Pteranodon
| Pteranodon | |
|---|---|
 | |
| Intervallo geologico | |
| Stato di conservazione | |
Fossile | |
| Classificazione scientifica | |
| Dominio | Eukaryota |
| Regno | Animalia |
| Phylum | Chordata |
| Classe | Sauropsida |
| Ordine | † Pterosauria |
| Sottordine | † Pterodactyloidea |
| Famiglia | † Pteranodontidae |
| Sottofamiglia | † Pteranodontinae Marsh, 1876 |
| Genere | † Pteranodon Marsh, 1876 |
| Nomenclatura binomiale | |
| † Pteranodon longiceps Marsh, 1876 | |
| Sinonimi | |
| |

Pteranodon (il cui nome significa "ala senza denti") è un genere estinto di pterosauro pterodactyloide pteranodontide vissuto nel Cretaceo superiore, circa 86-75 milioni di anni fa (Santoniano-Campaniano), negli attuali Kansas, Alabama, Nebraska, Wyoming e Dakota del Sud, Stati Uniti. La specie tipo del genere, P. longiceps, è uno dei più grandi pterosauri conosciuti, con un'apertura alare di oltre 6 metri. Lo Pteranodon è universalmente lo pterosauro più conosciuto, anche grazie alla notevole raccolta di fossili, superiore a qualsiasi altro pterosauro, con circa 1.200 esemplari conosciuti, molti dei quali erano ben conservati con crani quasi completi e scheletri articolati: la sua abbondanza indica che questo animale era una parte importante nella comunità animale del Mare interno occidentale.[1]
Sebbene venga più volte dipinto nella cultura di massa come un dinosauro, lo Pteranodon era uno pterosauro. Per definizione, tutti i dinosauri appartengono a uno dei due gruppi all'interno di Dinosauria, ossia Saurischia o Ornithischia. Come tale, questo esclude gli pterosauri. Pur non essendo dinosauri, gli pterosauri formano un clade con i dinosauri all'interno del clade Avemetatarsalia.[2]
Descrizione
[modifica | modifica wikitesto]
Le specie di Pteranodon sono estremamente ben rappresentate nei reperti fossili, consentendo descrizioni dettagliate sulla loro anatomia e sulla loro ontogenesi. Attualmente, sono stati identificati oltre 1.000 esemplari, sebbene meno della metà siano abbastanza completi da fornire ai ricercatori informazioni anatomiche valide. Tuttavia, il numero di esemplari raccolti è superiore a qualsiasi altra specie di pterosauro, e comprende sia esemplari maschi che femmine, con varie fasce d'età, e possibilmente diverse specie.[2]
Gli esemplari adulti di Pteranodon delle principali specie possono essere divisi in due classi di dimensioni distinte. La classe dalle dimensioni più piccole sono gli esemplari dalla cresta cranica più piccola ed arrotondata e canali pelvici molto larghi. Le dimensioni del canale pelvico, probabilmente, permetteva a questi individui di deporre le uova, venendo identificati come esemplari femmina. La classe dalle dimensioni più grandi, che rappresentano individui maschi, hanno fianchi stretti e creste craniche molto grandi, che probabilmente venivano impiegate nel riconoscimento intraspecifico e durante il corteggiamento.

Uno Pteranodon maschio adulto era tra i più grandi pterosauri mai vissuti, nonché uno dei più grandi animali volanti conosciuti fino alla fine del XX secolo, quando furono scoperti i giganteschi pterosauri azhdarchidi.[2] L'apertura alare di un maschio adulto medio di Pteranodon era di 5,6 metri. Le femmine adulte erano molto più piccole, con una media di 3,8 metri di apertura alare. Il più grande esemplare di Pteranodon longiceps, rinvenuto all'interno della Formazione Niobrara, misurava 6,25 metri di apertura alare. Un campione ancora più grande, noto dalla Formazione Pierre Shale, possiede un'apertura alare di 7,25 metri, sebbene questo esemplare potrebbe appartenere al genere distinto e alla specie Geosternbergia maysei.[2] Sebbene la maggior parte degli esemplari fossili venga ritrovata schiacciata, esistono abbastanza fossili in tre dimensioni da mettere insieme una descrizione dettagliata dell'animale.
I metodi usati per stimare la massa corporea dei grandi esemplari maschi di Pteranodon (quelli con un'apertura alare di circa 7 metri) sono notoriamente inaffidabili, producendo una vasta gamma di stime da un minimo di 20 chilogrammi fino a 93 chilogrammi. In una revisione sulle stime delle dimensioni degli pterosauri pubblicate nel 2010, i ricercatori Mark Witton e Mike Habib hanno dimostrato che le stime più grandi sono quasi certamente errate dato il volume totale del corpo di Pteranodon, e potrebbero essere corrette solo se l'animale "fosse costituito principalmente da alluminio".[3] Witton e Habib hanno considerato i metodi utilizzati dai ricercatori che hanno ottenuto stime di massa più basse, ma ugualmente difettose. La maggior parte è stata prodotta scalando animali moderni come pipistrelli e uccelli fino alle dimensioni di Pteranodon, nonostante il fatto che gli pterosauri abbiano proporzioni corporee e anatomiche diverse da qualsiasi altro animale vivente.[3]
Altre caratteristiche distintive che distinguono Pteranodon da altri pterosauri sono le sottili spine neurali delle vertebre, i legamenti ossei simili a placche che rafforzano le vertebre sopra l'anca e una coda relativamente lunga. Pteranodon, infatti, possedeva una vertebra caudale "sacralizzata" (ossia facente parte del sinsacro) e 11 o più vertebre caudali libere rese molto particolari dalla presenza di elementi distali formanti una coppia di centro (porzione centrale delle vertebre) fusi e allungati, che si separavano distalmente, formando due "aste caudali" parallele. Inizialmente si pensava che questa struttura fosse una qualche forma di proiezione caudale per le vertebre posteriori, ma sembrano invece essere una serie di piccole vertebre fuse con una scanalatura al centro, apparentemente parte dell'arco neurale.[4] La lunghezza di questa struttura dava a Pteranodon una coda insolitamente lunga per uno pterodactyloide, con una lunghezza della coda di circa il 3,5% dell'apertura alare, o fino a 25 centimetri nei maschi più grandi.[4] Tuttavia, la funzione di questa curiosa struttura rimane poco chiara: Bennett (2001) propose che essa supportasse e controllasse un piccolo uropatagio teso tra le cosce dell'animale, ma ciò è in contrasto con i fossili di pterosauri che suggeriscono che la coda non fosse collegata all'uropatagio. È possibile che Pteranodon fosse l'eccezione alla regola, ma altre funzioni, come ad esempio di supporto per il nuoto, di ancoraggio per strutture di display di tessuti molli o di una banderuola, non possono essere escluse (Witton, 2013).
Cranio e becco
[modifica | modifica wikitesto]
A differenza di pterosauri primitivi, come Rhamphorhynchus e Pterodactylus, lo Pteranodon possedeva un becco sdentato, simile a quello degli uccelli. Il becco dello Pteranodon era fatto di solidi margini ossei che sporgevano dalla base delle mascelle. Il becco era lungo, sottile e terminava in una punta acuminata e sottile. La mascella, che era più lunga della mandibola, era curvata verso l'alto; mentre questo normalmente è stato attribuito solo al becco che si piega verso l'alto, un campione (UALVP 24238) ha una curvatura corrispondente all'ampiezza del becco verso la punta. Mentre la punta del becco non è conosciuta in questo esemplare, il livello di curvatura suggerisce che il becco di questo esemplare doveva essere estremamente lungo. La forma unica del becco in questo esemplare ha portato Alexander Kellner ad assegnarlo a un genere distinto, Dawndraco, nel 2010.[5] Tuttavia, successivi studi di Elizabeth Martin-Silverstone et al. (2017) hanno dimostrato che le differenze nel becco tra i vari individui probabilmente erano una caratteristica comune negli pteranodontidi. Pertanto conclusero che "Dawndraco" era semplicemente un maschio di Geosternbergia sternbergi.[6]
Inoltre molti ricercatori ritengono che, con ogni probabilità, lo Pteranodon possedesse una piccola sacca sotto il becco, analoga nella funzione a quella del pellicano oggi, e certe impronte fossili sembrerebbero indicarlo.[7]
La caratteristica più distintiva di Pteranodon è la sua voluminosa cresta cranica. Queste creste erano costituite dall'osso frontale che sporgevano verso il retro del cranio e verso l'alto. Le dimensioni e la forma di queste creste varia in base ad una serie di fattori, tra cui età, sesso e specie. I maschi di Pteranodon sternbergi, la specie più antica descritta fino ad oggi (e talvolta collocata nel genere distinto Geosternbergia), possedeva una cresta verticale con un'ampia proiezione in avanti, mentre i loro discendenti, Pteranodon longiceps, possedevano una cresta più stretta, e arretrata nel capo.[1] Le femmine di entrambe le specie avevano invece creste piccole e arrotondate.[8] Le creste erano probabilmente strutture espositive e comunicative, sebbene potessero avere anche altre funzioni.[9] Si ritiene infatti che gli Pteranodon potessero utilizzare la cresta cranica per cambiare posizione durante il volo come una sorta di timone.
Classificazione
[modifica | modifica wikitesto]Evoluzione
[modifica | modifica wikitesto]
La maggior parte dei fossili di Pteranodon provengono dalla Formazione Niobrara, degli Stati Uniti centrali. Lo Pteranodon visse per più di 4 milioni di anni, tra il tardo Coniaciano e i primi del Campaniano, nel Cretaceo superiore.[8] Il genere è presente nella maggior parte degli strati della Formazione Niobrara ad eccezione dei due superiori; nel 2003 Kenneth Carpenter ha esaminato la distribuzione e la datazione dei fossili in questa formazione, dimostrando che la presunta specie P. sternbergi visse tra 88-85 milioni di anni fa, mentre la successiva specie tipo P. longiceps visse tra 86-84,5 milioni di anni fa. Una possibile terza specie, che Kellner ha nominato Geosternbergia maysei, nel 2010, è stata ritrovata nelle Sharon Springs della Formazione Pierre Shale, in Kansas, Wyoming e Dakota del Sud, e sembrerebbe essere vissuto tra 81,5-80,5 milioni di anni fa.[10]
All'inizio degli anni '90, Bennett notò che le due principali forme di Pteranodon presenti nella Formazione Niobrara erano precisamente separati nel tempo con poche, se non nessuna, sovrapposizioni. A causa di ciò, e alla loro grossolana somiglianza generale, Bennett suggerì che le due forme rappresentassero delle cronospecie all'interno di un unico lignaggio evolutivo della durata di circa 4 milioni di anni. In altre parole, solo una specie di Pteranodon sarebbe stata presente in un determinato periodo, e P. sternbergi (o Geosternbergia) con ogni probabilità era l'antenato diretto di P. longiceps.[2]
Specie valide
[modifica | modifica wikitesto]
Molti ricercatori affermano che ci siano almeno due specie di Pteranodon. Tuttavia, a parte le differenze tra maschi e femmine descritte sopra, gli scheletri post-cranici di Pteranodon mostrano poca o nessuna variazione tra specie o esemplari, e il corpo e le ali di tutti gli Pteranodon erano essenzialmente identici.[8]
Tradizionalmente, sono riconosciute come valide due specie di Pteranodon: la specie tipo Pteranodon longiceps, e Pteranodon sternbergi. Le specie due differiscono solo nella forma della cresta nei maschi adulti (descritte sopra) e possibilmente nell'angolo di alcune ossa del cranio.[8] Poiché i fossili del cranio di Pteranodon ben conservati sono estremamente rari, i ricercatori usano la stratigrafia (cioè quale strato di roccia della formazione geologica in cui si trova un fossile) per determinare l'identità delle specie nella maggior parte dei casi.
Pteranodon sternbergi è l'unica specie conosciuta di Pteranodon con una cresta verticale. La mascella inferiore di P. sternbergi era lunga 1,25 metri.[11] La specie venne recuperata da George F. Sternberg, nel 1952 e descritta da John Christian Harksen, nel 1966, dalla parte inferiore della Formazione Niobrara. La specie è più antica di P. longiceps ed è considerata da Bennett l'antenato diretto delle specie successive.[8]
Poiché i fossili identificabili come P. sternbergi si trovano esclusivamente negli strati inferiori della Formazione Niobrara, e i fossili di P. longiceps si trovano esclusivamente negli strati superiori, un fossile privo di cranio può essere identificato in base alla sua posizione nella colonna geologica (sebbene per molti ritrovamenti di fossili precoci, dati precisi sulla loro posizione non sono stati registrati, rendendo molti fossili non identificabili).[5]
Di seguito è riportato un cladogramma che mostra i risultati di un'analisi filogenetica presentata per la prima volta da Andres e Myers (2013)[12], e aggiornata con dati aggiuntivi di Andres et al. (2018):[13]
| Ornithocheiroidea |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Classificazioni alternative
[modifica | modifica wikitesto]
A causa delle sottili variazioni tra gli esemplari di pteranodontidae all'interno della Formazione Niobrara, molti ricercatori hanno assegnato tutti i materiali ritrovati al genere Pteranodon, in almeno due specie (P. longiceps e P. sternbergi) che si distinguono principalmente per la forma della cresta. Tuttavia, la classificazione di queste due forme varia da ricercatore a ricercatore. Nel 1972 Halsey Wilkinson Miller pubblicò un articolo in cui sosteneva che le varie forme di Pteranodon erano abbastanza diverse tra loro da essere collocate in sottogeneri distinti. Egli nominò questi esemplari come Pteranodon (Occidentalia) occidentalis (per le specie ora in disuso P. occidentalis) e Pteranodon (Sternbergia) sternbergi. Tuttavia, il nome Sternbergia era già in uso, e nel 1978 Miller rinominò la specie Pteranodon (Geosternbergia) sternbergi, nominando una terza combinazione sottogenere/specie per P. longiceps, come Pteranodon (Longicepia) longiceps. Tuttavia, i più importanti ricercatori di pterosauri del tardo XX secolo, tra cui S. Christopher Bennett e Peter Wellnhofer, non hanno adottato questi nomi subgenerici e hanno continuato a collocare tutti gli esemplari di pteranodontide nel singolo genere Pteranodon.
Nel 2010, il ricercatore di pterosauri Alexander Kellner ha rivisitato la classificazione di H.W. Miller. Kellner, seguendo le idee di Miller secondo cui le differenze tra le specie di Pteranodon erano abbastanza da poterli classificare in generi diversi, collocò P. sternbergi nel genere nominato da Miller, Geosternbergia, insieme all'esemplare del cranio di Pierre Shale che Bennett aveva precedentemente considerato un grande P. longiceps di sesso maschile. Kellner sosteneva che la cresta di questo esemplare, sebbene non completamente conservata, fosse molto simile a quella di Geosternbergia. Poiché l'esemplare era di milioni di anni più giovane di qualsiasi Geosternbergia conosciuto, Kellner lo assegnò alla nuova specie Geosternbergia maysei. Numerosi altri esemplari di pteranodonti sono noti nella stessa formazione, e Kellner ha suggerito che potrebbero appartenere alla stessa specie di G. maysei, ma poiché sono privi di teschi, non è stato in grado di identificarli con sicurezza.[5]
Specie rinnegate
[modifica | modifica wikitesto]
Dal 1870 sono state nominate diverse specie di Pteranodon, anche se la maggior parte sono ora considerate sinonimi junior di due o tre specie valide. La specie meglio conosciuto è la specie tipo, P. longiceps, basata su un esemplare ben conservato incluso il primo cranio conosciuto di S. W. Williston. Questo individuo aveva un'apertura alare di 7 metri (23 piedi).[14] Altre specie valide includono la possibile specie P. sternbergi, con un'apertura alare originariamente stimata di 9 metri.[14] Le specie P. occidentalis, P. velox, P. umbrosus, P. harpyia e P. comptus sono considerati nomina dubia da Bennett (1994) e altri studiosi ne mettono in dubbio la validità. Probabilmente rappresentano tutti sinonimi delle specie più conosciute.
Poiché la caratteristica distintiva fondamentale che Marsh notò in Pteranodon era la sua mancanza di denti, qualsiasi frammento di mascella di pterosauro sdentato, ovunque fosse stato ritrovato nel mondo, tendeva ad essere attribuito a Pteranodon, durante la fine del diciannovesimo secolo e all'inizio del ventesimo. Ciò portò alla denominazione di una pletora di nuove specie e ad una grande confusione tassonomica. Il nome Pteranodon divenne un refugium peccatorum, un po' come successe per il dinosauro Megalosaurus, per etichettare tutti i resti di pterosauri che non potevano essere distinti se non dall'assenza di denti. Le specie (spesso di dubbia fama che si basano su variazioni sessuali o caratteri giovanili) sono state riclassificate un certo numero di volte, e diversi sottogeneri sono stati eretti negli anni '70 da Halsey Wilkinson Miller per tenerli in varie combinazioni, confondendo ulteriormente la tassonomia (i sottogeneri includono Longicepia, Occidentalia e Geosternbergia). Tra gli autori di spicco che hanno discusso i vari aspetti di Pteranodon figurano Bennett, Padian, Unwin, Kellner e Wellnhofer. Due specie, P. orogensis e P. orientalis, non erano neanche pteranodontidi e sono state rinominate rispettivamente Bennettazhia oregonensis e Bogolubovia orientalis.
Elenco delle specie e dei sinonimi
[modifica | modifica wikitesto]Lo stato dei nomi elencati di seguito è basato su un sondaggio di Bennett, 1994, se non diversamente indicato.[8]
| Nome | Autore | Anni | Stato | Note |
|---|---|---|---|---|
| Pterodactylus occidentalis | Marsh | 1872 | Riclassificato come Pteranodon occidentalis | Riclassificato come Pterodactylus oweni Marsh 1871 (pre-occupato da Seeley 1864) |
| Pterodactylus ingens | Marsh | 1872 | Riclassificato come Pteranodon ingens | |
| Pterodactylus velox | Marsh | 1872 | Nomen dubium | Riclassificato come Pteranodon velox |
| Ornithochirus umbrosus | Cope | 1872 | Nomen dubium | |
| Ornithochirus harpyia | Cope | 1872 | Nomen dubium | |
| Pterodactylus umbrosus | (Cope) Cope | (1872) 1874 | Riclassificato come Ornithochirus umbrosus | |
| Pteranodon longiceps | Marsh | 1876 | Valida | Specie tipo |
| Pteranodon ingens | (Marsh) Williston | (1872) 1876 | Nomen dubium | Riclassificato come Pterodactylus ingens |
| Pteranodon occidentalis | Marsh | (1872) 1876 | Nomen dubium | Riclassificato da Pterodactylus occidentalis |
| Pteranodon velox | Marsh | (1872) 1876 | Nomen dubium | Riclassificato da Pterodactylus velox, basato su un esemplare giovane |
| Pteranodon gracilis | Marsh | 1876 | Riclassificato come Nyctosaurus gracilis | |
| Pteranodon comptus | Marsh | 1876 | Nomen dubium | |
| Pteranodon nanus | Marsh | 1876 | Riclassificato come Nyctosaurus nanus | |
| Ornithocheirus umbrosus | (Cope) Newton | (1872) 1888 | Riclassificato come Pteranodon umbrosus | Correzione ortografica di Ornithochirus umbrosus |
| Ornithocheirus harpyia | (Cope) Newton | (1872) 1888 | Riclassificato come Pteranodon harpyia | Correzione ortografica di Ornithochirus harpyia |
| Pteranodon umbrosus | (Cope) Williston | (1872) 1892 | Nomen dubium | Riclassificato come Ornithochirus umbrosus |
| Ornithostoma ingens | (Marsh) Williston | (1872) 1893 | Sinonimo di Pteranodon ingens | Riclassificato da Pteranodon ingens |
| Ornithostoma umbrosum | (Cope) Williston | (1872) 1897 | Sinonimo di Pteranodon umbrosus | Riclassificato da Pteranodon umbrosus |
| Pteranodon oregonensis | Gilmore | 1928 | Riclassificato come Bennettazhia oregonensis | |
| Pteranodon sternbergi | Harksen | 1966 | Valida | |
| Pteranodon marshi | Miller | 1972 | Sinonimo di Pteranodon longiceps | |
| Pteranodon bonneri | Miller | 1972 | Riclassificato come Nyctosaurus bonneri | |
| Pteranodon walkeri | Miller | 1972 | Sinonimo di Pteranodon longiceps | |
| Pteranodon (Occidentalia) eatoni | (Miller) Miller | (1972) 1972 | Sinonimo di Pteranodon sternbergi | |
| Pteranodon eatoni | (Miller) Miller | (1972) 1972 | Sinonimo di Pteranodon sternbergi | Riclassificato da Pteranodon (Occidentalia) eatoni |
| Pteranodon (Longicepia) longicps | (Marsh) Miller | (1872) 1972 | Sinonimo di Pteranodon longiceps | Riclassificato da Pteranodon longiceps |
| Pteranodon (Longicepia) marshi | (Miller) Miller | (1972) 1972 | Sinonimo di Pteranodon longiceps | Riclassificato da Pteranodon marshi |
| Pteranodon (Sternbergia) sternbergi | (Harksen) Miller | (1966) 1972 | Riclassificato come Pteranodon (Geosternbergia) sternbergi | Riclassificato da Pteranodon sternbergi |
| Pteranodon (Sternbergia) walkeri | (Miller) Miller | (1972) 1972 | Riclassificato come Pteranodon (Geosternbergia) walkeri | Riclassificato da Pteranodon walkeri |
| Pteranodon (Pteranodon) marshi | (Miller) Miller | (1972) 1973 | Sinonimo di Pteranodon longiceps | Riclassificato da Pteranodon marshi |
| Pteranodon (Occidentalia) occidentalis | (Marsh) Olshevsky | (1872) 1978 | Sinonimo di Pteranodon occidentalis | Riclassificato da Pteranodon occidentalis |
| Pteranodon (Longicepia) ingens | (Marsh) Olshevsky | (1872) 1978 | Sinonimo di Pteranodon ingens | Riclassificato da Pteranodon ingens |
| Pteranodon (Pteranodon) ingens | (Marsh) Olshevsky | (1872) 1978 | Sinonimo di Pteranodon ingens | Riclassificato da Pteranodon ingens |
| Pteranodon (Geosternbergia) walkeri | (Miller) Miller | (1972) 1978 | Sinonimo di Pteranodon longiceps | Riclassificato da Pteranodon walkeri |
| Pteranodon (Geosternbergia) sternbergi | (Harksen) Miller | (1966) 1978 | Sinonimo di Pteranodon sternbergi | Riclassificato da Pteranodon (Sternbergia) sternbergi |
| Pteranodon orientalis | (Bogolubov) Nesov & Yarkov | (1914) 1989 | Riclassificato come Bogolubovia orientalis | Riclassificato da Ornithostoma orientalis |
| Geosternbergia walkeri | (Miller) Olshevsky | (1872) 1991 | Sinonimo di Pteranodon sternbergi | Riclassificato da Pteranodon (Sternbergia) walkeri |
| Geosternbergia sternbergi | (Harksen) Olshevsky | (1966) 1991 | Sinonimo di Pteranodon sternbergi | Riclassificato da Pteranodon (Geosternbergia) sternbergi |
Storia della scoperta
[modifica | modifica wikitesto]
Primi ritrovamenti
[modifica | modifica wikitesto]Pteranodon fu il primo pterosauro ritrovato al di fuori dell'Europa. I suoi fossili furono scoperti per la prima volta da Othniel Charles Marsh nel 1870, nei depositi di calcare risalenti al Cretaceo superiore, delle Smoky Hill nel Kansas occidentale. Questi letti di gesso si depositarono sul fondo di quello che un tempo era il Mare Interno Occidentale, un grande mare poco profondo che si estendeva in quella che oggi è la parte centrale del continente nordamericano. Questi primi esemplari, YPM 1160 e YPM 1161, consistevano in ossa parziali delle ali, nonché di alcuni denti del pesce preistorico Xiphactinus, che Marsh assegnò erroneamente a questo nuovo pterosauro (tutti gli pterosauri noti fino a quel momento avevano i denti). Nel 1871 Marsh nominò il ritrovamento "Pterodactylus oweni", assegnandolo al noto (ma molto più piccolo) genere europeo Pterodactylus. Marsh raccolse nuove ossa delle ali di questo grande pterosauro. Rendendosi conto che il nome che aveva scelto era già stato usato per un'altra specie europea di pterosauro, Pterodactylus oweni, nominato da Harry Seeley nel 1864, Marsh ribattezzò il suo gigantesco pterosauro nordamericano come Pterodactylus occidentalis, il nome significava "dito alato occidentale", nella sua descrizione del nuovo esemplare del 1872. Inoltre nominò altre due specie, sulla base delle differenze nelle dimensioni: Pterodactylus ingens (il più grande esemplare raccolto finora) e Pterodactylus velox (il più piccolo).[8]
Nel frattempo, anche il rivale di Marsh, Edward Drinker Cope, aveva portato alla luce diversi esemplari del grande pterosauro nordamericano. Sulla base di questi esemplari, Cope nominò due nuove specie, Ornithochirus umbrosus e Ornithochirus harpyia, nel tentativo di assegnarle al grande genere europeo Ornithocheirus, sebbene avesse scritto male il nome (dimenticando la 'e').[8] Il documento di Cope che nomina la sua specie fu pubblicato nel 1872, appena cinque giorni dopo il documento di Marsh. Ciò provocò una disputa, combattuta nella letteratura pubblicata, su quale nome avesse la priorità su quelle che ovviamente erano la stessa specie.[8] Cope ammise nel 1875 che i nomi di Marsh avevano priorità sui suoi, ma sostenne che Pterodactylus umbrosus fosse una specie distinta (ma non un genere) da quella che Marsh aveva precedentemente nominato.[15] La rivalutazione da parte di scienziati successivi ha sostenuto il caso di Marsh e ha scoperto che l'affermazione di Cope secondo cui P. umbrosus era una specie più grande e distinta era errata.[8]
Uno pterosauro sdentato
[modifica | modifica wikitesto]
Mentre le prime ossa delle ali di Pteranodon furono raccolte da Marsh e Cope nei primi del 1870, il primo cranio di Pteranodon venne ritrovato il 2 maggio 1876, lungo il fiume Smoky Hill, nella Contea di Wallace (ora Contea di Logan), Kansas, USA, da Samuel Wendell Williston, un collezionista di fossili che lavorava per Marsh.[1] Ben presto venne scoperto un secondo cranio più piccolo. Questi crani dimostrarono che gli pterosauri nordamericani erano diversi da qualsiasi specie europea, in quanto non possedevano denti e presentavano grandi creste ossee sul capo. Marsh riconobbe questa grande differenza, descrivendo gli esemplari come "distinti da tutti i generi precedentemente noti dell'ordine Pterosauria per l'intera assenza di denti". Marsh riconobbe che questa caratteristica garantiva un nuovo genere, e così coniò il nome Pteranodon (che significa per l'appunto "ala senza dente") nel 1876. Marsh riclassificò tutte le specie nordamericane precedentemente nominate come Pterodactylus in Pteranodon. Marsh assegnò il cranio più piccolo alla specie Pteranodon occidentalis, in base alle dimensioni, e assegnò il cranio più grande, YPM 1117, alla nuova specie Pteranodon longiceps, che pensava fosse una specie di medie dimensioni tra il piccolo P. occidentalis e il grande P. ingens.[8][16] Marsh nominò diverse altre specie: Pteranodon comptus e Pteranodon nanus sono stati nominati sulla base di scheletri frammentari di piccoli individui, mentre Pteranodon gracilis si basava su un osso dell'ala che aveva scambiato per un osso pelvico. Presto si rese conto del suo errore e riclassificò di nuovo quell'esemplare in un genere separato, che chiamò Nyctosaurus. P. nanus in seguito fu anch'esso riconosciuto come un esemplare di Nyctosaurus.[8][17]
Nel 1892 Samuel Williston esaminò la questione della classificazione di Pteranodon. Notò che, nel 1871, Seeley aveva menzionato l'esistenza di una serie parziale di mascelle di pterosauro sdentato dalla Cambridge Greensand d'Inghilterra, che chiamò "Ornithostoma". Poiché la caratteristica principale che Marsh aveva usato per separare Pteranodon dagli altri pterosauri era la sua mancanza di denti, Williston concluse che "Ornithostoma" doveva essere considerato il sinonimo senior di Pteranodon. Tuttavia, nel 1901, Pleininger fece notare che "Ornithostoma" non era mai stato descritto scientificamente né aveva nemmeno assegnato un nome specifico fino al lavoro di Williston, e quindi era stato un nomen nudum e non poteva battere Pteranodon per priorità di denominazione. Williston accettò questa conclusione e tornò a chiamare il genere Pteranodon.[8] Tuttavia, sia Williston che Pleininger erano errati, poiché inosservato di entrambi era il fatto che, nel 1891, lo stesso Seeley aveva finalmente descritto e correttamente nominato Ornithostoma, assegnandolo alla specie O. sedgwicki. Nel 2010, ulteriori ricerche sull'identità di Ornithostoma hanno dimostrato che probabilmente Ornithostoma non era uno Pteranodon e non era neppure un suo parente stretto, ma potrebbe essere stato in realtà un azhdarchoide, un diverso tipo di pterosauro sdentato.[18]
Revisione delle specie
[modifica | modifica wikitesto]Williston è stato anche il primo scienziato a valutare criticamente tutte le specie di Pteranodon classificate da Cope e Marsh. Williston concordò con la maggior parte della classificazione di Marsh, con alcune eccezioni. In primo luogo, non credeva che P. ingens e P. umbrosus potessero essere considerati sinonimi, cosa che persino Cope credeva. Considerava specie dubbie sia P. velox che P. longiceps; il primo era basato su frammenti non diagnostici e il secondo, sebbene noto da un cranio completo, probabilmente apparteneva a un'altra delle specie precedentemente nominate. Nel 1903 Williston rivisitò la questione della classificazione di Pteranodon, e rivise la sua precedente conclusione, portando il numero di specie da sette a tre. Considerò sia P. comptus che P. nanus esemplari di Nyctosaurus, e divise gli altri in specie piccole (P. velox), medie (P. occidentalis) e grandi (P. ingens), basate principalmente sulla forma delle ossa del braccio. Pensava che P. longiceps, l'unic< specie conosciuta da un cranio, potesse essere sinonimo di P. velox o P. occidentalis, in base alle dimensioni.[8]
Nel 1910 Eaton divenne il primo scienziato a pubblicare una descrizione dettagliata dell'intero scheletro di Pteranodon, come era noto all'epoca. Eaton usò i suoi risultati per rivedere la classificazione del genere ancora una volta sulla base di una migliore comprensione delle differenze nell'anatomia di Pteranodon, conducendo esperimenti usando modelli di ossa di argilla per aiutarsi a determinare gli effetti di schiacciamento e appiattimento sulle forme delle ossa del braccio che Williston aveva usato nella sua stessa classificazione. Eaton scoprì che la maggior parte delle differenze nelle forme ossee potrebbe essere facilmente spiegata dalle pressioni della fossilizzazione e concluso che nessuno degli scheletri di Pteranodon mostrava differenze significative tra loro oltre alle dimensioni. Pertanto, Eaton ideò un nuovo schema di classificazione che si basava sulle sole differenze dei crani, che assegnò alle varie specie, proprio come fece Marsh in base alle dimensioni. Alla fine, Eaton riconobbe solo tre specie valide: P. occidentalis, P. ingens e P. longiceps.[8]
La scoperta di esemplari con creste verticali, classificate da Harksen nel 1966 come la nuova specie Pteranodon sternbergi, complicò ulteriormente la situazione, spingendo un'altra revisione del genere da parte di Miller nel 1972. Perché era impossibile determinare la forma della cresta per tutte le specie sulla base di scheletri privi di cranio, Miller concluse che tutte le specie di Pteranodon, tranne le due basate su teschi (P. longiceps e P. sternbergi), dovevano essere considerate nomena dubia e abbandonate. Il cranio che Eaton pensava appartenesse a P. ingens fu collocato nella nuova specie Pteranodon marshi, e il cranio che Eaton aveva assegnato a P. occidentalis fu rinominato Pteranodon eatoni. Miller riconobbe anche un'altra specie basata su un cranio con una cresta simile a quella di P. sternbergi; Miller chiamò questa specie Pteranodon walkeri. Per aiutare a mettere ordine in questo groviglio di nomi, Miller creò tre categorie o "sottogeneri" per loro. P. marshi e P. longiceps sono stati collocati nel sottogenere Longicepia, anche se questo è stato successivamente cambiato semplicemente in Pteranodon a causa delle regole di priorità. P. sternbergi e P. walkeri, le specie a cresta verticale, ricevettero il sottogenere Sternbergia, che in seguito fu cambiato in Geosternbergia perché Sternbergia era un nome già in uso ("preoccupato"). Infine, Miller nominò il sottogenere Occidentalia per P. eatoni, il cranio precedentemente associato a P. occidentalis. Miller ha ulteriormente ampliato il concetto di Pteranodon per includere Nyctosaurus come quarto sottogenere. Miller li considerava una progressione evolutiva, con il primitivo Nyctosaurus, all'epoca ritenuto un genere senza cresta, che aveva dato origine agli Occidentalia (dalla cresta più piccola), che a sua volta diede origine a Pteranodon con la sua lunga cresta all'indietro, portando infine a Geosternbergia con la sua grande cresta verticale. Tuttavia, Miller fece diversi errori nel suo studio riguardo a quali esemplari Marsh aveva assegnato a quali specie, e la maggior parte degli scienziati ignorò il suo lavoro sull'argomento nelle loro ricerche successive, sebbene Wellnhofer (1978) seguì l'elenco delle specie di Miller, e Schoch (1984) pubblicò in qualche modo un'altra revisione che essenzialmente è tornata allo schema di classificazione originale di Marsh, in particolare affondando P. longiceps come sinonimo di P. ingens.[8]
Riconoscimento delle variazioni
[modifica | modifica wikitesto]All'inizio degli anni '90, S. Christopher Bennett pubblicò diversi importanti articoli che analizzavano l'anatomia, la tassonomia e la storia della vita di Pteranodon.[5]
Fossili frammentari assegnati a Pteranodon sono stati scoperti anche a Scania, in Svezia.[19]
Paleobiologia
[modifica | modifica wikitesto]Volo
[modifica | modifica wikitesto]
La forma dell'ala dello Pteranodon suggerisce che questo animale avesse uno stile di volo simile a quello di un moderno albatros. Ciò si basa sul fatto che lo Pteranodon aveva un rapporto di aspetto elevato (apertura alare per la lunghezza della corda alare) molto simile a quello di un albatros, circa 9:1 per lo Pteranodon, rispetto all'8:1 dell'albatros. Gli albatros trascorrono lunghi periodi in mare per pescare ed usano un modello di volo chiamato "veleggiamento dinamico" che sfrutta il gradiente verticale della velocità del vento vicino alla superficie dell'oceano per percorrere lunghe distanze senza battere le ali, e senza l'ausilio di correnti termiche, assenti in oceano aperto.[20] Anche se gran parte del volo dell'animale si sarebbe basato sul veleggiamento, come avviene per gli uccelli marini con ali lunghe, il volo dello Pteranodon richiedeva comunque un occasionale battito d'ali attivo, e studi sul carico alare (la forza delle ali rispetto al peso del corpo) indicano che l'animale era in grado di volare in modo sostanziale sbattendo attivamente le ali, contrariamente ad alcune ipotesi precedenti secondo cui l'animale era così grande che avrebbe potuto solamente planare.[21]
Come molti altri pterosauri, Pteranodon, probabilmente, decollava in una posizione quadrupede, usando i loro lunghi arti anteriori come leva, per lanciarsi in aria con un balzo in avanti seguito da un rapido battito d'ali per prendere quota. Quasi tutta l'energia sarebbe stata generata dagli arti anteriori. Le ali si sarebbero spiegate solo quando l'animale si fosse sollevato abbastanza dal terreno e avesse completato il lancio in aria.[21]
Locomozione terrestre
[modifica | modifica wikitesto]
Storicamente, la discussione sulla locomozione terrestre di Pteranodon, in particolare se l'animale fosse bipede o quadrupede, così come la locomozione terrestre dagli pterosauri in generale, è stato oggetto di dibattito.
Attualmente la maggior parte dei ricercatori sostiene che gli pterosauri fossero animali quadrupedi, spostandosi utilizzando tutti e quattro gli arti, e ripiegando le ali dietro il corpo, durante la locomozione a terra. Ciò è stato confermato anche dal ritrovamento di piste di impronte attribuite a pterosauri.[22]
La possibilità di una locomozione acquatica è stata discussa in diverse pubblicazioni, in particolare negli studi effettuati dal paleontologo Christopher Bennett (2001), che evidenziarono la netta somiglianza degli arti posteriori di Pteranodon con quelli dell'ibis rosso (somiglianze trovate nelle dita I, III e IV) e dei gabbiani (somiglianze trovate nelle dita II e IV).[23]
Al contrario gli arti di Pteranodon risultavano diversi nella morfologia da quelli di altre specie prese in esame, come quella del pavone, dell'estinta alca impenne e del coccodrillo marino.[23]
Bennett ritenne fortemente probabile che lo Pteranodon fosse capace di nuotare e che la sua struttura fisica fosse idonea perfino a brevi immersioni subacquee, in modo analogo allo stile di caccia delle moderne sule e pellicani.[24].
Dieta e caccia
[modifica | modifica wikitesto]
È noto che la dieta di Pteranodon fosse costituita in gran parte da pesce; diverse ossa di pesce fossilizzate sono state ritrovate nella regione dello stomaco di uno Pteranodon, e un bolo di pesce fossilizzato è ristato trovato tra le fauci dell'esemplare AMNH 5098. Numerosi altri esemplari conservano frammenti di squame e vertebre nella zona del torso, indicando che il pesce costituiva la maggior parte della dieta di questo animale, sebbene sia possibile si possa essere cibato anche di molluschi o altri vertebrati marini[25]
Tradizionalmente, la maggior parte dei ricercatori ipotizzava che lo Pteranodon catturasse il pesce immergendo il becco in acqua mentre era in volo radente sul mare. Tuttavia, questa si ipotesi si basato sul presupposto che questi animali non potessero decollare dalla superficie dell'acqua.[26] Tuttavia, è molto probabile che Pteranodon fosse in grado di decollare dall'acqua, e che potesse immergersi per pescare. Anche un piccolo Pteranodon femmina avrebbe potuto raggiungere una profondità di almeno 80 centimetri (31 pollici) con il becco e il collo mentre il resto del corpo galleggiava sulla superficie, e potrebbe aver raggiunto profondità anche maggiori tuffandosi in acqua dall'aria come alcuni moderni uccelli marini dalle ali lunghe.[25] Nel 1994 Bennett evidenziò che la testa, il collo e le spalle di Pteranodon avevano una costituzione robusta, proprio come molti uccelli subacquei. Ciò suggerisce che l'animale era in grado di tuffarsi da grandi altezze e raggiungere anche grandi profondità, ripiegando le ali come le moderne sule.[24]
Funzione della cresta
[modifica | modifica wikitesto]
Una delle caratteristiche più note di Pteranodon è la sua iconica cresta cranica, sebbene la sua funzione sia ancora oggetto di dibattito. La maggior parte dei dibattiti si è concentrata sulla forma a lama della cresta, con la punta all'indietro nei maschi di P. longiceps, ignorando però l'ampia gamma di variazioni individuali di età e sesso. Il fatto che le creste varino così tanto tra individui della stessa specie esclude la maggior parte delle funzioni pratiche diverse da quelle utilizzate in display d'accoppiamento.[27] Pertanto, la funzione principale della cresta era probabilmente quella di display per attrarre il partner, mentre tutte le altre funzioni erano secondarie.[9]
Le possibili interpretazioni scientifiche della funzione della cresta iniziarono nel 1910, quando George Francis Eaton propose due possibilità: un contrappeso aerodinamico e/o un punto di attacco muscolare. Eaton ipotizzò che la cresta potesse essere un punto di ancoraggio per grandi e lunghi muscoli mascellari, ma ammise che questa funzione da sola non poteva spiegare il perché delle grandi dimensioni di alcune creste.[28] Bennett (1992) concordò con la stessa valutazione di Eaton che la cresta era troppo grande e variabile per essere un sito di attacco muscolare.[9] Eaton aveva ipotizzato che una possibile funzione secondaria della cresta era di contrappeso al lungo becco, riducendo la necessità di pesanti muscoli del collo per controllare l'orientamento della testa.[28] Alcuni test effettuati nella galleria del vento hanno dimostrato che la cresta funzionava effettivamente in qualche modo come un efficace contrappeso, ma Bennett notò che, ancora una volta, l'ipotesi si concentra solo sulle lunghe creste dei maschi di P. longiceps, e non sulle più grandi creste di P. sternbergi o su quelle più piccole delle femmine. Bennett scoprì che le creste delle femmine non avevano alcun effetto di controbilanciamento e che le creste dei maschio di P. sternbergi avrebbero, da sole, avuto un effetto negativo sull'equilibrio della testa. In effetti, il movimento laterale delle creste avrebbe richiesto un'elevata muscolatura del collo per controllare l'equilibrio della testa.[9]
Nel 1943 Dominik von Kripp ipotizzò che le creste avrebbero potuto avere la funzione di timone, un'idea abbracciata da numerosi ricercatori successivi.[9][29] Ross S. Stein ha persino suggerito che la cresta potesse essere sostenuta da una membrana di pelle che collegava la cresta rivolta all'indietro al collo e alla schiena, aumentando la sua superficie ed efficacia come un timone.[30] L'ipotesi del timone, ancora una volta, non tiene conto degli individui femminili né di P. sternbergi, che aveva una cresta rivolta verso l'alto, non rivolta all'indietro. Bennett scoprì anche che, anche nella sua qualità di timone, la cresta non avrebbe fornito la stessa forza direzionale della semplice manovra delle ali. L'ipotesi che la cresta fosse un freno ad aria e che gli animali volgessero la testa di lato per rallentare, soffre di un problema simile.[31] Inoltre, le ipotesi del timone e del freno ad aria non spiegano perché esista una variazione così grande nella dimensione della cresta anche tra gli adulti.[9]
Alexander Kellner suggerì che le grandi creste dello pterosauro Tupandactylus, così come altre specie, potrebbero essere state utilizzate per lo scambio di calore, consentendo a questi pterosauri di assorbire o disperdere calore e regolare la propria temperatura corporea, il che spiegherebbe anche la correlazione tra dimensione della cresta e taglia corporea. Tuttavia, non ci sono prove di vasi sanguigni nella cresta per questo scopo, tuttavia, e le grandi ali membranose piene di vasi sanguigni avrebbero svolto tale compito in modo molto più efficace.[9]
Con queste ipotesi escluse, l'ipotesi meglio supportata per la funzione della cresta sembra essere quella di display sessuale. Ciò è coerente con la variazione dimensionale osservata negli esemplari fossili, dove femmine e giovani hanno creste piccole o addirittura assenti e i maschi hanno creste grandi, elaborate e variabili.[9]
Variazione sessuale
[modifica | modifica wikitesto]
Gli esemplari adulti di Pteranodon possono essere divisi in due classi di dimensioni distinte, piccola e grande, con la classe degli esemplari più grandi, che è circa una volta e mezza più grande della classe più piccola, e la classe degli esemplari più piccoli, che è circa due volte più comune della classe degli esemplari più grandi. Entrambe le classi di dimensioni vivevano fianco a fianco e sebbene inizialmente i ricercatori avevano ipotizzato che si trattassero di specie diverse, Christopher Bennett ha dimostrato che le differenze tra loro sono coerenti con il concetto che rappresentino esemplari femmine e maschi e che le specie di Pteranodon erano sessualmente dimorfiche. I crani della classe di dimensioni maggiori possiedono creste grandi, rivolte verso l'alto e/o all'indietro, mentre le creste della classe di dimensioni più piccole sono piccole e triangolari. Alcuni crani più grandi mostrano anche la prova di una seconda cresta che si estendeva in lungo e in basso, verso la punta del becco, che non si trova in esemplari più piccoli.[9]
Il sesso delle diverse classi di dimensioni era determinato non dai crani, ma dalle ossa pelviche. Contrariamente a quanto ci si potrebbe aspettare, la classe di dimensioni più piccole aveva ossa pelviche sproporzionatamente grandi e larghe. Bennett interpretò ciò come un canale del parto più spazioso, attraverso il quale passavano le uova. Bennett concluse quindi che la classe di piccole dimensioni con creste piccole e triangolari rappresentino esemplari femmine e gli esemplari più grandi con cresta grandi ed elaborate rappresentano esemplari maschi.[9]
Si noti anche che le dimensioni complessive e le dimensioni della cresta corrispondono anche all'età dell'individuo. Sono noti esemplari giovani sia femmine che maschi, e i maschi più giovani presentano spesso piccole creste simili a quelle delle femmine adulte. Pertanto, sembra che le grandi creste si siano sviluppate nei maschi solo quando raggiungevano le loro dimensioni adulte, rendendo difficile stabilire il sesso di esemplari immaturi da resti parziali.[32]
Il fatto che il numero delle femmine superi quello dei maschi, di due a uno, suggerisce che, come negli animali moderni con dimorfismo sessuale correlato alle dimensioni, come i leoni marini e altri pinnipedi, Pteranodon era probabilmente poligamo, con alcuni maschi che competevano per assicurarsi il dominio su un gran numero di femmine. In modo simile ai pinnipedi moderni, i maschi Pteranodon potrebbero aver gareggiato per stabilire un proprio territorio sulle scogliere o sulle spiagge, con i maschi più grandi e con la cresta più grande che occupavano un maggior territorio e avevano più possibilità di accoppiandosi con le femmine. Le creste dei maschio di Pteranodon non sarebbero state usate in combattimento, ma piuttosto come "simboli visivi di rango dominante", con rituali di esibizione che sostituivano la competizione fisica con altri maschi. Se questa ipotesi è corretta, è anche probabile che i maschi di Pteranodon non prendevano parte nell'allevamento dei piccoli; un simile comportamento non si riscontra nei maschi dei moderni animali poligami che generano più prole allo stesso tempo.[9] Tuttavia, i giovani pterosauri sono noti per precoci, ed essere indipendenti e capaci di volare sin dalla schiusa, pertanto non era necessario l'aiuto di nessuno dei genitori.[33]
Paleoecologia
[modifica | modifica wikitesto]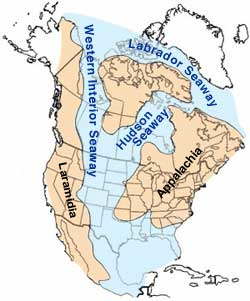
Gli esemplari fossili assegnati a Pteranodon sono stati ritrovati sia nei depositi di Smoky Hill Chalk della Formazione Niobrara, sia nei depositi di Sharon Springs leggermente più recenti della Formazione di Pierre Shale. All'epoca di Pteranodon, questa zona era coperta da un grande mare interno, noto come Mare interno occidentale. Famose per i fossili raccolti dal 1870, queste formazioni si estendono dal sud del Kansas, negli Stati Uniti, fino a Manitoba, in Canada. Tuttavia, gli esemplari di Pteranodon (o eventuali esemplari di altri pterosauri) sono stati ritrovati solo nella metà meridionale della formazione, in Kansas, Wyoming e Dakota del Sud. Nonostante il fatto che numerosi fossili siano stati trovati nelle parti contemporanee della formazione in Canada, nessun esemplare di pterosauro è mai stato trovato lì. Ciò suggerisce che la gamma geografica naturale di Pteranodon copriva solo la parte meridionale della Formazione Niobrara e che il suo habitat non si estendeva più a nord del Dakota del Sud.[8]
Alcuni fossili molto frammentari appartenenti ai pterosauri pteranodonti, e forse allo stesso Pteranodon, sono stati ritrovati anche sulla Costa del Golfo e sulla costa orientale degli Stati Uniti. Ad esempio, alcuni frammenti di ossa della Formazione Mooreville, in Alabama, e la Formazione Merchantville del Delaware potrebbero appartenere a Pteranodon, anche se questi fossili sono troppo frammentari per dare un'identificazione precisa.[8] Anche alcuni resti dal Giappone sono stati provvisoriamente attribuiti a Pteranodon, ma la loro distanza geografica dall'habitat dell'animale rende improbabile questa identificazione.[8]

Pteranodon longiceps condivideva il cielo con lo pterosauro Nyctosaurus, dalla grande cresta. Rispetto a P. longiceps, che era una specie molto comune, Nyctosaurus era più raro, costituendo solo il 3% dei fossili di pterosauro dalla formazione. Anche meno comune era l'uccello dentato, Ichthyornis.[10]
È probabile che, come in altri animali poligami (in cui i maschi competono per il maggior numero di femmine nel proprio harem), Pteranodon viveva principalmente sulle scogliere o isole rocciose al largo, dove poteva nidificare lontano dai predatori terrestri e nutrirsi lontano dalla costa; la maggior parte dei fossili di Pteranodon si trovano in luoghi che all'epoca erano a centinaia di chilometri dalla costa.[9]
Sotto la superficie, il mare era popolato principalmente da invertebrati come ammoniti e calamari. I vertebrati marini erano estremamente comuni ed, oltre ai pesci, comprendevano tartarughe marine, come Toxochelys, il plesiosauro Styxosaurus e l'uccello marino incapace di volare Parahesperornis. I mosasauri erano i rettili marini più comuni, con generi tra cui Clidastes e Tylosaurus.[1] Alcuni fossili eccezionali ci dimostrano che almeno alcuni di questi rettili marini si sono nutriti di Pteranodon. Barnum Brown, nel 1904, riportò lo scoperta del contenuto dello stomaco di un plesiosauro che conteneva ossa di "pterosauro", molto probabilmente uno Pteranodon.[36]
All'interno dei gessi della Formazione Niobrara sono stati ritrovati anche i resti di dinosauri terrestri, suggerendo che gli animali morti sulla riva venivano portati via dal mare o trascinati dai fiumi, dove le loro carcasse venivano consumate da organismi marini (come nel caso di un esemplare di adrosauro che mostra segni di morsi di uno squalo).[37]
Periodo di vita ed evoluzione
[modifica | modifica wikitesto]In termini generali, lo Pteranodon sarebbe vissuto nella fine degli anni del Coniaciano ai primi anni delle fasi del Cretaceo. Però, gli anni precisi in cui è vissuto non sono ancora stati del tutto confermati; nel 2003, Kenneth Carpenter ha esaminato la distribuzione e la datazione dei fossili, dimostrando che lo Pteranodon esisteva 88-85 milioni di anni fa, mentre Pteranodon Longiceps esisteva tra i 86 e 84,5 milioni di anni fa. Invece nel 2010, la formazione Pierre Shale in Kansas, Wyoming e Dakota del Sud, ha ipotizzato che il noto rettile preistorico sia risalente tra 81,5 e 80,5 milioni di anni fa.
Nei primi anni del 1990, è stato notato che il principale Polimorfismo dello Pteranodon si è separato in poco tempo. Ciò ha suggerito, insieme alla loro somiglianza globale, che probabilmente rappresentano all'interno un unico ceppo evolutivo della durata di circa 4 milioni di anni. In altre parole, una sola specie di Pteranodon sarebbe stata presente in un dato momento, e lo Pteranodon sternbergi con ogni probabilità è stato l'antenato diretto dello Pteranodon longiceps.
Nella cultura di massa
[modifica | modifica wikitesto]Lo Pteraodon è per tutti l'immagine del rettile volante per eccellenza, diventando quindi per tutti il più famoso membro dei rettili volanti e l’animale preistorico volante più famoso, alcune volte viene tuttavia erroneamente chiamato Pterodattilo per via della somiglianza tra i due nomi, le apparizioni dello Pteranodon nei media cinematografici e nei libri sono innumerevoli, tra cui:
- Uno Pteranodon viene assalito e ucciso in volo da un Tyrannosaurus nel film Il mondo perduto.
- Nel film King Kong uno Pteranodon rapisce Ann Darrow e viene affrontato e ucciso da Kong. Lo pteranodonte compare con ruoli simili anche in Un milione di anni fa dove si scontra con un Rhamphorhynchus gigantesco, e La vendetta di Gwangi.
- Nei film animati della saga Alla ricerca della Valle Incantata uno Pteranodon di nome Petrie è uno dei personaggi principali.
- Nella serie animata Il treno dei dinosauri i protagonisti sono pteranodonti.
- Nel film Rodan, il mostro alato fa il suo debutto Rodan, uno Pteranodon modificato dalle radiazioni, che in seguito diventerà uno dei più famosi mostri del cinema giapponese.
- Uno Pteranodonte appare nel film Adèle e l'enigma del faraone del 2010 diretto da Luc Besson e viene cavalcato dalla protagonista Adèle Blanc-Sec.
- Lo Pteranodonte appare anche nella celebre saga cinematografica Jurassic Park nella sequenza finale di Il mondo perduto - Jurassic Park (1997), in Jurassic Park III (2001) con delle dimensioni assai superiori, simili quelle del Quetzalcoatlus per impressionare il pubblico, in Jurassic World (2015), Jurassic World - Il regno distrutto (2018) e Jurassic World - Il dominio (2022), compare anche nelle serie animate Jurassic World - Nuove avventure, Jurassic World - Teoria del caos e nel prodotti LEGO non canonici, come LEGO Jurassic World: L'evasione di Indominus Rex, LEGO Jurassic World: La mostra segreta e LEGO Jurassic World - La leggenda di Isla Nublar. Si può sbloccare in Jurassic World Evolution grazie al DLC basato sulla trilogia di Jurassic Park con il design del terzo film, e in Jurassic World Evolution 2.
- Il Pokémon Aerodactyl è basato sul modello di questo animale.
- King, personaggio del manga e anime One Piece, ha la capacità di trasformarsi in uno Pteranodon grazie ai poteri del suo frutto del diavolo.
- Degli pteranodonti appaiono in Fantasia (1940).
- Appare come mostro nelle serie animate The Scooby-Doo Show e Be Cool, Scooby-Doo! e nel film Scooby-Doo 2 - Mostri scatenati.
- Alcune ricostruzioni di pteranodonte sono presenti in diversi parchi tematici tra cui: il Parco della Preistoria di Rivolta d'Adda (CR), il Parco dei Dinosauri di Castellana Grotte (BA), il Parco Natura Viva di Bussolengo (VR) e il parco "Il Mondo della Preistoria" di Simbario (VV)
- Nella sigla della serie televisiva Teen Titans, il personaggio di Beast Boy si trasforma in uno Pteranodon, questa sua trasformazione avviene anche molto frequente all’interno della serie.
Note
[modifica | modifica wikitesto]- ^ a b c d Bennett, S.C. (2000). "Inferring stratigraphic position of fossil vertebrates from the Niobrara Chalk of western Kansas." Current Research in Earth Sciences: Kansas Geological Survey Bulletin, 244(Part 1): 26 pp.
- ^ a b c d e S.C. Bennett, The Pterosaurs of the Niobrara Chalk, in The Earth Scientist, vol. 11, n. 1, 1994, pp. 22-25.
- ^ a b M.P. Witton e M.B. Habib, On the Size and Flight Diversity of Giant Pterosaurs, the Use of Birds as Pterosaur Analogues and Comments on Pterosaur Flightlessness, in PLoS ONE, vol. 5, n. 11, 2010, p. e13982, Bibcode:2010PLoSO...513982W, DOI:10.1371/journal.pone.0013982, PMC 2981443, PMID 21085624.
- ^ a b Bennett, S. C. (1987). "New evidence on the tail of the pterosaur Pteranodon (Archosauria: Pterosauria)." Pp. 18–23 in Currie, P. J. e E. H. Koster (eds.), Fourth Symposium on Mesozoic Terrestrial Ecosystems, Short Papers. Occasional Papers of the Tyrrell Museum of Paleontology, #3.
- ^ a b c d Kellner, A.W.A., Comments on the Pteranodontidae (Pterosauria, Pterodactyloidea) with the description of two new species (PDF), in Anais da Academia Brasileira de Ciências, vol. 82, n. 4, 2010, pp. 1063-1084, DOI:10.1590/S0001-37652010000400025.
- ^ Martin-Silverstone, E., Glasier, J., Acorn, J., Mohr, S., and Currie, P. 2017. Reassesment of Dawndraco kanzai Kellner, 2010 and reassignment of the type specimen to Pteranodon sternbergi Harksen, 1966. Vertebrate Anatomy Morphology Palaeontology, 3:47-59. DOI: 10.18435/B5059J
- ^ Piero e Alberto Angela, Rettili Volanti, in "Il pianeta dei dinosauri", Arnoldo Mondadori Editore, 1993, p. 111, ISBN 88-04-41235-6.
- ^ a b c d e f g h i j k l m n o p q r s Bennett, S.C., Taxonomy and systematics of the Late Cretaceous pterosaur Pteranodon (Pterosauria, Pterodactyloida), in Occasional Papers of the Natural History Museum, University of Kansas, vol. 169, 1994, pp. 1-70.
- ^ a b c d e f g h i j k l Bennett, S.C., Sexual dimorphism of Pteranodon and other pterosaurs, with comments on cranial crests, in Journal of Vertebrate Paleontology, vol. 12, n. 4, 1992, pp. 422-434, DOI:10.1080/02724634.1992.10011472.
- ^ a b K Carpenter, Vertebrate Biostratigraphy of the Smoky Hill Chalk (Niobrara Formation) and the Sharon Springs Member (Pierre Shale), in High-Resolution Approaches in Stratigraphic Paleontology, vol. 21, 2003, pp. 421-437, DOI:10.1007/978-1-4020-9053-0.
- ^ Zimmerman, H., Preiss, B., and Sovak, J. (2001). Beyond the Dinosaurs!: sky dragons, sea monsters, mega-mammals, and other prehistoric beasts, Simon and Schuster. ISBN 0-689-84113-2.
- ^ B. Andres e T. S. Myers, Lone Star Pterosaurs, in Earth and Environmental Science Transactions of the Royal Society of Edinburgh, vol. 103, 2013, p. 1, DOI:10.1017/S1755691013000303.
- ^ B. Andres, J. Clark e X. Xu, The Earliest Pterodactyloid and the Origin of the Group, in Current Biology, vol. 24, 2014, pp. 1011-6, DOI:10.1016/j.cub.2014.03.030, PMID 24768054.
- ^ a b Peter Wellnhofer, The Illustrated Encyclopedia of Pterosaurs, New York, Barnes and Noble Books, 1996 [1991], p. 139, ISBN 0-7607-0154-7.
- ^ Cope, E.D. (1875). "The Vertebrata of the Cretaceous formations of the West." Report, U. S. Geological Survey of the Territories (Hayden), 2: 302 pp., 57 pls.
- ^ Marsh, O.C., Notice of a new sub-order of Pterosauria, in American Journal of Science, Series 3, vol. 11, n. 65, 1876a, pp. 507-509, DOI:10.2475/ajs.s3-11.66.507.
- ^ Marsh, O.C., Principal characters of American pterodactyls, in American Journal of Science, Series 3, vol. 12, n. 72, 1876b, pp. 479-480, DOI:10.2475/ajs.s3-12.72.479.
- ^ A.O. Averianov, Ornithostoma sedgwicki – valid taxon of azhdarchoid pterosaurs, in Proceedings of the Zoological Institute RAS, vol. 316, n. 1, 2012, pp. 40-49.
- ^ Allt om Vetenskap, 10-2012, p.72
- ^ Padian, K. (1983), "A functional analysis of flying and walking in pterosaurs"., in Paleobiology., 9 (3), 218–239.
- ^ a b Witton, M.P.; Habib, M.B. (2010). "On the Size and Flight Diversity of Giant Pterosaurs, the Use of Birds as Pterosaur Analogues and Comments on Pterosaur Flightlessness", su journals.plos.org.
- ^ Unwin, David M., The Pterosaurs: From Deep Time, New York: Pi Press, 2006, pp. 210-222, ISBN 978-0-13-146308-0.
- ^ a b CLAW MORPHOLOGY OF PTERANODON AND POSSIBLE AQUATIC LOCOMOTION, su gsa.confex.com. URL consultato il 15 aprile 2019 (archiviato dall'url originale il 7 gennaio 2011).
- ^ a b Bennett, S.C. (1994), "The Pterosaurs of the Niobrara Chalk", in The Earth Scientist., vol. 11, (1): 22–25.
- ^ a b Bennett, S.C. (1994), "The Pterosaurs of the Niobrara Chalk", in The Earth Scientist., vol. 11, (1): 22-25.
- ^ Bennett, S.C. (1994), "The Pterosaurs of the Niobrara Chalk", in The Earth Scientist, vol. 11, (1): 22-25.
- ^ J. L. Tomkins, N. R. Lebas, M. P. Witton, D. M. Martill e S. Humphries, Positive Allometry and the Prehistory of Sexual Selection (PDF), in The American Naturalist, vol. 176, n. 2, 2010, pp. 141-148, DOI:10.1086/653001, PMID 20565262. URL consultato il 2 settembre 2019 (archiviato dall'url originale il 9 agosto 2017).
- ^ a b Eaton, G.F. (1910). "Osteology of Pteranodon." Memoirs of the Connecticut Academy of Arts and Sciences, 2:1–38, pls. i–xxxi.
- ^ von Kripp, D. (1943). "Ein Lebensbild von Pteranodon ingens auf flugtechnischer Grundlage." Nova Acta Leopoldina, N.F., 12(83): 16–32 [in German].
- ^ Stein, R.S., Dynamic analysis of Pteranodon ingens: a reptilian adaptation to flight, in Journal of Paleontology, vol. 49, 1975, pp. 534-548.
- ^ Bramwell, C.D. e Whitfield, G.R. (1974). "Biomechanics of Pteranodon." Philosophical Transactions Royal Society B, 267.
- ^ Bennett, S.C., The osteology and functional morphology of the Late Cretaceous pterosaur Pteranodon. General description of osteology, in Palaeontographica, Abteilung A, vol. 260, 2001, pp. 1-112.
- ^ Xiaolin Wang, Alexander W.A. Kellner, Shunxing Jiang, Qiang Wang, Yingxia Ma, Yahefujiang Paidoula, Xin Cheng, Taissa Rodrigues, Xi Meng, Jialiang Zhang, Ning Li e Zhonghe Zhou, Sexually dimorphic tridimensionally preserved pterosaurs and their eggs from China, in Current Biology, vol. 24, n. 12, 2014, pp. 1323-1330, DOI:10.1016/j.cub.2014.04.054, PMID 24909325.
- ^ vertebre cervicali di un pteradonte con un dente di squalo conficcato in una di esse
- ^ Mark P. Witton's Blog: No, Cretaceous sharks did not leap from water to eat flying pterosaurs
- ^ Brown, B., Stomach stones and the food of plesiosaurs, in Science, vol. 20, n. 501, 1904, pp. 184-185, Bibcode:1904Sci....20..184B, DOI:10.1126/science.20.501.184, PMID 17737868.
- ^ M.J. Everhart e K. Ewell, [27:sdhcvf2.0.co;2 Shark-bitten dinosaur (Hadrosauridae) vertebrae from the Niobrara Chalk (Upper Coniacian) of western Kansas], in Transactions of the Kansas Academy of Science, vol. 109, 1–2, 2006, pp. 27-35, DOI:10.1660/0022-8443(2006)109[27:sdhcvf]2.0.co;2.
Altri progetti
[modifica | modifica wikitesto]Wikimedia Commons contiene immagini o altri file su Pteranodon
Wikispecies contiene informazioni su Pteranodon
Collegamenti esterni
[modifica | modifica wikitesto]- (EN) Pteranodon, su Enciclopedia Britannica, Encyclopædia Britannica, Inc.
- (EN) Pteranodon / Pteranodon (altra versione) / Pteranodon (altra versione) / Pteranodon (altra versione), su Fossilworks.org.
| Controllo di autorità | Thesaurus BNCF 59790 · LCCN (EN) sh85108544 · BNF (FR) cb125455988 (data) · J9U (EN, HE) 987007546282105171 |
|---|