Génétique
Le mot génétique vient de l'adjectif grec γενετικός / genetikós, qui qualifie « ce qui est en rapport aux fonctions de génération »[1]. Il dérive du grec γενέτης / genétês, « géniteur ».

On trouve également comme étymologie le grec genos (race, clan) : la partie de la biologie qui étudie les lois de l'hérédité.
Une de ses branches, la génétique formelle, ou mendélienne, s'intéresse à la transmission des caractères héréditaires entre des géniteurs et leur descendance.
L'invention du terme « génétique » revient au biologiste anglais William Bateson (1861-1926), qui l'utilise pour la première fois en 1905. La génétique moderne est souvent datée de la mise en évidence de la structure en double hélice de l'ADN effectuée par James Dewey Watson et Francis Crick en 1953[2].
Histoire
modifierLa génétique est l’étude des gènes et de l'hérédité. Une grande communauté de chercheurs étudiait l’hérédité en Europe mais ce n’est qu’en 1906, quand William Bateson utilisa le terme génétique, que l’institution fut créée.
Le sujet était en expansion depuis 1900. Hugo de Vries, Carl Correns et Erich von Tschermak-Seysenegg avaient chacun redécouvert les lois de Mendel indépendamment. En 1902, Bateson avait montré que les lois de Mendel s’appliquaient aussi aux animaux et non pas seulement aux plantes.
Gregor Mendel avait déjà précisé dans son mémoire de 1866 que les caractéristiques d’un organisme étaient transmises par quelque chose de physique. Dans les deux dernières décennies du XIXe siècle, les chromosomes et leur séparation pendant la mitose et la méiose commençaient à être bien connus. En 1902, Theodor Boveri et Walter Sutton proposèrent que les chromosomes sont le support d’information des caractéristiques des individus. Cette théorie n’était pas très acceptée par la communauté scientifique de l'époque y compris par Bateson. En 1915, Thomas Hunt Morgan publia son livre The Mechanism of Mendelian Heredity qui associa définitivement la génétique avec les chromosomes. Cependant, le mécanisme qui lie l'information contenue sur le chromosome et les caractéristiques que possèdent un individu n'était pas connues.
En 1941, George Beadle et Edward Lawrie Tatum publient l’article « Genetic control of Biochemical reaction in neurospora » qui donna le premier exemple de réaction chimique contrôlée par un gène. Ils avaient compris que les chromosomes avaient un rôle, mais étaient-ils les catalyseurs des réactions ? C’est en 1960 que le débat est tranché par Seymour Benzer et que le gène est reconnu comme support d’informations pour les protéines.
S'ensuit une série de découvertes sur les gènes, l'épissage, le sens de lecture, les opérons etc. qui remettent en cause la définition du gène. De plus l'arrivée de l'épigénétique pose encore plus de questions sur l'héritabilité des caractéristiques.
Différents champs de recherche
modifierTrès tôt, la génétique s'est diversifiée en plusieurs branches :
- la génétique du développement étudie les acteurs moléculaires (et les gènes qui les codent) impliqués dans la formation de l'organisme à partir du stade unicellulaire d'œuf fécondé. Elle se focalise tout particulièrement sur la mise en place de la symétrie bilatérale et les mécanismes qui permettent de passer d'un système biologique simple (unicellulaire, symétrie radiaire) à un organisme complexe (pluricellulaire, souvent métamérisé, et construit en organes spécialisés). Elle utilise souvent des espèces modèles pour étudier les mécanismes de formation de l'organisme (la drosophile, le nématode Caenorhabditis elegans, le poisson-zèbre, une plante du genre Arabidopsis) ;
- la génétique médicale étudie l'hérédité des maladies génétiques humaines, leur ségrégation dans les familles de malades. Elle cherche à identifier par ce biais les mutations responsables des maladies, afin de mettre au point des traitements pour les soigner ;
- la génomique étudie la structure, la composition et l'évolution des génomes (la totalité de l'ADN, trois milliards de paires de bases chez l'être humain, organisées en chromosomes), et tente d'identifier des motifs dans l'ADN pouvant avoir un sens biologique (gènes, unités transcrites non traduites, miRNAs, unités de régulations, promoteurs, CNGs, etc.) ;
- la génétique quantitative étudie la composante génétique expliquant la variation de caractères quantitatifs (la taille, la couleur du pelage, la vitesse de croissance, la concentration d'une molécule, etc.) et leur héritabilité ;
- la génétique de l'évolution étudie les signatures de la sélection naturelle sur le génome des espèces, et tente d'identifier les gènes qui ont joué un rôle essentiel dans l'adaptation et la survie des espèces dans des environnements changeants ;
- la génétique des populations étudie les forces (et leurs effets) qui influencent la diversité génétique des populations[3] et des espèces (mutation, dérive, sélection) par (entre autres) le développement de modèles mathématiques et statistiques.
- la génétique chronologique étudie l'âge de la séparation des espèces en se fiant à la différence génétique entre elles et à la vitesse d'augmentation de la différence génétique, calibrée par d'autres méthodes de chronologie, du groupe d'espèces dont elles font partie.
L'hérédité, qui étudie le phénotype et tente de déterminer le génotype sous-jacent se fonde toujours sur les lois de Mendel. La biologie cellulaire et la biologie moléculaire étudient les gènes et leur support matériel (ADN ou ARN) au sein de la cellule, la biologie cellulaire pour leur expression. Les progrès de la branche ingénierie de la génétique, le génie génétique, ont permis de passer le stade de la simple étude en réussissant à modifier le génome, à implanter, supprimer ou modifier de nouveaux gènes dans des organismes vivants : il s'agit des organismes génétiquement modifiés (OGM). Les mêmes progrès ont ouvert une nouvelle voie d'approche thérapeutique : la « thérapie génique ». Il s'agit d'introduire de nouveaux gènes dans l'organisme afin de pallier une déficience héréditaire.
L'évolution sans cesse croissante de la connaissance en génétique pose plusieurs problèmes éthiques liés au clonage, aux divers types d'eugénismes possibles, à la propriété intellectuelle de gènes et aux possibles risques environnementaux dus aux OGM. La compréhension du fonctionnement de la machinerie cellulaire est ainsi rendue plus complexe : en effet, plus on l'étudie, plus les acteurs sont nombreux (ADN, ARN messager, de transfert, microARN, etc.) et le nombre de rétro-actions (épissage, édition, etc.) entre ces acteurs grandit.
Chronologie
modifierEn 1862, Charles Naudin est primé par l'Académie des sciences pour son Mémoire sur les hybrides du règne végétal.
En 1865, passionné de sciences naturelles, le moine autrichien Gregor Mendel, dans le jardin de la cour de son monastère, décide de travailler sur des pois comestibles présentant sept caractères (forme et couleur de la graine, couleur de l'enveloppe, etc.), dont chacun peut se retrouver sous deux formes différentes. À partir de ses expériences, il publie, en 1866 sous l'autorité de la Société des sciences naturelles de Brünn, un article où il énonce les lois de transmission de certains caractères héréditaires. Cet article, « Recherche sur les hybrides végétaux », est envoyé aux scientifiques des quatre coins du monde : les réactions sont mitigées, voire inexistantes. Ce n'est qu'en 1907 que son article fut reconnu et traduit en français[4].
En 1869, l'ADN est isolé par Friedrich Miescher, un médecin suisse. Il récupère les bandages ayant servi à soigner des plaies infectées et il isole une substance riche en phosphore dans le pus. Il nomme cette substance nucléine. Il trouve la nucléine dans toutes les cellules et dans le sperme de saumon.
En 1879, Walther Flemming décrit pour la première fois une mitose. La mitose avait déjà été décrite 40 ans avant par Carl Nageli mais celui-ci avait interprété la mitose comme une anomalie. Walter Flemming invente les termes prophase, métaphase, et anaphase pour décrire la division cellulaire. Son travail est publié en 1882.
En 1880, Oskar Hertwig et Eduard Strasburger découvrent que la fusion du noyau de l'ovule et du spermatozoïde est l'élément essentiel de la fécondation.
En 1891, Theodor Boveri démontre et affirme que les chromosomes sont indispensables à la vie.
En 1900, redécouverte des lois de l'hérédité : Hugo de Vries, Carl Correns et Erich von Tschermak-Seysenegg redécouvrent de façon indépendante les lois de Mendel.
En 1902, Walter Sutton observe pour la première fois une méiose, propose la théorie chromosomique de l'hérédité, c'est-à-dire que les chromosomes seraient les supports des gènes. Il remarque que le modèle de séparation des chromosomes supporte tout à fait la théorie de Mendel. Il publie son travail la même année[5]. Sa théorie sera démontrée par les travaux de Thomas Morgan. Première description d'une maladie humaine héréditaire par Archibald Garrod : l'alcaptonurie[6].
En 1909, Wilhelm Johannsen crée le terme gène et fait la différence entre l'aspect d'un être (phénotype) et son gène (génotype). William Bateson, quatre ans avant, utilisait le terme génétique dans un article et la nécessité de nommer les variations héréditaires.
En 1911, Thomas Morgan démontre l'existence de mutations en conduisant des expériences sur des drosophiles mutantes aux yeux blancs (mouches du vinaigre). Il montre que les chromosomes sont les supports des gènes, grâce à la découverte des liaisons génétiques (genetic linkage) et des recombinaisons génétiques. Il travaille avec Alfred Sturtevant, Hermann Muller, et Calvin Bridges[7]. Il reçoit le prix Nobel de médecine en 1933. Ses expériences permettront de consolider la théorie chromosomique de l'hérédité.
En 1913, Morgan et Alfred Sturtevant publient la première carte génétique du chromosome X de la drosophile, montrant l'ordre et la succession des gènes le long du chromosome.
En 1928, Fred Griffith découvre la transformation génétique des bactéries, grâce à des expériences sur le pneumocoque. La transformation permet un transfert d'information génétique entre deux cellules. Il ne connaît pas la nature de ce principe transformant.
En 1941, George Beadle et Edward Tatum émettent l'hypothèse qu'un gène code une (et uniquement une) enzyme en étudiant Neurospora crassa[8].
En 1943, la diffraction au rayon X de l'ADN par William Astbury permet d'émettre la première hypothèse concernant la structure de la molécule : une structure régulière et périodique qu'il décrit comme une pile de pièces de monnaie (like a pile of pennies).
En 1944, Oswald Avery, Colin MacLeod, et Maclyn McCarty démontrent que l'ADN est une molécule associée à une information héréditaire et peut transformer une cellule[9].
Barbara McClintock montre que les gènes peuvent se déplacer et que le génome est beaucoup moins statique que prévu[10]. Elle reçoit le prix Nobel de Médecine en 1983.
En 1950, la structure chimique de l'ADN a été définie par Phoebus Levene (post mortem) et Alexander Robert Todd.
En 1952, Alfred Hershey et Martha Chase découvrent que seul l'ADN d'un virus a besoin de pénétrer dans une cellule pour l'infecter. Leurs travaux renforcent considérablement l'hypothèse que les gènes sont faits d'ADN[11].
En 1953, simultanément aux travaux de recherche de Maurice Wilkins et Rosalind Franklin qui réalisèrent un cliché d'une molécule d'ADN, James Watson et Francis Crick présentent le modèle en double hélice de l'ADN, expliquant ainsi que l'information génétique puisse être portée par cette molécule. Watson, Crick et Wilkins recevront en 1962 le prix Nobel de médecine pour cette découverte.
En 1955, Joe Hin Tjio fait le premier compte exact des chromosomes humains : 46[11]. Arthur Kornberg découvre l'ADN polymérase, une enzyme permettant la réplication de l'ADN.
En 1957, le mécanisme de réplication de l'ADN est mis en évidence.
En 1958, le Pr Raymond Turpin de l'hôpital Trousseau, Marthe Gautier et Jérôme Lejeune réalisent une étude des chromosomes d’un enfant dit « mongolien » et découvre l’existence d’un chromosome en trop sur la 21e paire[12]. Pour la première fois au monde est établi un lien entre un handicap mental et une anomalie chromosomique. Par la suite, Jérôme Lejeune et ses collaborateurs découvrent le mécanisme de bien d’autres maladies chromosomiques, ouvrant ainsi la voie à la cytogénétique et à la génétique moderne.
Dans les années 1960, François Jacob et Jacques Monod élucident le mécanisme de la biosynthèse des protéines. Introduisant la distinction entre « gènes structuraux » et « gènes régulateurs », ils montrent que la régulation de cette synthèse fait appel à des protéines et mettent en évidence l'existence de séquences d'ADN non traduites mais jouant un rôle dans l'expression des gènes. Le principe de code génétique est admis.
En 1961, François Jacob, Jacques Monod et André Lwoff avancent conjointement l'idée de programme génétique.
En 1962, Crick, Watson et Wilkins reçoivent le prix Nobel de médecine pour avoir établi que les triplets de bases étaient des codes. Le comité Nobel évoquera « la plus grande réussite scientifique de notre siècle ».
En 1966, J. L. Hubby et Richard C. Lewontin ouvrent la voie au domaine de la recherche sur l'évolution moléculaire en introduisant les techniques de la biologie moléculaire comme l'électrophorèse sur gel dans la recherche sur la génétique des populations.
- 1968 : prix Nobel décerné pour le déchiffrage du code génétique.
- 1975 : autre prix Nobel pour la découverte du mécanisme de fonctionnement des virus.
La génomique devient dès lors l'objet d'intérêts économiques importants.
Dans le même temps, la sociobiologie et la psychologie évolutionniste d’Edward O. Wilson se fondent sur l'idéologie du déterminisme génétique que génère l'idée — devenue caduque[13] — de programme génétique. De la sorte, c'est-à-dire selon une conception évolutionniste linéaire et réductionniste[14] néodarwinienne et le mythe du Graal[15] de la génétique, ces deux domaines débordent sur la sphère sociale et politique. C'est ainsi que, tout en apportant une conception scientifique « dialectique », Stephen Jay Gould, Richard C. Lewontin et quelques autres membres du groupe de Science for the People ont démarré la polémique encore en cours sur la sociobiologie et la psychologie évolutionniste.
En 1980, la Cour suprême des États-Unis admet pour la première fois au monde le principe de brevetabilité du vivant pour une bactérie génétiquement modifiée (oil-eating bacteria). Cette décision juridique est confirmée en 1987 par l’Office Américain des Brevets, qui reconnaît la brevetabilité du vivant, à l’exception notable de l’être humain.
En 1986, est réalisé le premier essai en champ de plante transgénique (un tabac résistant à un antibiotique).
En 1989, il est décidé de décoder les 3 milliards de paires de bases du génome humain pour identifier les gènes afin de comprendre, dépister et prévenir les maladies génétiques et tenter de les soigner. Une première équipe se lance dans la course : le Human Genome Project, coordonné par le NIH (National Institutes of Health) et composé de 18 pays dont la France avec le Génoscope d'Évry qui sera chargée de séquencer le chromosome 14.
Dans les années 1990, à Évry, des méthodologies utilisant des robots sont mises au point pour gérer toute l'information issue de la génomique.
En 1992, l’Union européenne reconnaît à son tour la brevetabilité du vivant et accorde un brevet pour la création d’une souris transgénique. Elle adopte en 1998 la directive sur la brevetabilité des inventions biotechnologiques : sont désormais brevetables les inventions sur des végétaux et animaux, ainsi que les séquences de gènes. En 1998, l’Europe adopte une Directive fondamentale relative à la protection des inventions biotechnologiques : sont désormais brevetables les inventions sur des végétaux et animaux, ainsi que les séquences de gènes.
Dans le même temps les premiers Mouvement anti-OGM se forment contre le lobby du « complexe génético-industriel »[16] dans le domaine de l'OGM. Les OGM, organismes génétiquement modifiés sont en réalité pour le généticien Richard C. Lewontin et Jean-Pierre Berlan des CCB, clones chimériques brevetés[17]. Cela ouvre de nombreux débats politiques et médiatiques, divers et variés, sur l'OGM conduisant à des réglementations.
En 1992-1996, les premières cartes génétiques du génome humain sont publiées par J. Weissenbach et D. Cohen dans un laboratoire du Généthon.
En 1998, créée par Craig Venter et Perkin Elmer (leader dans le domaine des séquenceurs automatiques), la société privée Celera Genomics commence elle aussi le séquençage du génome humain en utilisant une autre technique que celle utilisée par le NIH.
En 1999, un premier chromosome humain, le 22, est séquencé par une équipe coordonnée par le centre Sanger, au Royaume-Uni.
En , le NIH et Celera Genomics annoncent chacun l'obtention de 99 % de la séquence du génome humain. Les publications suivront en 2001 dans les journaux Nature pour le NIH et Science pour Celera Genomics.
En , des chercheurs japonais de l'université de Tokyo ont introduit 2 nouvelles bases, S et Y, aux 4 déjà existantes (A, T, G, C) sur une bactérie de type Escherichia coli, ils l'ont donc dotée d'un patrimoine génétique n'ayant rien de commun avec celui des autres êtres vivants et lui ont fait produire une protéine encore inconnue dans la nature. Certains n'hésitent pas à parler de nouvelle genèse, puisque d'aucuns y voient une nouvelle grammaire autorisant la création d'êtres vivants qui non seulement étaient inimaginables avant mais qui, surtout, n'auraient jamais pu voir le jour[18].
Le , la fin du séquençage du génome humain est annoncée.
Les années 2010 vont vers la fin du « tout gène » et du réductionnisme de la génétique moléculaire des quarante dernières années avec la découverte de phénomènes épigénétiques liés à l'influence de l'environnement sur le gène[19].
Rudiments concernant la génétique moléculaire
modifierUne réorganisation et une clarification du contenu sont nécessaires.
Qu'est-ce qu'un gène ?
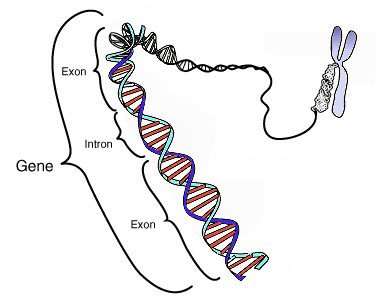
modifierUn gène est une unité d'information génétique, constitué par plusieurs nucléotides (1 nucléotide est constitué par un groupement phosphate, un sucre et une base azotée). Les gènes sont soit codant et leur information génétique est utilisée : 1/pour la biosynthèse des protéines 2/lors de la formation d'un embryon, ou bien sont de l'ADN non codant et dont l'information génétique ne sera pas traduite directement en protéine, mais assurera toute une série d'autres fonctions, comme l'activation et la désactivation de l'expression de certains gènes.
Plus largement, dans une définition prenant en compte les découvertes récentes, notamment sur les microARN, on peut dire qu'un gène est « l'ensemble des séquences d'ADN qui concourent à la production régulée d'un ou plusieurs ARN, ou d'une ou plusieurs protéines »[20].
Quel est le support de l'information génétique ?
modifierL'information génétique est portée par l'acide désoxyribonucléique, ou ADN. L'ADN est une macromolécule formée par l’enchaînement de nombreux nucléotides. Chaque nucléotide est formé d'un groupement phosphate, d'un glucide, le désoxyribose, et d'une base azotée. Il existe quatre bases azotées différentes donc quatre nucléotides différents dans l'ADN : l'adénine, la cytosine, la guanine et la thymine.
La molécule d'ADN est formée de deux chaînes de nucléotides enroulées en double hélice. Les nucléotides sont complémentaires deux à deux : en face d'une cytosine se trouve toujours une guanine ; en face d'une adénine se trouve toujours une thymine.
C'est la séquence, c'est-à-dire l'ordre et le nombre des nucléotides d'un gène, qui porte l'information génétique.
Comment l'information génétique gouverne-t-elle le phénotype ?
modifierL'ADN sert de support pour la synthèse des protéines. L'information génétique portée par l'ADN est « reportée » dans une molécule d'ARNm (acide ribonucléique « messager ») lors de la transcription, puis l'ARNm sert de support pour la synthèse d'une protéine lors de la traduction. Chaque triplet de nucléotide (ou codon) de l'ARNm « code » un acide aminé (cela signifie que chaque triplet « appelle » un acide aminé précis), selon la correspondance établie par le code génétique. Ainsi la séquence en acides aminés de la protéine dépend directement de la séquence en nucléotides de l'ADN. Or les protéines forment le phénotype moléculaire de la cellule ou de l'individu. Le phénotype moléculaire conditionne le phénotype cellulaire et finalement le phénotype de l'organisme.
Comment l'information génétique est-elle transmise lors de la division cellulaire ?
modifierTous les organismes vivants : animaux, végétaux, sont constitués de cellules. Ainsi, un être humain est composé de, selon les auteurs, 50 000 milliards à 100 000 milliards de cellules. Toutes les cellules d'un être vivant proviennent de la même cellule initiale qui s'est divisée un très grand nombre de fois, au cours de l'embryogenèse puis du développement fœtal.
Au cours d'un cycle cellulaire (succession des étapes de la vie de la cellule), la cellule réplique son ADN, c'est-à-dire que toute l'information génétique est dupliquée à l'identique : elle se retrouve avec deux « copies » complètes de son information génétique, ses chromosomes sont constitués de deux chromatides identiques. Lors de la division cellulaire, ou mitose, les deux chromatides de chaque chromosome se séparent pour former deux lots identiques de chromosomes (à une seule chromatide). Chaque cellule fille reçoit un de ces lots. Ainsi, au terme d'une mitose, les deux cellules filles issues de la cellule mère possèdent exactement le même patrimoine génétique : elles sont des copies conformes l'une de l'autre[20].
Génétique et société
modifierIdéologie et philosophie
modifierLes débuts de la génétique ont été influencés par deux idéologies dominantes et hégémoniques opposées et exacerbées dans les années 1930 :
- dans les pays occidentaux, la plupart des généticiens ont adhéré à l'eugénisme. L'eugénisme est une invention du néoconservateur Francis Galton à la fin du XIXe siècle sur la base d'une sélection artificielle des individus. Cette idéologie fut amalgamée à l'idée spencériste. Ce dernier se fonde sur la sélection naturelle en société par analogie à la sélection naturelle existante dans la nature. Avec le malthusianisme, elles sont, selon le point de vue de la bourgeoisie, en adéquation avec la société capitaliste du XIXe siècle. Ces pratiques sont, par la suite, soutenues par la fondation Rockefeller[21] dès 1912 et l'UNESCO par son premier président Julian Huxley en 1945 ;
- dans le bloc soviétique jusque dans les années 1960, la génétique a été interdite par l'intervention du transformiste Trofim Lyssenko, qui s'oppose à la génétique « mendelo-morgannienne ». Staline va placer sa confiance dans Lyssenko dans la période de troubles des années 1930, parce que les principes transformistes de Lyssenko sont en adéquation avec le lamarkisme social de la société soviétique. Cette confiance a conduit Lyssenko à faire faire condamner Nikolaï Vavilov et ses collaborateurs darwino-mendéliens qui sont ainsi envoyés au goulag en 1939. Dans les années 1950, Staline y voit un non-sens[22] dans la terminologie de « science prolétarienne » inventée par Lyssenko qui veut l'opposer à une science bourgeoise. Mais, c'est surtout pendant l'ère post-stalinienne, la période Nikita Khrouchtchev, que Lyssenko aura le plus de soutiens. Ce soutien vient non de la part de scientifiques, comme le biologiste Jacques Monod proche du PCF ou encore le généticien marxiste et communiste J.B.S. Haldane, mais de militants non scientifiques comme le philosophe, communiste Jean-Paul Sartre[23]. Lyssenko est limogé en 1965 après la chute de son protecteur Nikita Khrouchtchev[23].
Ces oppositions idéologiques s'ouvrent sur la question philosophique de l'inné ou de l'acquis de l'acquisition des connaissances des individus et du développement de la culture humaine bien que Charles Darwin y ait déjà répondu dès 1871 dans le passé-inaperçu La Filiation de l'homme[24]. Il n'y a pas d'opposition entre l'inné (génétique) et l'acquis (l'environnement). Cependant, pour les néo-darwinistes ou sociobiologistes, la question du comportement de l'homme se pose encore. Mais, pour le généticien Richard C. Lewontin, « Il n’y a pas de « part » respective des gènes et de l’environnement, pas plus qu’il n’y a de « part » de la longueur et de la largeur dans la surface d’un rectangle, pour reprendre une métaphore classique. L’exposition à l’environnement commence d’ailleurs dans le ventre maternel, et inclut des événements biologiques comme la qualité de l’alimentation ou l’exposition aux virus. Génétique et milieu ne sont pas en compétition, mais en constante interaction : on dit qu’ils sont covariants. Le comportement d’un individu serait donc à la fois 100 % génétique et 100 % environnemental ».
La génétisation est un concept introduit par l’épidémiologiste et militante radicale Abby Lippman (en) travaillant à McGill dans le département d’épidémiologie en 1991. Ce concept est défini comme la réduction d’un individu à son génome. Plus précisément, tout problème d’un individu pourrait être réduit à un problème de génétique, qu’un gène pourrait expliquer un comportement, un trait, etc. Abby Lippman était constructionniste : les conclusions tirées de la science sont le produit de l’esprit humain et des interactions entre êtres humains.
En développant sa théorie sur la génétisation, Lippman avait en tête que les conclusions étaient constructionnistes. Les trois concepts pouvant amener un problème donné à une construction sont : le réductionnisme, le déterminisme et l’essentialisme en génétique [incompréhensible].
- Le réductionnisme réduit l’explication de traits biologiques à l'expression d'un gène spécifique.
- Le déterminisme est l’exclusion de tous autres facteurs (comme l’environnement).
- L’essentialisme est l’idée que les gènes, composants immuables de l’être humain pourront le définir entièrement.
L’idée derrière ces trois concepts est que l’on peut définir l'humain à travers une liste de gènes qui le détermineront définitivement et immuablement.
Plusieurs auteurs ont ainsi soulevé différentes inquiétudes concernant ces thèmes. Alpers et Beckwith écrivaient en 1993 que le déterminisme génétique pouvait justifier les inégalités et la discrimination. Nelkin (en) et Lindee (en) professeurs à l’université de New York et de Pennsylvanie respectivement en 1995 disaient que la génétique dans la culture populaire était synonyme d’essentialisme génétique ce qui favorise aussi les discriminations.
Dans les années 2000, la génétisation évolue vers une théorie beaucoup moins radicale que celle énoncée par Lippman. En 2003, le projet Génome Humain qui avait pour but de séquencer le génome humain se termine mettant en avant plus de 20 000 gènes. Cette découverte signe la fin de l’idée que les gènes étaient une liste de caractéristiques physiologiques et fit reculer l'idée déterminisme en génétique (Fox Keller, 2000). Plusieurs interactions entre différents gènes ont aussi été découvertes et la complexité de la génétique explosa mettant fin au réductionnisme.
Le professeur Adam Hedgecoe de l’université de Londres avance dans une étude faite en 2001 que les chercheurs en schizophrénie s’intéressaient déjà plus à la combinaison de plusieurs facteurs pour l’origine de cette maladie qu’un simple gène qui pourrait directement décrire la schizophrénie même si le rôle des gènes gardait une place importante pour eux. La théorie devait donc évoluer pour donner aux gènes une place centrale, mais pas absolue[26].
Les avancées ont continué dans les années 2010 (arrivée de l’épigénétique, complexité entre les interactions, etc) qui poussaient le réductionnisme à disparaître. À la suite de toutes ces évolutions et de la perte d’une vue claire de la génétisation, Arribas-Ayllon professeur de science sociale à l’université de Cardiff en 2016 dans l’article After Geneticization écrit que le terme de génétisation avait effectivement un sens lors de sa création, mais qu’avec les avancements des technologies et des connaissances en génétique, ce terme a perdu de son utilité.
Économie, éthique et loi
modifierCas du génome Humain
modifierLors de l'ouverture de la quête du Graal que fut le Projet génome humain pour les généticiens[27], le laboratoire Celera Genomics dirigée par Craig Venter conduit une course contre le consortium international public pour obtenir le premier des séquences génétiques dans le but de les breveter et de les vendre aux sociétés pharmaceutiques.
Une étude[28] de 2005 révèle que 20 % des gènes humains font l’objet d’un brevet : 63 % de ces brevets appartiennent à des firmes privées, 28 % à des universités.
Le brevetage d'une partie des gènes constitue donc un frein à la découverte de leur fonction.
Par exemple[29], la compagnie Myriad dépose un brevet sur l’utilisation des gènes BRCA1 et BRCA2 séquencé en 1994/1995 comme indicateurs de risques pour le cancer du sein et de l’ovaire, maladie dont les gènes ont été associés. Le test coûte d'abord 1 600 $ US et son prix est passé à 3 200 $ en 2009. Ce brevet représente l’essentiel des revenus annuels de la compagnie. Ainsi, le brevet qui donne le droit de propriété exclusif sur la séquence, empêche complètement d’autres compagnies de développer des tests alternatifs utilisant ces mêmes gènes.
Cependant en 2010, ce brevet est annulé car le caractère inventif du test est contesté par le bureau européen des brevets. la Convention européenne sur les brevets, faisaient valoir que l'invention revendiquée ne remplissait pas les conditions de nouveauté, d'activité inventive et d'application industrielle et qu'elle était insuffisamment décrite pour permettre à une personne du métier de l'exécuter[30]. En effet, le séquençage ne constitue pas une invention, mais une découverte. La méthode se limite à comparer une séquence de l’échantillon à une séquence de référence, et n’est pas brevetable en soi (Directive sur la brevetabilité des inventions biotechnologiques). Ainsi, pour une plus grande liberté de la recherche alternative la cour a donné son verdict : « les produits naturels et propriétés naturelles des objets vivants sont légalement distincts d’objets manufacturés en usant d’une ingéniosité substantielle. »
Ce cas met en évidence l'importance des critères techniques de brevetabilité pour protéger l'intérêt public. Ces critères servent à empêcher l'octroi de brevets pour des inventions qui n'apportent aucune véritable avancée dans la technologie et qui sont utilisées pour restreindre l'accès à des objets du domaine public. Cependant, trouver un équilibre dans cette démarche demeure complexe. Tant que la recherche génétique visant à développer des solutions en matière de traitement et de diagnostic est principalement menée par le secteur privé, des entreprises biopharmaceutiques comme "Myriad" continueront à revendiquer un certain monopole sur ces technologies pour rentabiliser leurs investissements. Il est toutefois préoccupant que certains brevets génétiques puissent accorder à leurs détenteurs une compensation excessive, notamment lorsque les revendications sont formulées de manière trop générale, ce qui limite toute innovation ultérieure[31].
Cas des OGM
modifierRéférences
modifier- « étymologie / génétique », sur CNRTL,
- Dans l'article de Nature Molecular Structure of Nucleic Acids: A Structure for Deoxyribose Nucleic Acid.
- (en) https://www.nytimes.com/2008/08/13/science/13visual.html?_r=1&em&oref=slogin : carte génétique de l'Europe sur le site du New York Times
- Johann Gregor Mendel, « Recherches sur des hybrides végétaux » communications faites le 8 février et le 8 mars 1865, publiées dans Verhandlungen des naturforschenden Vereines in Brünn, p. 3-47 du tome IV, 1865. Traduction de l'allemand par Albert Chappelier parue en 1907 dans le Bulletin scientifique de la France et de la Belgique, tome 41, 1907, p. 371-419.
- (en) Sutton, Walter, « The chromosomes in heredity », Biological Bulletin 4 (1903): 231-251.
- (en) Garrod, A. E. « The incidence of alkaptonuria: a study in chemical individuality » Lancet II: 1616-1620, 1902.
- (en) Morgan, Thomas Hunt et al. The mechanism of Mendelian heredity, (New York: Henry Holt and Co., 1915)
- (en) Beadle, G. and Tatum, E. « Genetic control of biochemical reactions in Neurospora » Proc Natl Acad Sci. 27: 499-506, 1941.
- (en) Avery, Oswald T., MacLeod, Colin M., and McCarty, Maclyn « Studies on the Chemical Nature of the Substance Inducing Transformation of Pneumococcal Types: Induction of Transformation by a Deoxyribonucleic Acid Faction Isolated from Pneumococcus Type III » Journal of Experimental Medicine février 1979;149:297-326. (Ré-impression de la publication originale de 1944).
- (en) Barbara McClintock « The origin and behavior of mutable loci in maize », Proceedings of the National Academy of Sciences, vol. 36, p. 344-355, 1950.
- (en) Tjio and Levan, « The chromosome number in man », Hereditas, vol. 42, p. 1, 1956.
- J. Lejeune, M. Gautier et R. Turpin. Les chromosomes humains en culture de tissus. C. R. Acad. Sciences, 26 janvier 1959.
- Jean-Jacques Kupiec, Olivier Gandrillon, Michel Morange, Marc Silberstein (sous la direction de) Le hasard au cœur de la cellule. Probabilités, déterminisme, génétique, Paris, Syllepse, 2009. Nouvelle édition revue et augmentée aux Éditions Matériologiques, 2011.
- Stephen Jay Gould dans Le renard et le hérisson - Comment combler le fossé entre la science et les humanités montre que le terme de Consilience employé par Edward O. Wilson dans l'Unité du savoir est employé comme synonyme de réductionnisme, c'est-à-dire une vision linéaire du mouvement des choses et non de celui de consilience défini par William Whewell et S.J. Gould.
- Richard Lewontin, Le rêve du génome humaine, Politique et écologie no 5, 1993
- Jean-Pierre Berlan et Richard C. Lewontin, La menace du complexe génético-industriel, le Monde diplomatique, décembre 1998.
- Jean-Pierre Berlan et Richard C. Lewontin, OGM ou CCB ? - Les organismes génétiquement modifiés sont, en réalité, des clones chimériques brevetés, 2004.
- les lois du code genetique violees : et maintenant ?.
- Dossier Pour la science, « L'hérédité sans gènes - comment l'environnement rebat les cartes », hors série 81, octobre-décembre 2013.
- « La Génétique pour les Nuls », sur Pour les nuls (consulté le ).
- Alain Drouard, « Un cas d’eugénisme « démocratique » », La Recherche, .
- Yann Kindo, « L’affaire Lyssenko, ou la pseudo-science au pouvoir » SPS no 286, juillet-septembre 2009 : « En 1950, Staline lui-même condamne la distinction science bourgeoise/science prolétarienne (dans un opuscule sur la linguistique), ce qui coupe court à la campagne du PCF. ».
- Yann Kindo, « L’affaire Lyssenko, ou la pseudo-science au pouvoir » SPS no 286, juillet-septembre 2009.
- Charles Darwin, La Filiation de l’Homme et la sélection liée au sexe, trad. sous la direction de P. Tort, coord. par M. Prum. Précédé de Patrick Tort, « L’anthropologie inattendue de Charles Darwin ». Paris, Champion Classiques, 2013.
- (en) Michael Arribas-Ayllon, « After geneticization », Social Science & Medicine, vol. 159, , p. 132–139 (ISSN 0277-9536, DOI 10.1016/j.socscimed.2016.05.011, lire en ligne, consulté le )
- (en) Adam Hedgecoe, « Schizophrenia and the Narrative of Enlightened Geneticization », Social Studies of Science, vol. 31, no 6, , p. 875–911 (ISSN 0306-3127 et 1460-3659, DOI 10.1177/030631201031006004, lire en ligne, consulté le )
- Richard C. Lewontin, Le rêve du génome humain, Politique et écologie, 1993.
- (en) Jensen & Murray (2005) « Intellectual Property Landscape of the Human Genome » Science 310: 239 - 240.
- (en) Kesselheim and Mello « Gene patenting--is the pendulum swinging back? » N Engl J Med. (2010) vol. 362 (20) p. 1855-8.
- « Bioéthique et droit des brevets: L’affaire Myriad », sur www.wipo.int (consulté le )
- « Bioéthique et droit des brevets: L’affaire Myriad », sur www.wipo.int (consulté le )
Bibliographie
modifier- Bertrand Jordan, L'humanité au pluriel : la génétique et la question des races, Paris, Seuil, coll. « Science ouverte », , 227 p. (ISBN 978-2-02-096658-0)
Voir aussi
modifierArticles connexes
modifierLiens externes
modifier
- Ressources relatives à la santé :
- Ressource relative à la recherche :
- Notices dans des dictionnaires ou encyclopédies généralistes :
- Didaquest.org : « Génétique Chronologie »
- Glossaire de la biotechnologie de la FAO
- « Il était une fois... l'ADN » : un site éducatif sur les bases de la génétique classique et moléculaire
- Génétique dans l'élevage chiens de race : consanguinité, retrempe, gènes codant la couleur de la robe, la longueur de la queue.
- (en) DNA from the beginning 75 animations pour expliquer la génétique pas à pas.
- (en) Base de données libre sur la génétique humaine
- (en) Base de données des génomes séquencés, ainsi que les projets en cours
- (en) Découverte en 2007 des zones d'instabilité du génome chez la souris (voir aussi équilibres ponctués)
- Voir aussi pour les végétaux : http://www.interactif-agriculture.org/pages/l-evolution-des-techniques/la-genetique.php