Célula piramidal
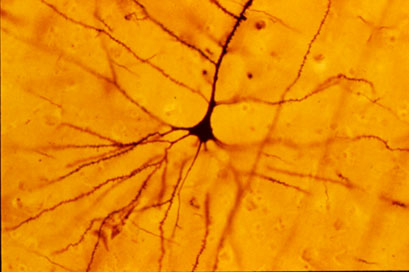

As células piramidais ou neuronas piramidais son un tipo de neurona multipolar que se atopa en áreas do cerebro, como o córtex cerebral, o hipocampo e a amígdala. As células piramidais son as unidades de excitación primarias do córtex prefrontal e tracto corticoespiñal de mamíferos. Son tamén un dos tipos celulares nos que se encontran signos característicos, concretamente corpos de Negri, en infeccións de rabia post-mortem.[1] As neuronas piramidais foron descubertas e estudadas por primeira vez por Santiago Ramón y Cajal.[2][3] Desde entón, os estudos sobre as neuronas piramidais centráronse en campos que van desde a neuroplasticidade á cognición.
Estrutura
[editar | editar a fonte]-
Neurona piramidal visualizada por medio de proteína fluorescente verde (GFP)
-
Unha célula piramidal do hipocampo


Unha das principais características estruturais da neurona piramidal é a forma cónica do seu soma ou corpo celular, polo cal recibe o seu nome. Outras características clave son ter un só axón, unha dendrita apical grande, moitas dendritas basais, e a presenza de espiñas dendríticas.[4]
Dendritas apicais
[editar | editar a fonte]A dendrita apical emerxe do ápice do soma da célula piramidal. É unha soa dendrita longa e grosa que se ramifica varias veces a medida que a distancia desde o soma se incrementa e estende cara á superficie cortical.[4]
Dendritas basais
[editar | editar a fonte]As dendritas basais orixínanse desde a base do soma. A árbore dendrítica basal consta de tres a cinco dendritas primarias. A medida que aumenta a distancia desde o soma, as dendritas basais ramifícanse profusamente.[4]
As células piramidais son unhas das neuronas máis grandes do cerebro. Tanto en humanos coma en roedores, os corpos ou somas das células piramidais teñen como media unha lonxitude duns 20 μm. As dendritas piramidais normalmente teñen un diámetro desde medio micron a varios microns. A lonxitude dunha soa dendrita adoita ser de varios centos de microns. Debido á ramificación, a lonxitude dendrítica total dunha célula piramidal pode acadar varios centímetros. O axón da célula piramidal adoita ser incluso máis longo e extensamente ramificado, chegando a unha lonxitude total de moitos centímetros.
Espiñas dendríticas
[editar | editar a fonte]As espiñas dendríticas reciben a maioría dos impulsos excitatorios (EPSPs) que entran nunha célula piramidal. As espiñas dendríticas foron detectadas primeiramente por Ramón y Cajal en 1888 usando o método de Golgi. Ramón y Cajal foi tamén a primeira persoa en propoñer o papel fisiolóxico do incremento da área superficial receptiva da neurona. Canto maior é a área superficial da célula piramidal, maior é a capacidade da neurona de procesar e integrar grandes cantidades de información. As espiñas dendríticas están ausentes no soma, mentres que o seu número aumenta a medida que nos afastamos del.[3] A dendrita apical típica na rata ten polo menos 3.000 espiñas dendríticas. A dendrita apical humana media é de aproximadamente o dobre de longo que a da rata, así que o número de espiñas dendríticas presentes nunha dendrita apical humana podería ser de ata 6.000.[5]
Crecemento e desenvolvemento
[editar | editar a fonte]Diferenciación
[editar | editar a fonte]A especificación piramidal ocorre durante o desenvolvemento temperán do cerebro. As células proxenitoras están destinadas á liñaxe neuronal na zona ventricular proliferativa subcortical e na zona subventricular. As células piramidais inmaturas migran para ocupar a placa cortical, onde seguen diversificándose. Os endocannabinoides son unha clase de moléculas que dirixen o desenvolvemento das células piramidais e a exploración do camiño durante o crecemento axonal.[6] Factores de transcrición como Ctip2 e Sox5 contribúen a establecer a dirección á cal as células piramidais dirixen os seus axóns.[7]
Desenvolvemento postnatal temperán
[editar | editar a fonte]As células piramidais de ratas sofren moitos cambios rápidos durante a vida postnatal temperá. Entre os días postnatais 3 e 21, as células piramidais duplican o tamaño do seu soma, multiplican por cinco a lonxitude da dendrita apical e por trece a das dendritas basais. Outros cambios son a diminución do potencial de repouso de membrana, a redución da resistencia da membrana e un incremento nos valores pico dos potenciais de acción.[8]
Sinalización
[editar | editar a fonte]Na maioría das outras neuronas, as dendritas son xeralmente as áreas de entrada do impulso, mentres que o axón é a vía de saída. Tanto os axóns coma as dendrits están moi ramificados. A gran cantidade de ramificacións permite que a neurona envíe e reciba sinais a e de diferentes neuronas.
As neuronas piramidais, igual que outras neuronas, teñen numerosas canles iónicas reguladas por voltaxe. Nas células piramidais, hai unha abundancia de canles de Na+, Ca2+ e K+ nas dendritas, e algunhas canles no soma.[9][10] As canles iónicas das dendritas das células piramidais teñen diferentes propiedades comparadas coas canles para o mesmo tipo de ión do seu soma.[11][12] As canles de Ca2+ dependentes de voltaxe das dendritas das células piramidais son activadas por potenciais postsinápticos excitatorios (EPSPs) sublimiares e por poenciais de acción que se retropropagan. O grao de retropropagación dos potenciais de acción en dendritas de células piramidais depende das canles de K+. Estas canles de K+ proporcionan un mecanismo para controlar a amplitude dos potenciais de acción.[13]
A capacidade das neuronas piramidais de integrar a información depende do número e distribución das entradas (inputs) sinápticas que reciben. Unha célula piramidal recibe unhas 30.000 entradas de impulsos excitatorios e 1.700 inhibitorios (IPSPs). As entradas excittorias (EPSPs) terminan exclusivamente nas espiñas dendríticas, mentres que as entradas inhibitorias (IPSPs) terminan nos talos dendríticos, no soma ou mesmo no axón. As neuronas piramidais poden ser excitadas polo neurotransmisor glutamato,[4][14] e inhibidas polo neurotransmisor GABA.[4]
Clasificación polo disparo de impulsos
[editar | editar a fonte]As neuronas piramidais foron clasificadas en varias subclases baseándose nas súas respostas de disparo de impulsos a pulsos de corrente de 400-1000 milisegundos. As subclases son: neuronas RSad, RSna e IB.
RSad
[editar | editar a fonte]As neuronas piramidais RSad ou neuronas con picos regulares adaptativoss, disparan con potenciais de acción individuais que van seguidos dun postpotencial (afterpotential) hiperpolarizante. O postpotencial incrementa a súa duración, o cal crea unha adaptación da frecuencia de picos na neurona.[15]
RSna
[editar | editar a fonte]As neuronas piramidais RSna ou neuronas con picos regulares non adaptativos, disparan un tren de potenciais de acción despois dun pulso. Estas neuronas non mostran signos de adaptación.[15]
IB
[editar | editar a fonte]Neuronas piramidais IB ou neuronas que explotan intrinsecamente, responden a pulsos limiares cunha explosión de dous a cinco potenciais de acción rápidos. As neuronas piramidais IB non mostran adaptación.[15]
Clasificacións moleculares
[editar | editar a fonte]Hai varios estudos que mostran que as propiedades morfolóxicas e eléctricas das células piramidais poderían deducirse da expresión xénica medida por secuenciación dunha soa célula.[16] Varios estudos propoñen que a clasificación de células individuais de neuronas de ratos[17] e humanas[18] baseándose na expresión xénica podería explicar varias propiedades neuronais . Os tipos neuronais destas clasificacións divídense en excitatorios, inhibitorios e centos de subtipos. Por exemplo, as células piramidais da capa 2-3 en humanos clasifícanse como de tipo FREM3[16] e adoitan ter gran cantidade de corrente Ih[19] xerada por canles de HCN.
Función
[editar | editar a fonte]Tracto corticoespiñal
[editar | editar a fonte]As neuronas piramidais son o principal tipo neural no tracto corticoespiñal. O control motor normal depende do desenvolvemento de conexións entre os axóns do tracto corticoespiñal e a medula espiñal. Os axóns das células piramidais seguen pistas como os factores de crecemento para facer conexións específicas. Coas conexións correctas, as células piramidais poden tomar parte na circuitería responsable da visión da función motora guiada.[20]
Cognición
[editar | editar a fonte]As neuronas piramidais do córtex prefrontal interveñen na capacidade cognitiva. En mamíferos a complexidade das células piramidais increméntase desde a rexión cerebral posterior á anterior. O grao de complexidade das neuronas piramidais está probablemente ligado ás capacidades cognitivas de diferentes especies de antropoides. As céllas piramidais do córtex prefrontal parecen ser responsables de procesar entradas (inputs) do córtex auditivo primario, o córtex somatosensorial primario e o córtex visual primario, todas as cales procesan modalidades sensoriais.[21] Estas células poderían tamén xogar un papel crítico no recoñecemento de obxectos complexos dentro das áreas de procesamento visual do córtex.[2] En relación con outras especies, o maior tamaño celular e complexidde das neuronas piramidais, xunto con certos padróns de organización celular e función, correlaciónanse coa evolución da cognición humana. [22]
Memoria e aprendizaxe
[editar | editar a fonte]As células piramidais do hipocampo son esenciais para certos tipos de memoria e aprendizaxe. Forman sinapses que axudan na integración das voltaxes sinápticas nas súas árbores dendríticas complexas polas interaccións con fibras musgosas das células gránulo. Como isto afecta ás voltaxes postsinápticas producidas pola activación de fibras musgosas, a situación das excrecencias espiñosas nas dendritas basais e apical é importante para a formación da memoria. Ao establecer un control dinámico da sensibilidade das células piramidais CA3, este agrupamento das sinapses das fibras musgosas nas células piramidais pode facilitar a iniciación dos picos somáticos. As interaccións entre as células piramidais e uns 41 botóns de fibras musgosas estimados, cada un orixinado a partir dunha única célula gránulo, salientan o papel destes botóns no procesamento da información e conectividade sináptica, os cales son esenciais para a memoria e a aprendizaxe. Fundamentalmente, os impulsos enviados polas fibras musgosas recíbenos as células piramidais do hipocampo, o cal integra as voltaxes sinápticas dentro da arquitectura dendrítica. A localización de protrusións espiñosas e o agrupamento de sinapses inflúen na sensibilidade e contribúen ao procesamento de información pertencente á memoria e aprendizaxe.[23]
Notas
[editar | editar a fonte]- ↑ Sketchy Group, LLC. "2.3 rhabdovirus". SketchyMedical. Arquivado dende o orixinal o 2017-04-13.
- ↑ 2,0 2,1 Elston GN (novembro de 2003). "Cortex, cognition and the cell: new insights into the pyramidal neuron and prefrontal function". Cerebral Cortex 13 (11): 1124–1138. PMID 14576205. doi:10.1093/cercor/bhg093.
- ↑ 3,0 3,1 García-López P, García-Marín V, Freire M (novembro de 2006). "Three-dimensional reconstruction and quantitative study of a pyramidal cell of a Cajal histological preparation". The Journal of Neuroscience 26 (44): 11249–11252. PMC 6674523. PMID 17079652. doi:10.1523/JNEUROSCI.3543-06.2006.
- ↑ 4,0 4,1 4,2 4,3 4,4 Megías M, Emri Z, Freund TF, Gulyás AI (2001). "Total number and distribution of inhibitory and excitatory synapses on hippocampal CA1 pyramidal cells". Neuroscience 102 (3): 527–540. PMID 11226691. doi:10.1016/S0306-4522(00)00496-6.
- ↑ Laberge D, Kasevich R (novembro de 2007). "The apical dendrite theory of consciousness". Neural Networks 20 (9): 1004–1020. PMID 17920812. doi:10.1016/j.neunet.2007.09.006.
- ↑ Mulder J, Aguado T, Keimpema E, Barabás K, Ballester Rosado CJ, Nguyen L, et al. (xuño de 2008). "Endocannabinoid signaling controls pyramidal cell specification and long-range axon patterning". Proceedings of the National Academy of Sciences of the United States of America 105 (25): 8760–8765. Bibcode:2008PNAS..105.8760M. PMC 2438381. PMID 18562289. doi:10.1073/pnas.0803545105.
- ↑ Fishell G, Hanashima C (febreiro de 2008). "Pyramidal neurons grow up and change their mind". Neuron 57 (3): 333–338. PMID 18255026. doi:10.1016/j.neuron.2008.01.018.
- ↑ Zhang ZW (marzo de 2004). "Maturation of layer V pyramidal neurons in the rat prefrontal cortex: intrinsic properties and synaptic function". Journal of Neurophysiology 91 (3): 1171–1182. PMID 14602839. doi:10.1152/jn.00855.2003.
- ↑ Spruston N (marzo de 2008). "Pyramidal neurons: dendritic structure and synaptic integration". Nature Reviews. Neuroscience 9 (3): 206–221. PMID 18270515. doi:10.1038/nrn2286.
- ↑ Georgiev DD, Kolev SK, Cohen E, Glazebrook JF (decembro de 2020). "Computational capacity of pyramidal neurons in the cerebral cortex". Brain Research 1748: 147069. PMID 32858030. arXiv:2009.10615. doi:10.1016/j.brainres.2020.147069.
- ↑ Golding NL, Mickus TJ, Katz Y, Kath WL, Spruston N (outubro de 2005). "Factors mediating powerful voltage attenuation along CA1 pyramidal neuron dendrites". The Journal of Physiology 568 (Pt 1): 69–82. PMC 1474764. PMID 16002454. doi:10.1113/jphysiol.2005.086793.
- ↑ Remy S, Beck H, Yaari Y (agosto de 2010). "Plasticity of voltage-gated ion channels in pyramidal cell dendrites". Current Opinion in Neurobiology 20 (4): 503–509. PMID 20691582. doi:10.1016/j.conb.2010.06.006.
- ↑ Magee J, Hoffman D, Colbert C, Johnston D (1998). "Electrical and calcium signaling in dendrites of hippocampal pyramidal neurons". Annual Review of Physiology 60 (1): 327–346. PMID 9558467. doi:10.1146/annurev.physiol.60.1.327.
- ↑ Wong, R. K. S.; Traub, R. D. (2009-01-01). Schwartzkroin, Philip A., ed. NETWORKS | Cellular Properties and Synaptic Connectivity of CA3 Pyramidal Cells: Mechanisms for Epileptic Synchronization and Epileptogenesis. Encyclopedia of Basic Epilepsy Research (en inglés) (Oxford: Academic Press). pp. 815–819. ISBN 978-0-12-373961-2. doi:10.1016/b978-012373961-2.00215-0. Consultado o 2020-11-18.
- ↑ 15,0 15,1 15,2 Franceschetti S, Sancini G, Panzica F, Radici C, Avanzini G (abril de 1998). "Postnatal differentiation of firing properties and morphological characteristics in layer V pyramidal neurons of the sensorimotor cortex". Neuroscience 83 (4): 1013–1024. PMID 9502243. doi:10.1016/S0306-4522(97)00463-6.
- ↑ 16,0 16,1 Berg J, Sorensen SA, Ting JT, Miller JA, Chartrand T, Buchin A, et al. (outubro de 2021). "Human neocortical expansion involves glutamatergic neuron diversification". Nature 598 (7879): 151–158. Bibcode:2021Natur.598..151B. PMC 8494638. PMID 34616067. doi:10.1038/s41586-021-03813-8.
- ↑ Gouwens NW, Sorensen SA, Berg J, Lee C, Jarsky T, Ting J, et al. (xullo de 2019). "Classification of electrophysiological and morphological neuron types in the mouse visual cortex". Nature Neuroscience 22 (7): 1182–1195. PMC 8078853. PMID 31209381. doi:10.1038/s41593-019-0417-0.
- ↑ Bakken TE, Jorstad NL, Hu Q, Lake BB, Tian W, Kalmbach BE, et al. (outubro de 2021). "Comparative cellular analysis of motor cortex in human, marmoset and mouse". Nature 598 (7879): 111–119. Bibcode:2021Natur.598..111B. PMC 8494640. PMID 34616062. doi:10.1038/s41586-021-03465-8.
- ↑ Kalmbach BE, Buchin A, Long B, Close J, Nandi A, Miller JA, et al. (decembro de 2018). "h-Channels Contribute to Divergent Intrinsic Membrane Properties of Supragranular Pyramidal Neurons in Human versus Mouse Cerebral Cortex". Neuron 100 (5): 1194–1208.e5. PMC 6447369. PMID 30392798. doi:10.1016/j.neuron.2018.10.012.
- ↑ Salimi I, Friel KM, Martin JH (xullo de 2008). "Pyramidal tract stimulation restores normal corticospinal tract connections and visuomotor skill after early postnatal motor cortex activity blockade". The Journal of Neuroscience 28 (29): 7426–7434. PMC 2567132. PMID 18632946. doi:10.1523/JNEUROSCI.1078-08.2008.
- ↑ Baker A, Kalmbach B, Morishima M, Kim J, Juavinett A, Li N, Dembrow N (xuño de 2018). "Specialized Subpopulations of Deep-Layer Pyramidal Neurons in the Neocortex: Bridging Cellular Properties to Functional Consequences". The Journal of Neuroscience 38 (24): 5441–5455. PMC 6001033. PMID 29798890. doi:10.1523/JNEUROSCI.0150-18.2018.
- ↑ Galakhova AA, Hunt S, Wilbers R, Heyer DB, de Kock CP, Mansvelder HD, Goriounova NA (novembro de 2022). "Evolution of cortical neurons supporting human cognition". Trends in Cognitive Sciences (en English) 26 (11): 909–922. PMC 9561064. PMID 36117080. doi:10.1016/j.tics.2022.08.012.
- ↑ Gonzales, R. B.; DeLeon Galvan, C. J.; Rangel, Y. M.; Claiborne, B. J. (2001-02-12). "Distribution of thorny excrescences on CA3 pyramidal neurons in the rat hippocampus". The Journal of Comparative Neurology 430 (3): 357–368. ISSN 0021-9967. PMID 11169473. doi:10.1002/1096-9861(20010212)430:3<357::aid-cne1036>3.0.co;2-k.
Véxase tamén
[editar | editar a fonte]Outros artigos
[editar | editar a fonte]- Córtex cerebral
- Tracto piramidal
- Célula candelabro - innervan os segmentos iniciais dos axóns piramidais
- Neurona de Rosehip