Axón
| Estrutura dunha neurona típica |
|---|
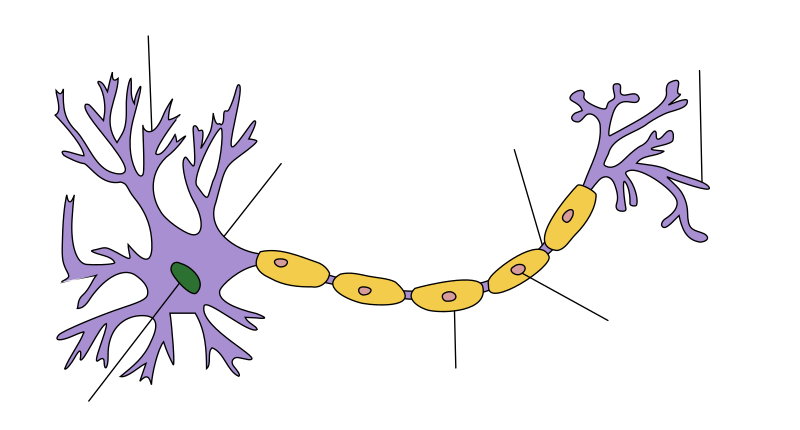
O axón[1][2][3][4] ou axon[5] é a prolongación longa e delgada que presentan as neuronas, que conduce os potenciais eléctricos do impulso nervioso fóra do soma da neurona. Tamén se chama cilindroeixe.[6]
O axón é un dos dous tipos de prolongacións que presentan as células nerviosas, o outro son as dendritas. Os axóns distínguense das dendritas en varios aspectos, como a forma (as dendritas xeralmente se van adelgazando cara ao seu extremo, mentres que o axón mantén case constante o seu diámetro), lonxitude (as dendritas son menos longas), ramificación (as dendritas xeralmente están máis ramificadas, pero os axóns dalgunhas neuronas poden ter tamén bastantes ramificacións), e función (o normal é que as dendritas reciban os sinais e os axóns os transmitan). Pero hai excepcións para cada un dos casos.
Algúns tipos de neuronas non teñen axón (células amácrinas da retina) e transmiten sinais polas súas dendritas. As neuronas teñen só un axón, pero en invertebrados (insectos, sambesugas) o axón consta ás veces de varias rexións que funcionan máis ou menos independentemente unhas das outras.[7] A maioría dos axóns ramifícanse, nalgúns casos moi profusamente. O conxunto de ramificacións do axón chámase telodendron. O final das ramificacións están os botóns sinápticos, zonas máis alargadas onde se realiza a sinapse.
Os axóns fan contacto con outras células, normalmente outras neuronas ou ás veces células musculares ou glandulares, por medio de sinapses. Nunha sinapse a membrana do axón se achega moi preto da membrana da outra célula e transmítelle sinais electroquímicas a través do espazo entre elas, xeralmente liberando unha substancia chamada neurotransmisor. Algunhas unións sinápticas están situadas ao final do axón e outras vanse formando ao longo da súa lonxitude. Un só axón con todas as súas ramificacións pode establecer miles de sinapses.
Características
[editar | editar a fonte]O axón é unha prolongación longa e delgada que se orixina nunha rexión especializada da neurona chamada cono axónico ou protuberancia ou eminencia axónica, ou ás veces nunha dendrita principal. O axón ten a forma dun cono que se adelgaza cara a periferia, moi pouco a pouco, polo que case mantén o seu diámetro en longas extensións. Na súa superficie obsérvanse constricións circulares periódicas chamadas nódulos de Ranvier. A membrana celular do axón recibe o nome de axolema.
O axoplasma é o citoplasma contido dentro do axón e do cono axónico. Contén microtúbulos, neurofilamentos, mitocondrias, gránulos e vesículas, e diferénciase do citoplasma do soma e das dendritas proximais, porque carece de ribosomas libres, corpos de Nissl e de aparato de Golgi.
Os axóns poden estar cubertos ou non por unha vaíña protectora denominada vaíña de mielina. No sistema nervioso periférico os axóns están sempre cubertos polas células de Schwann, que rodean o axón cunha capa múltiple formada a partir da membrana destas células e constitúen a vaíña de mielina. As neuronas do sistema nervioso periférico que non teñen vaíña de mielina están embutidas en células de Schwann, conformando o feixe de Remak. No sistema nervioso central os axóns que están mielinizados están cubertos polos oligodendrocitos, células da glía xunto con células de Schwann que forman a vaíña de mielina.
As interrupcións da vaíña de mielina chámanse nódulos de Ranvier, que aparecen a intervalos regularmente espazados. A mielinación illa electricamente o axón excepto nos nódulos de Ranvier e permite un modo rápido de propagación do impulso eléctrico chamado condución saltatoria. Nos nódulos de Ranvier (zonas desmielinizadas) os potenciais de acción son amplificados e transmitidos con gran rapidez. A perda da vaíña de mielina pode causar severos trastornos como a esclerose múltiple.
O segmento inicial do axón está tamén desmielinizado e considérase que é o lugar onde se inician os potenciais de acción do impulso nervioso.[8] Nese segmento a densidade de canles de sodio reguladas por voltaxe é moito maior ca no corpo celular, excepto no cono axónico.[9]
Tipos de neuronas segundo a lonxitude do axón
[editar | editar a fonte]O diámetro típico do axón é de 1 μm, pero a súa lonxitude pode ser considerable. Algúns son de poucos mm, pero o máis longo, o dunha célula do nervio ciático, que vai da medula espiñal á punta dos pés, pode ter máis dun metro.[10]
As neuronas pódense clasificar en dous tipos de acordo coa lonxitude do seu axón:
- Neuronas Golgi tipo I: Posúen un axón longo que pode chegar a medir máis dun metro. Por exemplo, as células de Purkinje.
- Neuronas Golgi tipo II: Posúen un axón curto, similar a unha dendrita que termina non moi lonxe do soma. Por exemplo, as células granulares.
Fisioloxía
[editar | editar a fonte]A fisioloxía do axón pode describirse co modelo de Hodgkin-Huxley, estendido aos vertebrados polas ecuacións de Frankenhaeuser-Huxley. As fibras nerviosas periféricas poden clasificarse baseándose na velocidade de condución do axón, mielinización, tamaño da fibra etc. Por exemplo, hai fibras C amielínicas de condución lenta e fibras delta A mielínicas de condución rápida. Hai varios tipos de fibras sensoriais e de fibras motoras. Outras fibras son as do sistema nervioso autónomo.
Nas táboas seguintes indícanse os tipos e características principais:
Fibras motoras
[editar | editar a fonte]As motoneuronas que se dirixen da medula espiñal e encéfalo aos músculos poden ter dous tipos de fibras:
| Tipo | Clasificación de Erlanger-Gasser |
Diámetro | Mielina | Velocidade de condución | fibras musculares asociadas |
|---|---|---|---|---|---|
| α | Aα | 13-20 µm | Si | 80–120 m/s | Fibras musculares extrafusais |
| γ | Aγ | 5-8 µm | Si | 4–24 m/s[11][12] | Fibras musculares intrafusais |
Fibras sensoriais
[editar | editar a fonte]Os diferentes receptores sensoriais están innervados por diferentes tipos de fibras nerviosas. Os propioceptores están innervados polos tipos Ia, Ib e II, os mecanoreceptores polos tipos II e III, e os nociceptores e termorreceptores polos tipos III e IV de fibras sensoriais.
| Tipo | Clasificación de Erlanger-Gasser |
Diámetro | Mielina | Velocidade de condución | receptor sensorial asociado |
|---|---|---|---|---|---|
| Ia | Aα | 13-20 µm | Si | 80–120 m/s | Receptores primarios do fuso muscular |
| Ib | Aα | 13-20 µm | Si | 80–120 m/s | órgano tendinoso de Golgi |
| II | Aβ | 6-12 µm | Si | 33–75 m/s | Receptores secundarios do fuso muscular Todos os mecanorreceptores cutáneos |
| III | Aδ | 1-5 µm | Delgada | 3–30 m/s | Terminacións nerviosas libres do tacto e presión Nociceptores do tracto neoespiñotalámico termoreceptores do frío |
| IV | C | 0.2-1.5 µm | Non | 0.5-2.0 m/s | Nociceptores do tracto paleoespiñotalámico receptores da calor |
Fibras autónomas
[editar | editar a fonte]O Sistema nervioso autónomo presenta dous tipos de fibras periféricas:
| Tipo | Clasificación de Erlanger-Gasser |
Diámetro | Mielina[13] | Velocidade de condución |
|---|---|---|---|---|
| fibras preganglionares | B | 1-5 µm | Si | 3–15 m/s |
| fibras postganglionares | C | 0.2-1.5 µm | Non | 0.5-2.0 m/s |
Funcións do axón
[editar | editar a fonte]O axón propaga o impulso nervioso, crece conectando a neurona con outras células e serve como vía de transporte de diversos elementos entre o soma e o extremo axónico da neurona.
Transporte de orgánulos e substancias
[editar | editar a fonte]O transporte de orgánulos, encimas, macromoléculas e metabolitos, é unha función do axoplasma na que interveñen directamente os microtúbulos. O transporte axoplásmico é necesario para o mantemento do axón e das células asociadas a el, e para permitir a chegada ao pericarion de factores reguladores que regulan a súa función.
O transporte polo interior do axón pode facerse en dúas direccións:
- Transporte anterógrado ou centrífugo: Desde o soma neuronal cara aos terminais axónicos.
- Transporte retrógrado ou centrípeto: Desde os terminais axónicos cara ao soma neuronal.
A velocidade deste transporte varía entre:
- Fluxo lento de 0,5 µm/min, velocidade á que se desprazan agregados moleculares como as subunidades proteicas que forman o citoesqueleto axonal.
- Fluxo rápido anterógrado ao que os orgánulos se desprazan a velocidades duns 300 µm/min. Unha molécula de cinesina, unida a un receptor na membrana do orgánulo transportado (unha vesícula por exemplo) desprázase, con gasto de ATP, desde o extremo negativo do microtúbulo, situado no pericarion cara ao seu extremo positivo.
- Fluxo rápido retrógrado ao que as vesículas membranosas procedentes dos botóns terminais do axón, son transportadas cara ao pericarion a uns 200 µm/min. A molécula de dineína citoplasmática (MAP1C) unida a un receptor na membrana do orgánulo transportado desprázase interactuando coa tubulina con gasto de ATP, desde o extremo positivo do microtúbulo, situado no terminal axónico cara ao seu extremo negativo.
Condución do impulso nervioso
[editar | editar a fonte]

Os axóns constitúen as fibras nerviosas funcionando como a rama eferente, que transmite, o potencial de acción, excitatorio ou inhibitorio, a través dunha ou máis sinapses. Os axóns tamén poden recibir impulsos a través de sinapsis axoaxónicas, que se realizan entre dous axóns, pero as función de envío de sinais de saída é a predominante nos axóns.
A condución do impulso nervioso non é outra cousa ca o desprazamento do potencial de acción xerado por cambios na permeabilidade a ións ao lango da membrana do axón (axolema) das fibras nerviosas, na que axudan as células de sostén que rodean como unha vaíña o axón.
No sistema nervioso central os axóns están rodeados pola mielina dos oligodendrocitos, mentres que no sistema nervioso periférico poden estar rodeados, xa sexa, por prolongacións citoplasmáticas das células de Schwann (fibras amielínicas) ou pola mielina das células de Schwann (fibras nerviosas mielínicas do sistema nervioso periférico).
Os impulsos nerviosos son ondas transitorias de inversión da voltaxe que existe na membrana plasmática, que se inician no lugar en que se produce o estímulo. Cada unha destas ondas corresponde a un potencial de acción.
Este proceso é posible grazas ás macromoléculas que son proteínas integrais de membrana ou complexos de proteínas, e ocupan todo o espesor do axolema. Entre elas:
- A bomba de sodio-potasio, que pode transportar activamente sodio cara ao medio extracelular intercambiándoo por potasio.
- Canles para sodio de apertura por voltaxe, que determinan a inversión da voltaxe da membrana xa que ao abrírense e permitiren a entrada de sodio fan que o interior da membrana se volva positivo.
- Canles para potasio de apertura por voltaxe, cuxa activación contribúe a recuperar a polaridade inicial, por saída de ións potasio desde o interior do axoplasma.
Nas fibras nerviosas amielínicas o impulso condúcese cara aos botóns terminais dos axóns como unha onda continua de inversión de voltaxe, a unha velocidade que é proporcional ao diámetro do axón, e varía de 1 a 100 metros por segundo.
Nas fibras nerviosas mielínicas, o axón está cuberto por unha vaíña de mielina formada pola superposición ou enrolamento dunha serie de capas de membrana celular, que actúa como un illante eléctrico do axón. Ao longo do axón, a mielina está formada por células sucesivas e en cada límite intercelular existe un anel sen mielina que corresponde ao nódulo de Ranvier.
Nos nódulos de Ranvier prodúcese o fluxo de ións a través da membrana axonal. O axolema dos nodos de Ranvier ten unha alta concentración de canles de sodio sensibles a voltaxe. A consecuencia é unha condución saltatoria do potencial de acción xa que a inversión da voltaxe inducida nun nódulo de Ranvier continúase por propagación pasiva rápida da corrente polo interior do axón e polo medio extracelular ata o nódulo seguinte, onde produce a inversión da voltaxe.
A consecuencia desta estrutura é que nos axóns mielínicos a condución do impulso nervioso é máis rápida. A velocidade de condución do impulso nervioso é proporcional ao diámetro do axón e á distancia entre os nódulos de Ranvier nos axóns mielínicos.
A primeira medición da velocidade do impulso nervioso atribúese a Hermann von Helmholtz [1], que en 1853 estableceu un valor medio de 27,25 m/s.
Crecemento e desenvolvemento do axón
[editar | editar a fonte]
Os axóns en crecemento crecen polos tecidos a partir do cono de crecemento que teñen no seu extremo. O cono de crecemento ten largas expansións chamadas lamelipodios das que saen outras máis finas chamadas filopodios. Os filopodios son un mecanismo polo cal o axón se adhire a superficies e explora a área que o rodea. A proteína actina xoga un importante papel na mobilidade deste sistema. As áreas con altos niveis de moléculas de adhesión celular ou CAM facilitan o crecemento axonal. Estas moléculas parecen proporcionar unha superficie "pegañenta" sobre a que os axóns crecen. Exemplos de moléculas de adhesión celular específicas dos sistemas neurais son as CAM neurais ou N-CAM, CAM neurogliais ou NgCAM, as TAG-1, as MAG, e as diciclohexilcarbodiimidas ou DCC, todas as cales forman parte da superfamilia das immunoglobulinas. Outro conxunto de moléculas chamadas moléculas de adhesión á matriz extracelular tamén proporcionan un substrato adherente para que os axóns crezan pola matriz extracelular. Exemplos destas moléculas son as lamininas, a fibronectina, a tenascina, e o perlecán. Algunhas destas están unidas ás superficies celulares e actúan como atractores ou repelentes de curto alcance. Outras son ligandos difusibles con efectos de longo alcance.
Tamén existen as células chamadas célula "fito" (guidepost cells), que xeralmente son outras neuronas inmaturas e células gliais, que axudan a dirixir o crecemento do axón.
Notas
[editar | editar a fonte]- ↑ Definicións no Dicionario da Real Academia Galega e no Portal das Palabras para axón.
- ↑ Dicionario de Bioloxía glego-castelán-inglés. Xunta. 2010. Páxina 28
- ↑ Vocabulario de Medicina. Servizo de Normalización Lingüística. USC. (2008) axón, axon Páxina 76.
- ↑ Vocabulario de Morfoloxía, anatomía e citoloxía veterinaria. Servizo de Normalización Lingüística. USC. (2008). axón, axon
- ↑ Luís Daviña Facal (2000): Diccionario das ciencias da natureza e da saúde. Tomo 1. A-B. A Coruña, Deputación da Coruña, páx. 501. ISBN 84-95335-45-X
- ↑ BUSCatermos cilindroeixe - axón
- ↑ Yau, K.-W. (1976) Receptive fields, geometry and conduction block of sensory neurones in the CNS of the leech. J. Physiol. (Lond) 263:513-538.
- ↑ Clark, Brian D.; Goldberg, Ethan M.; Rudy, Bernardo (2009-12). "Electrogenic Tuning of the Axon Initial Segment". The Neuroscientist (en inglés) 15 (6): 651–668. ISSN 1073-8584. PMC 2951114. PMID 20007821. doi:10.1177/1073858409341973.
- ↑ Wollner D. and Catterall WA (November, 1986). "Localization of sodium channels in axon hillocks and initial segments of retinal ganglion cells". Proc. Nati. Acad. Sci. USA 83: 8424–28. PMC 386941. PMID 2430289. doi:10.1073/pnas.83.21.8424. Consultado o 2009-05-02.
- ↑ DNA From The Beginning, section 6: Genes are real things., "Amination" section, final slide
- ↑ Andrew BL, Part NJ (1972) Properties of fast and slow motor units in hind limb and tail muscles of the rat. Q J Exp Physiol Cogn Med Sci 57:213-225.
- ↑ Russell NJ (1980). "Axonal conduction velocity changes following muscle tenotomy or deafferentation during development in the rat". J Physiol 298: 347–360.
- ↑ pp.187-9 ISBN 0-19-858527-6