Caenorhabditis elegans
| Caenorhabditis elegans | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
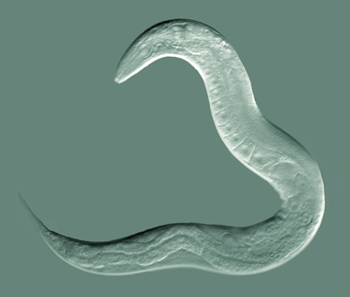
 Verme adulto hermafrodita de C. elegans | |||||||||||||||
| Clasificación científica | |||||||||||||||
| |||||||||||||||
| Nome binomial | |||||||||||||||
| Caenorhabditis elegans (Maupas, 1900)[1] | |||||||||||||||
Caenorhabditis elegans é unha especie de verme nematodo de vida libre, transparente, de 1 mm de lonxitude,[2] que vive en ambientes de solos temperados. O seu nome procede do grego caeno-, recente, rhabditis, con forma de bastón,[3] e do latín elegans, elegante. En 1900, Maupas chamouno inicialmente Rhabditides elegans, despois, en 1952, Osche situouno no subxénero Caenorhabditis, e, finalmente, Dougherty elevouno á categoría de xénero en 1955.[4] En 1974, Sydney Brenner empezou a investigar en bioloxía molecular e do desenvolvemento de C. elegans, o cal desde entón foi moi utilizado como organismo modelo.[5]
C. elegans foi o primeiro organismo multicelular do que se completou a secuenciación do xenoma. E en 2012, este verme era o único organismo do que se completou o conectoma (diagrama do "cableado" neuronal).[6]
Anatomía
[editar | editar a fonte]C. elegans é un verme non segmentado de simetría bilateral. Ten unha cutícula, catro cordóns musculares epidérmicos principais, e un pseudoceloma (cavidade corporal) cheo de líquido. Case todos os individuos de C. elegans son hermafroditas con aspecto de femia; e hai unha minoría (1 de cada mil) de machos verdadeiros.[7] A anatomía básica de C. elegans inclúe unha boca, farinxe, intestino, gónadas, e cutícula de coláxeno. Os machos teñen unha gónada dun só lobo, un conduto deferente, e unha cola especializada para o apareamento, a cal ten espículas. Os hermafroditas teñen dous ovarios, ovidutos, espermateca, e un só útero. As catro faixas musculares que discorren por toda a lonxitude do seu corpo están conectadas por un sistema neural que permite que os músculos movan o corpo do animal só na dirección dorsal/ventral; por tanto, calquera individuo vivo móbil está sempre sobre o seu lado esquerdo ou dereito cando se observa cruzando unha superficie horizontal.
Gránulos intestinais e fluorescencia
[editar | editar a fonte]Nas células do intestino ten numerosos gránulos intestinais, cuxa función non se coñece ben, o mesmo que ocorre con outros aspectos deste nematodo, malia os moitos anos que leva sendo estudado. Estes gránulos do tubo dixestivo encóntranse en todos os membros da orde Rhabdita. Son moi similares a lisosomas no sentido de que teñen un interior ácido e capacidade de endocitose, pero son considerablemente máis longos, o que reforza a idea de que son orgánulos de almacenamento. Unha característica salientable dos gránulos é que vistos baixo luz ultravioleta, reaccionan emitindo unha intensa fluorescencia azul. Outro fenómeno observado é a denominada fluorescencia da morte, que consiste en que cando morre o verme ten lugar unha forte explosión de fluorescencia azul. Esta fluorescencia da morte ten lugar normalmente nunha onda que vai desde a parte anterior á posterior do intestino, e obsérvase en vermes novos e vellos, sometidos a unha ferida mortal, ou en vermes vellos que morren placidamente. Propuxéronse moitas teorías sobre as funcións dos gránulos intestinais e moitas das iniciais foron desbotadas polos experimentos que se fixeron posteriormente. Crese que almacenan cinc como unha das súas funcións. Recentes análises químicas identificaron o material fluorescente azul, o cal contiña unha forma glicosilada de ácido antranílico (AA). Pero cuestiónase a necesidade das grandes cantidades de ácido antranílico que conteñen os gránulos. Unha posibilidade é que o ácido antranílico sexa un produto antibacteriano usado para defenderse dos patóxenos invasores. Outra posibilidade é que os gránulos proporcionen unha fotoprotección: a explosión de fluorescencia de ácido antranílico supón a conversión da luz ultravioleta daniña e luz visible relativamente inofensiva. Podería haber unha posible semellanza cos melanosomas que conteñen melanina doutros animais.[8]
Reprodución
[editar | editar a fonte]
O hermafrodita considérase que é unha forma especializada de femia autofecundable porque ten un soma de femia pero a súa liña xerminal produce primeiro gametos masculinos, despois produce ovocitos, e finalmente pon ovos por medio do seu útero despois de que se produciu unha fertilización interna. En condicións ambientais favorables para a reprodución, as larvas que saen dos ovos desenvólvense pasando por catro etapas ou mudas, denominadas de L1 a L4. Cando as condicións son estresantes como pode ser a insuficiencia de alimentos, C. elegans pode entrar nunha terceira fase larval alternativa chamada estado dauer (dauer é unha palabra alemá que significa permanente). As larvas dauer son resistentes ao estrés e non envellecen. Os individuos hermafroditas producen todos o seu propio esperma na etapa L4 (150 espermatozoides por brazo gonadal) e despois deixan de producir esperma e producen só ovocitos. Os espermatozoides almacénanse na mesma área da gónada que os ovocitos ata que o primeiro ovocito empuxa o espermatozoide cara á espermateca (unha cámara onde se produce a fertilización dos ovocitos polos espermatozoides).[9] Non sempre hai autofecundación, xa que os machos poden tamén inseminar aos hermafroditas, os cales prefiren usar o esperma de machos antes que o propio (ambos os tipos de esperma almacénense na espermateca). Cando se autofecunda un destes vermes de tipo salvaxe pon aproximadamente 300 ovos. Cando é inseminado por un macho, o número de ovos pode ser de máis de mil. A 20 °C, a cepa de laboratorio de C. elegans ten unha vida media de aproximadamente dúas ou tres semanas e un tempo de xeración de aproximadamente catro días.

Ecoloxía
[editar | editar a fonte]As especies do xénero Caenorhabditis ocupan varios ambientes ricos en nutrientes e bacterias. Aliméntanse das bacterias que viven na materia orgánica en descomposición. Xeralmente, o solo non ten materia orgánica dabondo para manter poboacións autosustentables. C. elegans pode sobrevivir cunha dieta de diversas especies de bacterias, pero a súa ecoloxía no ambiente salvaxe é bastante descoñecida. A maioría das cepas de laboratorio recolléronse de ambientes artificiais como xardíns e compost. Máis recentemente, viuse que C. elegans prospera noutros lugares con materia orgánica, como a froita podre.[10] Algúns invertebrados como milpés, insectos, isópodos, e gasterópodos poden transportar larvas dauer. Tense observado que estas larvas se alimentan do seu propio hóspede cando este morre.[11]
Os nematodos poden sobrevivir ao desecamento, e no caso de C. elegans o mecanismo para esta capacidade demostrouse que se debe ás proteínas abundantes na embrioxénese tardía (LEA; late embryogenesis abundant proteins).[12]
Uso en investigación
[editar | editar a fonte]En 1963, Sydney Brenner propuxo a utilización de C. elegans como organismo modelo para a investigación primaria do desenvolvemento neural nos animais, xa que era un dos organismos máis simples con sistema nervioso. No individuo hermafrodita, este sistema comprende 302 neuronas[13] cuxo patrón foi mapado e coñécese como conectoma, e mostrou ser un gráfico matemático de tipo rede de mundo pequeno.[14] Os investigadores exploraron os mecanismos neurais que controlan varios dos comportamentos máis interesantes de C. elegans; por exemplo, a quimiotaxe, termotaxe, mecanotransdución, e o comportamento de apareamento do macho.[15] Brenner tamén escolleu a esta especie como útil para facela crecer en poboacións masivas e conveniente para análises xenéticas.[16] É un organismo multicelular eucariótico que é o suficientemente simple como para ser estudado en grande detalle. As cepas reprodúcense doadamente e poden ser conxeladas. Cando posteriormente se desconxelan, permanecen viables e iso permite que sexan almacenadas durante longo tempo.[17]
A transparencia de C. elegans facilita o estudo da diferenciación celular e outros procesos do desenvolvemento no organismo intacto. A morfoloxía da rexión da cola distingue claramente os machos dos hermafroditas.
Mapouse tamén o destino en que acaba cada célula somática durante o desenvolvemento (959 células no hermafrodita adulto, e 1031 no macho adulto).[18][19] Estes patróns de liñaxes celulares son en grande medida invariables entre individuos, a diferenza do que ocorre nos mamíferos, nos que o desenvolvemento celular no embrión depende máis de sinais celulares.
As primeiras divisións celulares na embrioxénese temperá están entre os exemplos mellor coñecidos de división celular asimétrica.[20]
A morte celular programada (apoptose) elimina moitas células adicionais (131 no hermafrodita, a maioría das cales teríanse convertido doutro modo en neuronas); esta "preditibilidade apoptótica" contribuíu á detectar algúns xenes apoptóticos, principalmente por medio da observación de nematodos anormais que sobrevivían á apoptose.

A interferencia de ARN (RNAi), é un método relativamente directo de interromper a función de xenes específicos. Silenciar a función dun xene pode ás veces permitir aos investigadores inferir a súa posible función. O nematodo pode ser mergullado ou inxectado cunha solución de ARN de dobre cadea, cuxa secuencia é complementaria da secuencia do xene que os investigadores desexan inutilizar; os vermes poden alternativamente ser alimentados con bacterias transformadas xeneticamente que expresan o ARN de dobre cadea que interesa. Os experimentos de perda de función dos xenes en C. elegans son os máis fáciles de facer de entre todos os modelos aimais, o que permitiu aos científicos establecer que só o 9% dos 20.000 xenes do xenoma ten un papel funcional.[21]
A captación de ARNi ambiental é moito peor noutras especies de vermes do xénero Caenorhabditis. Aínda que inxectar o ARN na cavidade corporal do animal induce o silenciamento de xenes na maioría das especies, só en C. elegans e nuns poucos nematodos máis distantemente emparentados poden captarse ARN das bacterias das que se alimentaron para a interferencia de ARN.[22] Esta capacidade foi mapada nun xene chamado sid-2, o cal, cando se insire como un transxene noutras especies, permítelles captar ARN para o ARNi como fai C. elegans.[23]
O estudo da meiose é considerablemente simplificado neste verme. A medida que os núcleos dos ovocitos e espermatozoides se moven ao longo da gónada, progresan temporalmente polas fases meióticas. As dificultades que presenta o estudo de poboacións celulares heteroxéneas quedan eliminadas porque cada núcleo situado nunha posición dada na gónada está aproximadamente na mesma fase da meiose.
Pode tamén utilizarse para estudar a dependencia á nicotina porque o verme mostra respostas comportamentais á nicotina que son paralelas ás dos mamíferos; por exemplo, respostas agudas, tolerancia, síndrome de abstinencia, e sensibilización.[24]
Como ocorre coa maioría dos organismos modelo, os científicos que traballan neste campo manteñen unha coidada base de datos en liña para C. elegans chamada WormBase. A WormBase trata de inclír todas as informacións publicadas sobre C. elegans e outros nematodos relacionados.[25]
C. elegans utilizouse como organismo modelo para a investigación do envellecemento; por exemplo, a inhibición dunha vía de sinalización do factor de crecemento similar á insulina (IGF) incrementaba tres veces a duración da vida do adulto.[26] Ademais, unha extensa investigación en C. elegans identificou que as proteínas de unión ao ARN son factores esenciais durante o desenvolvemento do liña xerminal e o embrión temperán.
C. elegans ten cinco pares de autosomas e un par de cromosomas sexuais. O sexo en C. elegans está baseado no sistema de determinación do sexo X0. Os C. elegans hermafroditas teñen un par de cromosomas sexuais XX; os escasos machos teñen só un cromosoma sexual, polo que son X0. O espermatozoide de C. elegans é ameboide, sen flaxelo nin acrosoma.
C. elegans é notable tamén nos estudos do sono animal, xa que é o organismo máis primitivo que mostra estados similares ao sono. En C. elegans, hai unha fase "letárxica" pouco despois de cada muda.
Investigacións no espazo
[editar | editar a fonte]C. elegans foi noticia cando se descubriu que algúns espécimes sobreviviran ao desastre da lanzadeira espacial Columbia en febreiro de 2003.[27] Despois, en 2009, enviáronse mostas de espécimes vivos de C. elegans da Universidade de Nottingham á Estación Espacial Internacional, onde pasaron dúas semanas, nun proxecto para explorar os efectos da gravidade cero no desenvolvemento muscular e a fisioloxía. A investigación trataba principalmente sobre a base xenética da atrofia muscular, relacionada con viaxes espaciais, prostración prolongada en cama, casos xeriátricos, ou diabéticos.[28] Descendentes dos vermes que ían abordo da Columbia en 2003 foron enviados ao espazo na lanzadeira espacial Endeavour na súa misión STS-134.[29]
Xenoma
[editar | editar a fonte]
C. elegans foi o primeiro organismo multicelular do que se completou a secuenciación do xenoma, que foi publicada en 1998,[30] aínda que quedaban algúns ocos por secuenciar, os cales non foron resoltos ata 2002. O xenoma de C. elegans é de aproximadamente 100 millóns de pares de bases e consta de seis cromosomas denominados I, II, III, IV, V e X, e un xenoma mitocondrial. A súa densidade de xenes é de aproximadamente 1 xene/5kb. Os intróns, ou secuencias non expresadas, son o 26% do xenoma. Algunhas rexións interxénicas grandes conteñen secuencias de ADN repetitivo. Moitos xenes están dispostos formando operóns, que son series policistrónicas que se transcriben xuntas. C. elegans e outros nematodos son uns dos poucos nematodos que actualmente se sabe que teñen operóns (que son típicos das bacterias); entre estes están os tripanosomas, vermes planos como o trematodo Schistosoma mansoni, e o primitivo cordado tunicado Oikopleura dioica. Crese que moitos máis organismos poderían ter estes operóns.[31]
O xenoma contén aproximadamente 20.470 xenes que codifican proteínas.[32] O número de xenes de ARN coñecidos no xenoma aumentou moito debido ao descubrimento en 2006 da nova clase de xenes 21U-RNA,[33] e agora pénsase que o xenoma contén máis de 16.000 xenes de ARN, cando en 2005 pensábase que eran 1.300.[34] Os científicos continúan a avaliar o conxunto de novos xenes: engádense decote novas predicións de xenes e modifícanse ou elimínanse os que se consideran incorrectos.
En 2003, determinouse tamén a secuencia do xenoma do nematodo relacionado C. briggsae, o que permitiu aos investigadores estudar a xenómica comparada deses dous organismos.[35] As secuencias xenómicas doutros nematodos do mesmo xénero por exemplo, C. remanei,[36] C. japonica[37] e C. brenneri están a estudarse por medio da técnica da escopeta de xenoma completo, que é menos completa e precisa que a aproximación "xerárquica" ou clon por clon que foi a que se utilizou con C. elegans.[38]
A versión oficial da secuencia xenómica de C. elegans segue cambiando conforme aparecen novas evidencias que mostran os erros da secuenciación orixinal. A maioría dos cambios son menores, engadindo ou eliminando uns poucos pares de bases do ADN. Por exemplo, a actualización WS202 da WormBase de abril de 2009 engadiu dous pares de bases á secuencia xenómica.[39] Ás veces hai que facer cambios máis amplos; por exemplo, a actualización WS197 de decembro de 2008 engadiu unha rexión duns 4.600 pares de bases á secuencia.[40][41]
Evolución
[editar | editar a fonte]Unhas poucas secuencias de proteínas conservadas estudadas en esponxas mariñas son máis parecidas ás humanas que ás de C. elegans.[42] Isto indica que debeu de haber unha acelerada velocidade de evolución na liñaxe de C. elegans. O mesmo estudo encontrou que varios xenes filoxeneticamente antigos están ausentes en C. elegans.
Comunidade científica
[editar | editar a fonte]En 2002, outorgouse o Premio Nobel de Medicina e Fisioloxía a Sydney Brenner, H. Robert Horvitz e John Sulston polos seus traballos sobre a xenética do desenvolvemento de órganos e morte celular programada en C. elegans. O Premio Nobel de Medicina de 2006 concedéuselle a Andrew Fire e Craig C. Mello polo seu descubrimento da interferencia de ARN en C. elegans.[43] En 2008, Martin Chalfie compartiu o Premio Nobel de Química polo seu traballo sobre a proteína fluorescente verde; algúns destes traballos realizáronse usando C. elegans.
Moitos científicos que investigan en C. elegans están estreitamente conectados con Sydney Brenner, co cal empezaron case todas as investigacións neste eido na década de 1970; traballaron como investigadores posdoutorais ou posgraduados no laboratorio de Brenner ou no laboratorio de alguén que previamente traballara con Brenner. A maioría dos que traballaron neste laboratorio posteriormente estableceron os seus propios laboratorios de investigacións de vermes. A base de datos WormBase contén datos da bioloxía e xenoma de C. elegans.
Notas
[editar | editar a fonte]- ↑ Maupas, É (1900). "Modes et formes de reproduction des nématodes". Archives de Zoologie Expérimentale et Générale 8: 463–624.
- ↑ Wood, WB (1988). The Nematode Caenorhabditis elegans. Cold Spring Harbor Laboratory Press. p. 1. ISBN 0-87969-433-5.
- ↑ καινός (caenos) = novo, recente; ῥάβδος (rhabdos) = bastón, variña.
- ↑ Ferris, H (30 November 2013). "Caenorhabditis elegans". University of California, Davis. Arquivado dende o orixinal o 09 de decembro de 2013. Consultado o 2013-11-19.
- ↑ Brenner, S (1974). "The Genetics of Caenorhabditis elegans". Genetics 77 (1): 71–94. PMC 1213120. PMID 4366476.
- ↑ Jabr, Ferris (2012-10-2). "The Connectome Debate: Is Mapping the Mind of a Worm Worth It?". Scientific American. Consultado o 2014-1-18.
- ↑ Alberts, B; Johnson, A; Lewis, J; Raff, M; Roberts, K; Walter, P (2007). Molecular Biology of the Cell (5th ed.). Garland Science. p. 1321. ISBN 978-0-8153-4105-5.
- ↑ Coburn, C; Gems, D (2013). "The mysterious case of the C. Elegans gut granule: Death fluorescence, anthranilic acid and the kynurenine pathway". Frontiers in Genetics 4: 151. PMC 3735983. PMID 23967012. doi:10.3389/fgene.2013.00151.
- ↑ Nayak, S; Goree, J; Schedl, T (2004). "fog-2 and the Evolution of Self-Fertile Hermaphroditism in Caenorhabditis". PLoS Biology 3 (1): e6. PMC 539060. PMID 15630478. doi:10.1371/journal.pbio.0030006.
- ↑ Félix, MA; Braendle, C (2010). "The natural history of Caenorhabditis elegans". Current Biology 20 (22): R965–R969. PMID 21093785. doi:10.1016/j.cub.2010.09.050.
- ↑ Kiontke, K; Sudhaus, W (2006). "Ecology of Caenorhabditis species". WormBook: 1–14. PMID 18050464. doi:10.1895/wormbook.1.37.1.
- ↑ Gal, TZ; Glazer, I; Koltai, H (2004). "An LEA group 3 family member is involved in survival of C. elegans during exposure to stress". FEBS Letters 577 (1–2): 21–26. PMID 15527756. doi:10.1016/j.febslet.2004.09.049.
- ↑ Kosinski, RA; Zaremba, M (2007). "Dynamics of the Model of the Caenorhabditis elegans Neural Network". Acta Physica Polonica B 38 (6): 2201. Bibcode:2007AcPPB..38.2201K.
- ↑ Watts, DJ; Strogatz, SH (1998). "Collective dynamics of 'small-world' networks". Nature 393 (6684): 440–442. Bibcode:1998Natur.393..440W. PMID 9623998. doi:10.1038/30918.
- ↑ Schafer, W.R. Deciphering the neural and molecular mechanisms of C. Elegans behaviour. Curr.Biol.2005.Sep 6.PMID 16139205
- ↑ "Genetic code, C. elegans, and recombinant DNA". Albert and Mary Lasker Foundation. Consultado o 21/3/2016.
- ↑ Brenner, S (1974). "The Genetics of Caenorhabditis elegans". Genetics 77 (1): 71–94. PMC 1213120. PMID 4366476.
- ↑ Sulston, JE; Horvitz, HR (1977). "Post-embryonic cell lineages of the nematode, Caenorhabditis elegans". Developmental Biology 56 (1): 110–56. PMID 838129. doi:10.1016/0012-1606(77)90158-0.
- ↑ Kimble, J; Hirsh, D (1979). "The postembryonic cell lineages of the hermaphrodite and male gonads in Caenorhabditis elegans". Developmental Biology 70 (2): 396–417. PMID 478167. doi:10.1016/0012-1606(79)90035-6.
- ↑ Gönczy, P (2005). "Asymmetric cell division and axis formation in the embryo". WormBook: 1. doi:10.1895/wormbook.1.30.1.
- ↑ Kamath, RS; et al. (2003). "Systematic functional analysis of the Caenorhabditis elegans genome using RNAi". Nature 421 (6920): 231–237. Bibcode:2003Natur.421..231K. PMID 12529635. doi:10.1038/nature01278.
- ↑ Félix, M-A (2008). "RNA interference in nematodes and the chance that favored Sydney Brenner". Journal of Biology 7 (9): 34–56. PMC 2776389. PMID 19014674. doi:10.1186/jbiol97.
- ↑ Winston, WM; Sutherlin, M; Wright, AJ; Feinberg, EH; Hunter, CP (2007). "Caenorhabditis elegans SID-2 is required for environmental RNA interference". Proceedings of the National Academy of Sciences 104 (25): 10565. Bibcode:2007PNAS..10410565W. doi:10.1073/pnas.0611282104.
- ↑ Feng, Z; Li, W; Ward, A; Piggott, BJ; Larkspur, ER; Sternberg, PW; Xu, XZ (2006). "A C. elegans model of nicotine-dependent behavior: regulation by TRP family channels". Cell 127 (3): 621–633. PMC 2859215. PMID 17081982. doi:10.1016/j.cell.2006.09.035.
- ↑ Dolgin, E (2007). "Slime for a dime". Science 317 (5842): 1157. doi:10.1126/science.317.5842.1157b.
- ↑ Wolkow, CA; Kimura, KD; Lee, M-S; Ruvkun, G (2000). "Regulation of C. elegans Life-Span by Insulin-like Signaling in the Nervous System". Science 290 (5489): 147–150. Bibcode:2000Sci...290..147W. PMID 11021802. doi:10.1126/science.290.5489.147.
- ↑ "Worms survived Columbia disaster". BBC News. 1 May 2003. Consultado o 2008-07-11.
- ↑ "University sends worms into space". BBC News. 17 January 2009. Consultado o 2009-07-09.
- ↑ Klotz, I (16 May 2011). "Legacy Space Worms Flying on Shuttle". Discovery News. Arquivado dende o orixinal o 16 de xuño de 2012. Consultado o 2011-05-17.
- ↑ The C. elegans Sequencing Consortium (1998). "Genome sequence of the nematode C. elegans: A platform for investigating biology". Science 282 (5396): 2012–2018. PMID 9851916. doi:10.1126/science.282.5396.2012.
- ↑ Blumenthal, T (2004). "Operons in eukaryotes". Briefings in Functional Genomics and Proteomics 3 (3): 199–211. PMID 15642184. doi:10.1093/bfgp/3.3.199.
- ↑ "WS227 Release Letter". WormBase. 10 August 2011. Arquivado dende o orixinal o 28 de novembro de 2013. Consultado o 2013-11-19.
- ↑ Ruby, JG; Jan, C; Player, C; Axtell, MJ; Lee, W; Nusbaum, C; Ge, H; Bartel, DP (2006). "Large-Scale Sequencing Reveals 21U-RNAs and Additional MicroRNAs and Endogenous siRNAs in C. Elegans". Cell 127 (6): 1193–207. PMID 17174894. doi:10.1016/j.cell.2006.10.040.
- ↑ Stricklin, SL; Griffiths-Jones, S; Eddy, SR (2005). "C. elegans noncoding RNA genes". WormBook. doi:10.1895/wormbook.1.1.1.
- ↑ Stein, LD; et al. (2003). "The Genome Sequence of Caenorhabditis briggsae: A Platform for Comparative Genomics". PLoS Biology 1 (2): 166–192. PMC 261899. PMID 14624247. doi:10.1371/journal.pbio.0000045.
- ↑ Genome Sequencing Center. "Caenorhabditis remanei: Background". Washington University School of Medicine. Arquivado dende o orixinal o 16 de xuño de 2008. Consultado o 2008-07-11.
- ↑ Genome Sequencing Center. "Caenorhabditis japonica: Background". Washington University School of Medicine. Arquivado dende o orixinal o 26 de xuño de 2008. Consultado o 2008-07-11.
- ↑ "C. brenneri (Caenorhabditis brenneri) Genome Browser Gateway". rohsdb.cmb.usc.edu. Consultado o 2020-01-17.
- ↑ "WS202 Release Letter". WormBase. 29 May 2009. Consultado o 17 de xaneiro de 2020.
- ↑ "WS197 Release Letter". WormBase. 27 November 2008. Arquivado dende o orixinal o 17 de outubro de 2019. Consultado o 2013-11-19.
- ↑ "Genome sequence changes". WormBase. 15 June 2011. Arquivado dende o orixinal o 17 de outubro de 2019. Consultado o 2011-08-13.
- ↑ Gamulin, V; Muller, IM; Muller, WEG (2000). "Sponge proteins are more similar to those of Homo sapiens than to Caenorhabditis elegans". Biological Journal of the Linnean Society 71 (4): 821–828. doi:10.1111/j.1095-8312.2000.tb01293.x.
- ↑ Fire, A; Xu, S; Montgomery, MK; Kostas, SA; Driver, SE; Mello, CC (1998). "Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans". Nature 391 (6669): 806–11. Bibcode:1998Natur.391..806F. PMID 9486653. doi:10.1038/35888.
Véxase tamén
[editar | editar a fonte]Bibliografía
[editar | editar a fonte]- Bird, J; Bird, AC (1991). The structure of nematodes. Academic Press. pp. 1, 69–70, 152–153, 165, 224–225. ISBN 0-12-099651-0.
- Hope, IA (1999). C. elegans: a practical approach. Oxford University Press. pp. 1–6. ISBN 0-19-963738-5.
- Riddle, DL; Blumenthal, T; Meyer, RJ; Priess, JR (1997). C. elegans II. Cold Spring Harbor Laboratory Press. pp. 1–4, 679–683. ISBN 0-87969-532-3.
- Rodriguez, M; Snoek, LB; De Bono, M; Kammenga, JE (2013). "Worms under stress: C. elegans stress response and its relevance to complex human disease and aging". Trends in Genetics 29 (6): 367–374. PMID 23428113. doi:10.1016/j.tig.2013.01.010.
Ligazóns externas
[editar | editar a fonte]- Brenner S (2002) Nature's Gift to Science. In. https://web.archive.org/web/20131008213937/http://www.nobelprize.org/nobel_prizes/medicine/laureates/2002/brenner-lecture.pdf
- Horvitz HR (2002) Worms, Life and Death. In. https://web.archive.org/web/20110924061142/http://www.nobelprize.org/nobel_prizes/medicine/laureates/2002/horvitz-lecture.pdf
- Sulston JE (2002) The Cell Lineage and Beyond. In. https://web.archive.org/web/20140201235554/http://www.nobelprize.org/nobel_prizes/medicine/laureates/2002/sulston-lecture.pdf
- WormBase – ampla base de datos en liña sobre a bioloxía e xenómica de C. elegans e outros nematodos
- WormBook – un compendio de acceso libre en liña de todos os aspectos da bioloxía de C. elegans, incluíndo protocolos de laboratorio
- Wormatlas – base de datos en liña sobre o comportamento e anatomía estrutural de C. elegans
- Atlas dixital 3D de C. elegans – un atlas dixital en 3D a resolución dun só núcleo con funcións de procura, análise cuantitativa e ferramentas de visualización 3D para a anatomía estrutural de C. elegans
- Wellcome Trust Sanger Institute C. elegans page – a metade da secuencia xenómica é mantida por este instituto
- AceView WormGenes – outra base de datos xenómica para C. elegans, mantida no NCBI
- TCNJ Worm Lab – Protocolos e imaxes fáciles de seguir de investigacións en C. elegans.
- Worm Classroom – Portal educativo sobre C. elegans
- Textpresso Arquivado 08 de xaneiro de 2014 en Wayback Machine. – Motor de procura da WormBase
- Películas de C elegans feitas por investigadores
- C. elegans II – libro de texto en liña de acceso libre.
- Silencing Genomes Arquivado 10 de agosto de 2019 en Wayback Machine. Experimentos de interferencia de ARN (RNAi) e bioinformática en C. elegans para educación. Do Dolan DNA Learning Center do Cold Spring Harbor Laboratory.
- Modelos 3D de C. elegans do Ewbank Lab – Vídeos e fotos que explican a anatomía básica de C. elegans
- Abundancia de proteínas de C. elegans
- WormWeb.org: Visualización interactiva da rede neural de C. elegans – Conectividade entre as 302 neuronas, coa posibilidade de procurar a vía de conexión máis curta entre dúas neuronas dadas
- WormWeb.org: Visualización interactiva da liñaxe celular de C. elegans – As divisións que dan lugar a cada unha das células do verme
- Vista do xenoma de Caenorhabditis elegans en Ensembl