciała elasmozaurydów ograniczała szybkość, z jaką potrafiły pływać, a ich płetwy poruszały się na sposób przypominający ruchy ptasich skrzydeł, nie mogąc jednak obracać się trzymane były sztywno[78]. Plezjozaury były prawdopodobnie zdolne do utrzymania wysokiej temperatury ciała, pozwalającej na długotrwałe pływanie[79]. Praca z 2015 zaprezentowała konkluzję, zgodnie z którą ruch napędzały głównie przednie płetwy, tylne zaś zapewniały stabilność i manewrowość[80], natomiast badanie z 2017 pokazało, że tylne płetwy plezjozaurów dawały 60% więcej odrzutu i 40% więcej wydajności, kiedy poruszały się w harmonii z przednimi[81]. Płetwy plezjozaurów były tak sztywne i wyspecjalizowane do pływania, że uniemożliwiały zwierzętom wychodzenie na ląd i składanie tam jaj sposobem żółwi morskich. Wobec tego prawdopodobnie rodziły one młode jak węże morskie[82]. Dowodu na żyworodność plezjozaurów dostarcza skamieniałość dorosłej Polycotylus z pojedynczym płodem w obrębie ciała[83].
Ruchy i funkcja szyi
[edytuj | edytuj kod]
Cope w 1869 porównał budowę i zwyczaje Elasmosaurus do wężowych. Jako że sugerował, iż kręgosłup tułowia nie pozwalał na ruchy pionowe z powodu wydłużonych wyrostków kolczystych tworzących prawie że ciągłą linię z pozostawieniem bardzo małych przestrzeni pomiędzy przylegającymi do siebie kręgami, wyobrażał sobie znacznie bardziej giętki ogon i szyję. Pisał o wężowej głowie wzniesionej wysoko w powietrzu bądź zanurzonej wedle woli zwierzęcia, niekiedy wygiętej jak u łabędzia, oczekującej, by rzucić się na rybę, niekiedy zaś wyprostowanej, wystającej nad wodę bądź przegiętej w dół i eksplorującej głębie[4].
Pomimo przedstawień w popularnych mediach bardziej współczesne badania wykazały, że Elasmosaurus nie był zdolny podnieść głowę wyżej, niż nad wodę. Masa długiej szyi lokowała środek ciężkości za przednimi płetwami. Wobec tego Elasmosaurus mógł jedynie unosić głowę i szyję, przebywając w płytkiej wodzie, kiedy reszta ciała spoczywała na dnie. Ponadto masa szyi, jej ograniczone umięśnienie i ograniczone ruchy między kręgami uniemożliwiały elasmozaurowi podnoszenie głowy i szyi bardzo wysoko. Głowa i ramiona Elasmosaurus prawdopodobnie działały jako ster. Jeśli zwierzę skierowało przednią część ciała w pewnym kierunku, reszta ciała również się weń kierowała. Tak więc Elasmosaurus nie mógł płynąć w jednym kierunku, trzymając głowę i szyję poziomo lub pionowo w innym kierunku[84].

W jednym z badań odkryto, że szyje elasmozaurydów miały zakres ruchu brzusznie 75–177°, grzbietowo 87–155°, bocznie 94–176°, wartości zależą od ilości tkanki między kręgami, która prawdopodobnie zwiększała sztywność szyi wzdłuż grzbietu. Badacze doszli do wniosku, że wykonywanie bocznych i pionowych łuków i płytkich krzywych s-kształtnych było możliwe w przeciwieństwie do łąbędziej S-kształtnej sylwetki wymagającej ponad 360° zgięcia pionowego[85].
Dokładna funkcja szyi elasmozaurydów nie została poznana[2]. Szyja mogła pełnić ważną rolę w polowaniu[78]. Sugerowano także, że długie szyje plezjozaurów działały jak rurka i pozwalały zwierzętom oddychać powietrzem, gdy większość ciała spoczywała pod wodą. Pomysł ten wywołał spory, w których zwracano uwagę na duże różnice ciśnienia hydrostatycznego, szczególnie dotykałyby one długoszyich elasmozaurydów. Anatomia szyi tych zwierząt pozwalała im na delikatne nachylenie jej, umożliwiające oddychanie przy powierzchni, ale pływanie pod powierzchnią wymagałoby znacznych wydatków energetycznych. W dodatku długa szyja zwiększa także martwą przestrzeń, wobec której zauropsyd potrzebuje większych płuc. Szyja mogła powodować również inne problemy, na przykład stanowić dogodny cel ataku drapieżników[86].
Odżywianie się
[edytuj | edytuj kod]
W 1869 Cope zauważył łuski i zęby sześciu gatunków ryb bezpośrednio pod kręgami holotypu Elasmosaurus. Wyjaśnił to tezą, że ryby stanowiły dietę zwierzęcia. Dzięki tym pozostałościom Cope nazwał nowy gatunek barrakudowatych Sphyraena carinata[4].
Zgięcia elasmozaurzej szyi pozwalałyby zwierzęciu wykorzystywać liczne metody polowań takie jak benthic grazing (pasienie się na dnie morskim), podczas którego zwierzę pływało w pobliżu dna, sięgając po przebywającą na nim zdobycz swoją głową i szyją. Elasmosauridae mogły również aktywnie polować w pelagialu, cofając swą szyję, by wyprowadzić potem uderzenie, bądź wykonując nią zamachy na boki, by ogłuszyć lub zabić zdobycz swymi sterczącymi na boki zębami (jak u współczesnych piłonosowatych)[85].
Możliwe, że Elasmosaurus i jego krewni atakowali rybie ławice, ukrywając się pod nimi w głębinach i unosząc powoli głowy, gdy się doń zbliżyli. Oczy zauropsyda leżały na szczycie głowy, umożliwiając mu bezpośrednie widzenie do góry. Widzenie stereoskopowe pomagałoby mu znajdować niewielką zdobycz. Polowanie od dołu byłoby także możliwe dzięki widzeniu sylwetki ofiary w świetle słonecznym przez drapieżnika ukrywającego się w wodach poniżej. Elasmosauridae prawdopodobnie zjadały niewielkie ryby kostnoszkieletowe i bezkręgowce morskie. Ich niewielkie, nieruchliwe czaszki ograniczały wielkość zdobyczy, którą zauropterygi te mogły spożyć. Ponadto ze swymi długimi, smukłymi zębami zaadaptowanymi do chwytania zdobyczy, ale nie do jej rozszarpywania Elasmosauridae najprawdopodobniej połykały ją w całości[78][85].
Choć w elasmozaurydowych szczątkach powszechnie znajdowano kilka gastrolitów, Elamosaurus był jedynym niekontrowersyjnym znaleziskiem z kamieniami leżącymi w łuku kręgowym jednego z dystalnych kręgów ogona[20]. Okaz blisko spokrewnionego z nim Styxosaurus zawierał kamienie i pofragmentowane kości rybie w okolicy brzucha, za obręczą kończyny piersiowej. Szczątki rybie zidentyfikowano jako Enchodus i śledziopodobne. Kamienie pasują do skały znajdującej się 600 km od miejsca znalezienia tego okazu[87]. Zaproponowano kilka różnych funkcji gastrolitów, jak wspomaganie trawienia, mieszanie masy pokarmowej, suplementacja soli mineralnych, kontrola i zachowanie wyporności[88]. Elasmosauridae przypuszczalnie także padały ofiarami polowań. Kość ramienna niezidentyfikowanego bliżej prawie dorosłego elasmozauryda nosi ślady zębów rekina Cretoxyrhina[89], natomiast zmiażdżona czaszka Woolungasaurus nosi ślady zębów pasujących do pliozaura Kronosaurus[90].
Paleoekologia
[edytuj | edytuj kod]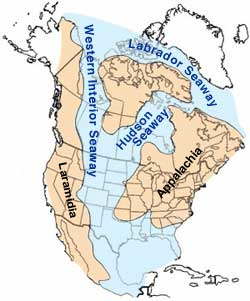
Elasmosaurus znany jest z ogniwa Sharon Springs kampańskiej (kreda górna) formacji Pierre Shale z zachodniego Kansas, datowanej na około 80,5 miliona lat. Pierre Shale reprezentuje fazę tworzenia się osadów morskich Morza Środkowego Zachodu, płytkiego morza śródlądowego rozpościerającego się na terenach dzisiejszej Ameryki Północnej w okresie kredowym[91]. Za największego swego zasięgu Morze Środkowego Zachodu zajmowało tereny od obecnych Gór Skalistych do Appalachów, i miało szerokość około 1000 km. Osiągało jedynie 800–900 m głębokości. Dwa wielkie kontynentalne zlewiska zaopatrywały je ze wschodu i z zachodu, niosąc z wodą wyerodowany z lądu muł tworzący delty rzek wzdłuż niskich brzegów. Na wschodzie sedymentacja była niewielka, na zachodzie zaś znaczna i osadził się tam gruby kompleks osadów ze zerodowanych mas lądowych[92][93]. Zachodni brzeg rozwinął wobec tego znaczącą zmienność, zależąc od zmian poziomu morza i dostaw osadów[92].
O ile miękkie, muliste dno morskie odbierało prawdopodobnie niewielką ilość światła słonecznego, roiło się na nim od życia dzięki stabilnemu strumieniowi szczątków organicznych pochodzących z planktonu i innych organizmów z kolumny wody. Na dnie dominował wielki małż Inoceramus, żyły tam ostrygi. Bioróżnorodność była niewielka. Muszle małży zbierały się przez stulecia, tworząc podmorskie warstwy, zapewniając kryjówki małym rybom. Inne znane z tych mórz zwierzęta obejmują przedstawicieli małży Hippuritoida, liliowce i głowonogi, w tym kalmary i amonity[94].
Morza te zamieszkiwały też wielkie ryby, jak kostnoszkieletowe Pachyrhizodus, Enchodus, Cimolichthys, Saurocephalus, Saurodon, Gillicus, Ichthyodectes, Xiphactinus, Protosphyraena i Martinichthys[95]; rekiny Cretoxyrhina, Cretolamna, Scapanorhynchus, Pseudocorax i Squalicorax[96]. Poza elasmozaurem gady morskie reprezentowały plezjozaury Libonectes, Styxosaurus, Thalassomedon, Terminonatator, Polycotylus, Brachauchenius, Dolichorhynchops i Trinacromerum[97]; mozazaury Mosasaurus, Halisaurus, Prognathodon, Tylosaurus, Ectenosaurus, Globidens, Clidastes, Platecarpus i Plioplatecarpus[11]; żółwie morskie Archelon, Protostega, Porthochelys i Toxochelys[98]. Towarzyszyły im nielotne ptaki wodne, jak Hesperornis[99]. Latały natomiast pterozaury Pteranodon i Nyctosaurus[100] oraz ptak Ichthyornis[99], znane także z lądu[101].
Przypisy
[edytuj | edytuj kod]- ↑ a b c M.P. Taylor, M.J. Wedel. Why sauropods had long necks; and why giraffes have short necks. „PeerJ”. 1, s. e36, 2013. DOI: 10.7717/peerj.36. PMID: 23638372. PMCID: PMC3628838.
- ↑ a b c d e f g S. Sachs, B.P. Kear. Fossil Focus: Elasmosaurs. „palaeontologyonline”. 5, s. 1–8, 2015. Palaeontology Online. [dostęp 2018-01-18].
- ↑ a b c d e f S. Sachs, B.P. Kear, Michael Everhart. Revised Vertebral Count in the „Longest-Necked Vertebrate” Elasmosaurus platyurus Cope 1868, and Clarification of the Cervical-Dorsal Transition in Plesiosauria. „PLoS ONE”. 8 (8), s. e70877, 2013. DOI: 10.1371/journal.pone.0070877. PMID: 23940656. PMCID: PMC3733804. Bibcode: 2013PLoSO...870877S.
- ↑ a b c d e f g h i E.D. Cope. Synopsis of the extinct Batrachia, Reptilia and Aves of North America, Part I. „Transactions of the American Philosophical Society”. 14, s. 44–55, 1869. DOI: 10.5962/bhl.title.60482.
- ↑ J.P. O’Gorman. A Small Body Sized Non-Aristonectine Elasmosaurid (Sauropterygia, Plesiosauria) from the Late Cretaceous of Patagonia with Comments on the Relationships of the Patagonian and Antarctic Elasmosaurids. „Ameghiniana”. 53 (3), s. 245–268, 2016. DOI: 10.5710/AMGH.29.11.2015.2928.
- ↑ a b c d e f g h i j k l m n o S. Sachs. Redescription of Elasmosaurus platyurus, Cope 1868 (Plesiosauria: Elasmosauridae) from the Upper Cretaceous (lower Campanian) of Kansas, U.S.A. „Paludicola”. 5 (3), s. 92–106, 2005.
- ↑ a b c d e f g h M.J. Everhart. Captain Theophilus H. Turner and the Unlikely Discovery of Elasmosaurus platyurus. „Transactions of the Kansas Academy of Science”. 120 (3–4), s. 233–246, 2017. DOI: 10.1660/062.120.0414.
- ↑ Everhart 2005 ↓, s. 121–123.
- ↑ J.P. Davidson, M.J. Everhart. Scattered and Shattered: A Brief History of the Early Methods of Digging, Preserving and Transporting Kansas Fossils. „Transactions of the Kansas Academy of Science”. 120 (3–4), s. 247–258, 2017. DOI: 10.1660/062.120.0416.
- ↑ E.D. Cope. Remarks on a new enaliosaurian, Elasmosaurus platyurus. „Proceedings of the Academy of Natural Sciences of Philadelphia”. 20, s. 92–93, 1868.
- ↑ a b Everhart 2005 ↓, s. 160–168.
- ↑ a b c d J.P. Davidson. Bonehead mistakes: The background in scientific literature and illustrations for Edward Drinker Cope’s first restoration of Elasmosaurus platyurus. „Proceedings of the Academy of Natural Sciences of Philadelphia”. 152 (1), s. 215–240, 2002. DOI: 10.1635/0097-3157(2002)152[0215:HPOVBM]2.0.CO;2.
- ↑ J. Leidy. Remarks on Elasmosaurus platyurus. „Proceedings of the Academy of Natural Sciences of Philadelphia”. 22, s. 9–10, 1870.
- ↑ O.C. Marsh: Wrong End Foremost. [w:] New York Herald [on-line]. 1890. [dostęp 2009-02-02].
- ↑ E.D. Cope. On Elasmosaurus platyurus Cope. „American Journal of Science”. 50 (148), s. 140–141, 1870.
- ↑ E.D. Cope. Synopsis of the extinct Batrachia, Reptilia and Aves of North America, Part I. „Transactions of the American Philosophical Society”. 14, s. 44–55, 1870. DOI: 10.5962/bhl.title.60499.
- ↑ a b G.W. Storrs. Elasmosaurus platyurus and a page from the Cope-Marsh war. „Discovery”. 17 (2), s. 25–27, 1984.
- ↑ a b c d e f g h S.W. Williston. North American plesiosaurs Elasmosaurus, Cimoliasaurus, and Polycotylus. „American Journal of Science”. s4-21 (123), s. 221–236, 1906. DOI: 10.2475/ajs.s4-21.123.221.
- ↑ M.J. Everhart. Elasmosaurid remains from the Pierre Shale (Upper Cretaceous) of western Kansas. Possible missing elements of the type specimen of Elasmosaurus platyurus Cope 1868?. „PalArch’s Journal of Vertebrate Palaeontology”. 4 (3), 2005.
- ↑ a b L.F. Noè, M. Gómez–Pérez. Postscript to Everhart, M.J. 2005. „Elasmosaurid remains from the Pierre Shale (Upper Cretaceous) of western Kansas. Possible missing elements of the type specimen of Elasmosaurus platyurus Cope 1868?” – PalArch’s Journal of Vertebrate Palaeontology 4, 3: 19–32. „PalArch’s Journal of Vertebrate Palaeontology”. 2 (1), 2007.
- ↑ S. Sachs, J. Ladwig. Reste eines Elasmosauriers aus der Oberkreide von Schleswig-Holstein in der Sammlung des Naturkunde-Museums Bielefeld. „Berichte des Naturwissenschaftlichen Vereins für Bielefeld und Umgegend”. 55, s. 28–36, 2017. (niem.).
- ↑ a b c d e f g h K. Carpenter. Revision of North American elasmosaurs from the Cretaceous of the western interior. „Paludicola”. 2 (2), s. 148–173, 1999.
- ↑ D.S. Brown. A taxonomic reappraisal of the families Elasmosauridae and Cryptoclididae (Reptilia: Plesiosauroidea). „Révue de Paléobiologie”. 7, s. 9–16, 1993.
- ↑ E.D. Cope. On the reptilian orders Pythonomorpha and Streptosauria. „Proceedings of the Boston Society of Natural History”. 12, s. 265–268, 1869.
- ↑ J. Leidy. April 5th. „Proceedings of the Academy of Natural Sciences of Philadelphia”. 22 (1), s. 18–22, 1870. JSTOR: 4624074.
- ↑ a b c d S.P. Welles. A review of North American Cretaceous elasmosaurs. „University of California Publications in the Geological Sciences”. 29, s. 47–144, 1952.
- ↑ E.D. Cope. The Fossil Reptiles of New Jersey (Continued). „The American Naturalist”. 3 (2), s. 84–91, 1869. DOI: 10.1086/270371. JSTOR: 2447100.
- ↑ E.D. Cope. Additional note on Elasmosaurus. „American Journal of Science”. 50, s. 268–269, 1870.
- ↑ Order – Enaliosauria. W: R. Owen: A History of British Fossil Reptiles. Wyd. 1. London: Cassell & Company Ltd, 1850, s. 215–217.
- ↑ a b c d e f S.P. Welles. A new species of elasmosaur from the Aptian of Columbia and a review of the Cretaceous plesiosaurs. „University of California Publications in the Geological Sciences”. 44, s. 1–96, 1962.
- ↑ S. Sachs, M. Wilmsen, J. Knüppe, J.J. Hornung i inni. Cenomanian–Turonian marine amniote remains from the Saxonian Cretaceous Basin of Germany. „Geological Magazine”. 154 (2), s. 237–246, 2017. DOI: 10.1017/S0016756815001004. Bibcode: 2017GeoM..154..237S.
- ↑ a b c d e f P.O. Persson. A revision of the classification of the Plesiosauria with a synopsis of the stratigraphical and geographical distribution of the group. „Lunds Universitets Arsskrift”. 59 (1), s. 1–59, 1963.
- ↑ N. Bardet, P. Godefroit. Plesiosaurus houzeaui Dollo, 1909 from the Upper Campanian of Ciply (Belgium) and a review of the Upper Cretaceous plesiosaurs from Europe. „Bulletin de l’Institut Royal des Sciences Naturelles de Belgique”. 65, s. 179–186, 1995.
- ↑ E.D. Cope. Report on the Geology of the Region of the Judith River, Montana: And on Vertebrate Fossils Obtained on Or Near the Missouri River. „Bulletin of the United States Geological and Geographical Survey of the Territories”. 3 (3), s. 565–598, 1877.
- ↑ a b c d e f g S.P. Welles. Elasmosaurid plesiosaurs with description of new material from California and Colorado. „Memoir of the University of California”. 13, s. 125–254, 1943.
- ↑ a b c d R.A. Otero. Taxonomic reassessment of Hydralmosaurus as Styxosaurus: new insights on the elasmosaurid neck evolution throughout the Cretaceous. „PeerJ”. 4, s. e1777, 2016. DOI: 10.7717/peerj.1777.
- ↑ Sauropterygia. W: S.W. Williston: Water Reptiles of the Past and Present. Chicago: University of Chicago Press, 1914, s. 86.
- ↑ E.S. Riggs. A specimen of Elasmosaurus serpentinus. „Geological Series of the Field Museum of Natural History”. 6 (25), s. 385–391, 1939. DOI: 10.5962/bhl.title.5289.
- ↑ S. Sachs, J. Lindgren, B.P. Kear: Re-description of Thalassomedon haningtoni – an elasmosaurid from the Cenomanian of North America. W: 5th Triennial Mosasaur Meeting – A Global Perspective on Mesozoic Marine Amniotes. Uppsala: Museum of Evolution, Uppsala University, 2016.
- ↑ a b E.D. Cope. On the Structure of the Skull in the Plesiosaurian Reptilia, and on Two New Species from the Upper Cretaceous. „Proceedings of the American Philosophical Society”. 33 (144), s. 109–113, 1894. JSTOR: 983364.
- ↑ a b c d M.J. Everhart. The occurrence of elasmosaurids (Reptilia: Plesiosauria) in the Niobrara Chalk of Western Kansas. „Paludicola”. 5 (4), s. 170–183, 2006.
- ↑ S.W. Williston. Structure of the Plesiosaurian Skull. „Science”. 16 (405), s. 262, 1890. DOI: 10.1126/science.ns-16.405.262. Bibcode: 1890Sci....16Q.262B.
- ↑ a b G.W. Storrs. An examination of Plesiosauria (Diapsida: Sauropterygia) from the Niobrara Chalk (Upper Cretaceous) of central North America. „University of Kansas Paleontological Contributions”. 11, s. 1–15, 1999.
- ↑ S.W. Williston. North American Plesiosaurs: Trinacromerum. „Journal of Geology”. 16 (8), s. 715–736, 1908. DOI: 10.1086/621573. Bibcode: 1908JG.....16..715W. JSTOR: 30068152.
- ↑ a b c P.O. Persson. Reptiles from the Senonian (U. Cret.) of Scania (S. Sweden). „Arkiv för Mineralogi och Geologi”. 2 (35), s. 431–519, 1959.
- ↑ a b P.A. Pravoslavlev. Геологическое распространенiе эласмозавровъ (Geological distribution of Elasmosaurus). „Bulletin of the Russian Academy of Sciences”. 12 (17), s. 1955–1978, 1918. (ros.).
- ↑ N.N. Bogolubov. Sur la présence de l'Elasmosaurus et du Polycotylus dans les dépots de la Russie (The occurrence of Elasmosaurus and Polycotylus in Russian deposits). „Annuaire Geologique et Minéralogique de la Russie”. 14, s. 174–176, 1912.
- ↑ a b c d Sauropterygia Завроптеригии. W: E. Pervushov, M.S. Arkhangelsky, A.V. Ivanov: Catalog of the locations of the remains of sea reptiles in the Jurassic and Cretaceous of the Lower Volga Region Каталог местонахождений остатков морских рептилий в юр ских и меловых отложениях Нижнего Поволжья. Saratov: Saratov State University Press, 1999, s. 28–34. DOI: 10.13140/RG.2.1.5178.3760.
- ↑ a b c d G.W. Storrs, M.S. Arkhangelsky, V.M. Efimov: Mesozoic marine reptiles of Russia and other former Soviet republics. W: M.J. Benton, M.A. Shishkin, D.M. Unwin, E.N. Kurochkin: The Age of Dinosaurs in Russia and Mongolia. Cambridge: Cambridge University Press, 2000, s. 187–210. ISBN 0-521-55476-4.
- ↑ a b A.O. Averianov, V.K. Popov. The first plesiosaur from the Primorye krai. „Doklady Biological Sciences”. 401 (1), s. 133–135, 2005. DOI: 10.1007/s10630-005-0056-3.
- ↑ Reptiles Fosiles. W: C. Gay: Zoologia, Vol. 2. Paris: Imprenta Maulde y Renou, 1848, s. 130–136, seria: Historia Física y Política de Chile [Physical and Political History of Chile]. (hiszp.).
- ↑ W. Deecke. Ueber Saurierreste aus den Quiriquina–Schichten (Concerning Dinosaur Remains from the Quiriquina Strata). „Neues Jahrbuch für Mineralogie Geologie und Paleontologie”. 14, s. 32–63, 1895. (niem.).
- ↑ E.H. Colbert. A new Cretaceous plesiosaur from Venezuela. „American Museum Novitates”. 1420, s. 1–22, 1949.
- ↑ R.A. Otero, S. Soto-Acuña, D. Rubilar-Rogers. Presence of Mauisaurus in the Maastrichtian (Late Cretaceous) of central Chile. „Acta Palaeontologica Polonica”. 55 (2), s. 361–364, 2010. DOI: 10.4202/app.2009.0065.
- ↑ J.P. O’Gorman, Z. Gasparini, L. Salgado. Postcranial morphology of Aristonectes (Plesiosauria, Elasmosauridae) from the Upper Cretaceous of Patagonia and Antarctica. „Antarctic Science”. 25 (1), s. 71–82, 2013. DOI: 10.1017/S0954102012000673.
- ↑ N. Hiller, J.P. O’Gorman, R.A. Otero, A.A. Mannering. A reappraisal of the Late Cretaceous Weddellian plesiosaur genus Mauisaurus Hector, 1874. „New Zealand Journal of Geology and Geophysics”. 60 (2), s. 112–128, 2017. DOI: 10.1080/00288306.2017.1281317.
- ↑ R. Owen. Notice of some Saurian Fossils discovered by J.H. Hood, Esq., at Waipara, Middle Island, New Zealand. „Geological Magazine”. 7 (68), s. 49–53, 1870. DOI: 10.1017/S0016756800209205. Bibcode: 1870GeoM....7...49O.
- ↑ J. Wiffen, W.L. Moisley. Late Cretaceous reptiles (Families Elasmosauridae and Pliosauridae) from the Mangahouanga Stream, North Island, New Zealand. „New Zealand Journal of Geology and Geophysics”. 29 (2), s. 205–252, 1986. DOI: 10.1080/00288306.1986.10427535.
- ↑ a b S.P. Welles. A new elasmosaur from the Eagle Ford Shale of Texas. „Fondren Science Series”. 1, s. 1–40, 1949.
- ↑ a b S. Sachs, B.P. Kear. Postcranium of the paradigm elasmosaurid plesiosaurian Libonectes morgani (Welles, 1949). „Geological Magazine”. 152 (4), s. 694–710, 2015. DOI: 10.1017/S0016756814000636. Bibcode: 2015GeoM..152..694S.
- ↑ a b K. Carpenter: Comparative cranial anatomy of two North American plesiosaurs. W: J.M. Callaway, E.L. Nicholls: Ancient Marine Reptiles. San Diego: Academic Press, 1997, s. 191–216. DOI: 10.1016/B978-012155210-7/50011-9.
- ↑ A.M. Sørensen, F. Surlyk, J. Lindgren. Food resources and habitat selection of a diverse vertebrate fauna from the upper lower Campanian of the Kristianstad Basin, southern Sweden. „Cretaceous Research”. 42, s. 85–92, 2013. DOI: 10.1016/j.cretres.2013.02.002.
- ↑ E.D. Cope. On the reptilian orders, Pythonomorpha and Streptosauria. „Proceedings of the Boston Society of Natural History”. 12, s. 250–266, 1869.
- ↑ F.R. O’Keefe, A Cladistic Analysis and Taxonomic Revision of the Plesiosauria (Reptilia: Sauropterygia), „Acta Zoologica Fennica”, 213, 2001, s. 1–63, ISBN 951-9481-58-3, ISSN 0001-7299.
- ↑ Introduction. W: C.W. Andrews: A Descriptive Catalogue of the Marine Reptiles of the Oxford Clay. London: British Museum (Natural History), 1910, s. v–xvii. LCCN 11013249.
- ↑ Introduction. W: C.W. Andrews: A Descriptive Catalogue of the Marine Reptiles of the Oxford Clay. London: British Museum (Natural History), 1913, s. v–xvi.
- ↑ The Subclass Synaptosauria. W: W.K. Gregory, S.W. Williston: The Osteology of the Reptiles. Cambridge: Harvard University Press, 1925, s. 246–252. LCCN 25019418.
- ↑ T.E. White. Holotype of Plesiosaurus longirostris Blake and Classification of the Plesiosaurs. „Journal of Paleontology”. 14 (5), s. 451–467, 1940. JSTOR: 1298550.
- ↑ F.R. O’Keefe. Preliminary description and phylogenetic position of a new plesiosaur (Reptilia: Sauropterygia) from the Toarcian of Holzmaden, Germany. „Journal of Paleontology”. 78 (5), s. 973–988, 2004. DOI: 10.1666/0022-3360(2004)078<0973:PDAPPO>2.0.CO;2.
- ↑ a b F. Großman. The taxonomic and phylogenetic position of the Plesiosauroidea from the Lower Jurassic Posidonia Shale of south-west Germany. „Palaeontology”. 50 (3), s. 545–564, 2007. DOI: 10.1111/j.1475-4983.2007.00654.x.
- ↑ a b c H.F. Ketchum, R.B.J. Benson. Global interrelationships of Plesiosauria (Reptilia, Sauropterygia) and the pivotal role of taxon sampling in determining the outcome of phylogenetic analyses. „Biological Reviews”. 85 (2), s. 361–392, 2010. DOI: 10.1111/j.1469-185X.2009.00107.x. PMID: 20002391.
- ↑ a b c R.B.J. Benson, H.F. Ketchum, D. Naish, L.E. Turner. A new leptocleidid (Sauropterygia, Plesiosauria) from the Vectis Formation (Early Barremian–early Aptian; Early Cretaceous) of the Isle of Wight and the evolution of Leptocleididae, a controversial clade. „Journal of Systematic Palaeontology”. 11 (2), s. 233–250, 2013. DOI: 10.1080/14772019.2011.634444.
- ↑ a b R.B.J. Benson, P.S. Druckenmiller. Faunal turnover of marine tetrapods during the Jurassic–Cretaceous transition. „Biological Reviews”. 89 (1), s. 1–23, 2014. DOI: 10.1111/brv.12038. PMID: 23581455.
- ↑ N. Bardet, P. Godefroit, J. Sciau. A new elasmosaurid plesiosaur from the Lower Jurassic of southern France. „Palaeontology”. 42 (5), s. 927–952, 1999. DOI: 10.1111/1475-4983.00103.
- ↑ Z. Gasparini, N. Bardet, J.E. Martin, M.S. Fernandez. The elasmosaurid plesiosaur Aristonectes Cabreta from the Latest Cretaceous of South America and Antarctica. „Journal of Vertebrate Paleontology”. 23 (1), s. 104–115, 2003. DOI: 10.1671/0272-4634(2003)23[104:TEPACF]2.0.CO;2.
- ↑ a b D.J. Serratos, P. Druckenmiller, R.B.J. Benson. A new elasmosaurid (Sauropterygia, Plesiosauria) from the Bearpaw Shale (Late Cretaceous, Maastrichtian) of Montana demonstrates multiple evolutionary reductions of neck length within Elasmosauridae. „Journal of Vertebrate Paleontology”. 37 (2), s. e1278608, 2017. DOI: 10.1080/02724634.2017.1278608.
- ↑ P.S. Druckenmiller, A.P. Russell, A phylogeny of Plesiosauria (Sauropterygia) and its bearing on the systematic status of ''Leptocleidus'' Andrews, 1922, „Zootaxa”, 1863, 2007, s. 1–120, ISBN 978-1-86977-262-8, ISSN 1175-5334.
- ↑ a b c Everhart 2005 ↓, s. 133–135.
- ↑ A. Houssaye. Bone histology of aquatic reptiles: what does it tell us about secondary adaptation to an aquatic life?. „Biological Journal of the Linnean Society”. 108 (1), s. 3–21, 2013-01-01. DOI: 10.1111/j.1095-8312.2012.02002.x. ISSN 0024-4066.
- ↑ Liu, S.; Smith, A.S.; Gu, Y.; Tan, J.; Liu, K.; Turk, G. Computer Simulations Imply Forelimb-Dominated Underwater Flight in Plesiosaurs. „PLoS Computational Biology”. 11 (12), s. e1004605, 2015. DOI: 10.1371/journal.pcbi.1004605. Bibcode: 2015PLSCB..11E4605L.
- ↑ Muscutt, L.E.; Dyke, G.; Weymouth, G.D.; Naish, D.; Palmer, C.; Ganapathisubramani, B. The four-flipper swimming method of plesiosaurs enabled efficient and effective locomotion. „Proceedings of the Royal Society B: Biological Sciences”. 284 (1861), s. 20170951, 2017. DOI: 10.1098/rspb.2017.0951.
- ↑ Everhart 2005 ↓, s. 140.
- ↑ O’Keefe, F.R., Chiappe, L. M. Viviparity and K-selected life history in a Mesozoic marine plesiosaur (Reptilia, Sauropterygia). „Science”. 333 (6044), s. 870–873, 2011. DOI: 10.1126/science.1205689. Bibcode: 2011Sci...333..870O.
- ↑ Everhart 2005 ↓, s. 132–133.
- ↑ a b c d Zammit M., Daniels, C.B., Kear, B. P. Elasmosaur (Reptilia: Sauropterygia) neck flexibility: Implications for feeding strategies. „Comparative Biochemistry and Physiology – Part A: Molecular & Integrative Physiology”. 150 (2), s. 124–130, 2008. DOI: 10.1016/j.cbpa.2007.09.004. PMID: 17933571.
- ↑ Noè, L.F., Taylor, M.A., Gómez-Pérez, M. An integrated approach to understanding the role of the long neck in plesiosaurs. „Acta Palaeontologica Polonica”. 62 (1), s. 137–162, 2017. DOI: 10.4202/app.00334.2016.
- ↑ Everhart, M. An elasmosaur with stomach contents and gastroliths from the Pierre Shale (Late Cretaceous) of Kansas. „Transactions of the Kansas Academy of Science”. 104 (3–4), s. 129–143, 2001. DOI: 10.1660/0022-8443(2001)104[0129:AEWSCA]2.0.CO;2.
- ↑ Wings, O. A review of gastrolith function with implications for fossil vertebrates and a revised classification. „Acta Palaeontologica Polonica”. 52 (1), s. 1–16, 2007.
- ↑ Everhart, M. Bite marks on an elasmosaur (Sauropterygia; Plesiosauria) paddle from the Niobrara Chalk (Upper Cretaceous) as probable evidence of feeding by the lamniform shark, Cretoxyrhina mantelli. „Journal of Vertebrate Paleontology”. 2 (2), s. 14–22, 2005.
- ↑ Thulborn, T.; Turner, S. An elasmosaur bitten by a pliosaur. „Modern Geology”. 18, s. 489–501, 1993.
- ↑ Everhart 2005 ↓, s. 6.
- ↑ a b Steven M. Stanley: Earth System History. New York: W.H. Freeman and Company, 1999, s. 487–489. ISBN 978-0-7167-2882-5.
- ↑ James S. Monroe, Wicander, Reed: The Changing Earth: Exploring Geology and Evolution. Wyd. 5. Belmont, CA: Brooks/Cole, Cengage Learning, 2009, s. 605. ISBN 978-0-495-55480-6.
- ↑ Everhart 2005 ↓, s. 28–38.
- ↑ Everhart 2005 ↓, s. 78, 82–84, 88, 93.
- ↑ Everhart 2005 ↓, s. 58.
- ↑ Everhart 2005 ↓, s. 125, 129, 132–133, 144.
- ↑ Everhart 2005 ↓, s. 108–109.
- ↑ a b Everhart 2005 ↓, s. 221.
- ↑ Everhart 2005 ↓, s. 210.
- ↑ K. Carpenter: Vertebrate Biostratigraphy of the Smoky Hill Chalk (Niobrara Formation) and the Sharon Springs Member (Pierre Shale). W: P.J. Harries: High-resolution Approaches in Stratigraphic Paleontology. Dordrecht: Kluwer Academic Publishers, 2008, s. 421–437, seria: Topics in Geobiology. DOI: 10.1007/978-1-4020-9053-0. ISBN 978-1-4020-9053-0.
Bibliografia
[edytuj | edytuj kod]- Michael Everhart: Oceans of Kansas – A Natural History of the Western Interior Sea. Indiana: Indiana University, 2005. ISBN 978-0-253-34547-9.