Homme de Néandertal
Homo neanderthalensis
« Néandertal » et « Neandertal » redirigent ici. Pour la vallée de Neander (Neandertal en allemand), voir Vallée de Neander.
Vous lisez un « article de qualité » labellisé en 2006.

l'homme de La Chapelle-aux-Saints 1

- en Europe
- au Moyen-Orient
- en Asie centrale
- dans l'Altaï
L'Homme de Néandertal (Homo neanderthalensis), ou Néandertalien, est une espèce éteinte du genre Homo, qui a vécu en Europe, au Moyen-Orient et en Asie centrale, jusqu'à environ 30 000 ans avant le présent. Selon une étude génétique publiée en 2016, il partage avec l'Homme de Denisova un ancêtre commun remontant à environ 450 000 ans. Cet ancêtre partage lui-même avec Homo sapiens un ancêtre commun remontant à environ 660 000 ans[1],[2],[3]. Les plus anciens Néandertaliens fossiles reconnus comme tels sont ceux de la sima de los Huesos (sierra d'Atapuerca, Espagne), datés de 430 000 ans[4],[1].
Depuis sa découverte en 1856, son statut a varié : un temps considéré comme une sous-espèce d'Homo sapiens et nommé en conséquence Homo sapiens neanderthalensis, il est aujourd'hui considéré comme une espèce à part entière.
Particulièrement bien adapté pour vivre dans un climat froid[5], l'Homme de Néandertal était physiquement plus robuste, plus lourd et plus trapu qu’Homo sapiens. La forme oblongue de son crâne se distingue nettement de celle de l'Homme moderne, plus globulaire. Néandertal avait un cerveau un peu plus volumineux en moyenne, mais avec un coefficient d'encéphalisation légèrement moindre.
Premier homme fossile identifié, contemporain d'Homo sapiens, l'Homme de Néandertal a longtemps pâti de jugements négatifs par rapport à l'Homme moderne. Les progrès de l'archéologie préhistorique depuis les années 1960 ont en fait révélé une espèce humaine d'un certain développement culturel. Il maitrisait différentes techniques avancées comme le collage au brai de bouleau, et certains vestiges fossiles datés de moins de 70 000 ans sont considérés comme des sépultures témoignant de rites funéraires.
De nombreux points restent encore à élucider, comme son ascendance précise ainsi que la date et les conditions de son extinction après plus de 400 000 ans d'existence. Les derniers vestiges fossiles ou archéologiques néandertaliens connus sont datés de moins de 30 000 ans, dans le sud de la péninsule Ibérique, en Crimée, et dans le Caucase. Toutefois, ces datations restent débattues au sein de la communauté scientifique.
Le séquençage de l'ADN nucléaire néandertalien réalisé depuis 2006 et publié à partir de 2010 a montré un « flux de gènes » ancien entre les hommes de Néandertal et les hommes modernes d'Eurasie. Les humains actuels non africains possèdent entre 1,8 et 2,6 % de gènes néandertaliens, acquis par hybridation il y a environ 50 000 ans, peu après leur sortie d'Afrique, et environ 20 % du génome de Néandertal survit dans l'ensemble de la population actuelle à différents endroits de notre génome[6]. Certains gènes néandertaliens auraient été fixés chez l'Homme moderne en raison de leur caractère adaptatif[7].
Historique de la découverte
[modifier | modifier le code]Les Néandertaliens avant Neandertal
[modifier | modifier le code]
Deux fossiles de Néandertaliens ont été découverts avant celui auquel on a donné ce nom. En 1829, un crâne d'enfant, Engis 2, fut mis au jour par Philippe-Charles Schmerling à Flémalle (Belgique) ; nommé comme la commune voisine parce que Schmerling a pensé qu'il se trouvait sur la commune d'Engis. En 1848, un crâne d'adulte fut trouvé à Gibraltar, dans le site de la carrière de Forbes. Si le premier appartenait à un jeune individu sur lequel les traits caractéristiques des Néandertaliens sont moins évidents, le deuxième aurait pu conduire à reconnaître l'existence d'une espèce humaine fossile. Sans doute était-il trop tôt, comme le prouvent d'ailleurs les difficultés pour faire admettre que les os recueillis en 1856 à Neandertal, en Allemagne, correspondaient bien à un homme fossile[8].
1856 : Neandertal ou la « vallée de l'homme nouveau »
[modifier | modifier le code]
Le mot « Néandertalien » est tiré de Neandertal, nom d'une petite vallée située sur le territoire des villes d'Erkrath et de Mettmann, entre Düsseldorf et Wuppertal (Allemagne). Au mois d', dans le cadre de l'exploitation d'une carrière, des ouvriers vidèrent une petite cavité de cette vallée, la petite grotte de Feldhofer (en). Ils y découvrirent des ossements et un fragment de crâne qu'ils remirent à Johann Carl Fuhlrott, un enseignant d'Elberfeld passionné d'histoire naturelle.
Par un heureux hasard, le toponyme Neandertal signifie « vallée de l'homme nouveau ». En effet le nom de Neander a été donné à cette vallée (en allemand tal, anciennement thal) en l’honneur de Joachim Neumann (1650-1680), appelé aussi Joachim Neander, car, suivant un usage familial datant de son grand-père et très courant à l'époque, il avait traduit en grec ancien son patronyme allemand, qui signifie littéralement « homme nouveau ». Ce pasteur et compositeur, auteur de cantiques religieux encore populaires dans le protestantisme allemand, aimait chercher son inspiration dans cette vallée, jadis idyllique.
Comme, à l'époque, le nom de la vallée s'écrivait encore Neanderthal, l'homme qui y fut découvert reçut le nom latin d’Homo neanderthalensis. Ultérieurement, une réforme orthographique de l'allemand a supprimé les h superflus, mais, la nomenclature évitant de revenir sur les formes latinisées, on a continué à écrire Homo neanderthalensis. La graphie française la plus courante, proposée par Henri Vallois en 1952, est Homme de Néandertal, même si l'on trouve parfois Homme de Neandertal, Homme de Néanderthal ou Homme de Neanderthal. En anglais, la forme ancienne Neanderthal est encore très répandue, ce qui peut induire pour la séquence thal une prononciation incorrecte du nom allemand originel[9].
Une découverte controversée
[modifier | modifier le code]
Johann Carl Fuhlrott comprend rapidement l'intérêt de la découverte et se rend sur place pour tenter en vain de découvrir d'autres ossements ou des vestiges qui leur seraient associés. Il se rend compte qu'il s'agit d'ossements anciens mais surtout incroyablement primitifs, correspondant à un homme nouveau, d'une « conformation naturelle jusqu'ici inconnue »[10],[11].
L'Homme de Néandertal est effectivement le premier homme fossile distinct d'Homo sapiens, et il est découvert avant l'Homme de Cro-Magnon (1868). L'idée même qu'une espèce d'homme distincte de la nôtre ait existé par le passé (et ait disparu) fut d'ailleurs particulièrement difficile à admettre. On se souviendra par exemple que Charles Darwin ne publiera L'Origine des espèces par la sélection naturelle qu'en 1859 et qu'il n'élargira explicitement sa théorie à l'homme qu'en 1871 dans La Filiation de l'homme et la sélection liée au sexe.
Malgré des différences importantes avec les os d'hommes modernes, Fuhlrott reconnait dans ses trouvailles des os humains et les soumet à Hermann Schaaffhausen pour un examen complémentaire. Ce dernier présente ses premières conclusions en 1857[12]. Il estime que les ossements datent d'une période antérieure aux Celtes et aux Germains, et sont ceux d'un individu appartenant à l'une des races sauvages du nord-ouest de l'Europe dont parlent les auteurs latins. Tous les chercheurs n'acceptent pas cette interprétation : pour certains, les os ont appartenu à un genre différent du nôtre, sans doute plus proche du singe ; pour d'autres, ils renvoient à un individu pathologique ou frappé de crétinisme ; d'autres encore évoquent un cosaque mongol aux jambes arquées ayant déserté les armées russes en 1814.
Reconnaissance de l'ancienneté de l'Homme et de son évolution
[modifier | modifier le code]
Peu à peu les découvertes se multiplient. Viennent d'abord celles de fossiles d'Homo sapiens associés à des vestiges lithiques et à des animaux disparus (dont l'Homme de Cro-Magnon en 1868) ; puis d'autres Homo neanderthalensis, encore en place dans les sédiments (mandibule de la grotte des Fées à Arcy-sur-Cure en 1859[13]), complets et présentant les mêmes spécificités anatomiques, mais souvent hors contexte archéologique (pas d'ossements d'animaux ou d'outils associés), ce qui rend difficile leur datation et leur interprétation. Parmi les plus spectaculaires, il faut citer les deux squelettes de la grotte de Spy (région wallonne de Belgique) en 1886 puis la sépulture de l'Homme de La Chapelle-aux-Saints 1 (Corrèze) en 1908. Elles contribuent à faire définitivement accepter l'existence d'une nouvelle espèce d'humains par la communauté scientifique.
Le nom scientifique Homo neanderthalensis est proposé en 1864 par William King, professeur au Queen's College de Galway en Irlande et ancien élève de Charles Lyell[14]. En 1866, Ernst Haeckel propose le nom surprenant d’Homo stupidus, qui n'est pas retenu en vertu des règles de nomenclature donnant priorité à l'appellation antérieure. Les partisans du rattachement à une sous-espèce parleraient sinon d’Homo sapiens stupidus !
Les premières études (et les reconstitutions qui en découlaient) donnèrent de l'Homme de Néandertal une image déformée, accentuant les traits primitifs, voire simiesques. Ce fut le cas de l'étude de l'Homme de La Chapelle-aux-Saints 1 publiée par Marcellin Boule en 1911 : même s'il s'agissait d'une étude très complète, qui fit référence pendant de nombreuses années, elle présentait un Homme de Néandertal voûté, la colonne vertébrale courbée (comme chez les gorilles) et les membres inférieurs semi-fléchis[15]. Il fallut presque un siècle à la communauté scientifique pour corriger cette perception influencée par des a priori peu scientifiques.
Au début du XXe siècle, certains furent scandalisés par le fait que ces découvertes se détachaient d'une lecture littérale de la Bible[16]. Ils reprochaient au prêtre catholique Jean Bouyssonie, qui découvrit le squelette d'un Néandertalien à La Chapelle-aux-Saints 1, de soutenir la théorie de l'évolution. Le , la légende d'une caricature de La Lanterne indique : « Les savants prétendent que c’est le crâne du plus ancien homme du Monde. C’est une malveillante insinuation destinée à faire croire que les hommes du Monde descendent du singe ». La caricature montre Jean Bouyssonie en soutane, présentant sa découverte à un savant[17],[18],[19].
Phylogénie et génétique
[modifier | modifier le code]Le statut phylogénétique de l'homme de Néandertal provoque encore quelques débats. Il s’agit d’un simple problème de définition de l'espèce. Deux sous-espèces peuvent se croiser et avoir une descendance fertile, mais c'est beaucoup plus variable pour deux espèces différentes (par exemple le cheval et l'âne, le tigre et le lion) : certaines le peuvent et d'autres pas. L'infertilité de la descendance prouve l'existence de deux espèces distinctes, mais l'inverse n'est pas vrai (s'il y a deux espèces, la descendance n'est pas nécessairement infertile). On peut rappeler ici qu'il existe une vingtaine de définitions de l'espèce, et que l'isolement reproductif n'est que l'une d'entre elles.
Lors de sa dénomination en 1864, l’hypothèse d’une espèce distincte avait été privilégiée. Dans les années 1960, certains spécialistes ont considéré les Néandertaliens comme une sous-espèce d'Homo sapiens, comme le généticien Theodosius Dobjansky ou encore le biologiste Ernst Mayr, qui déclarait que « jamais plus d'une seule espèce d'homme n'a existé au même moment ». Aujourd’hui, l’idée d’espèces distinctes est à nouveau dominante, notamment grâce aux apports de la génétique.
Les multiples études paléoanthropologiques effectuées sur les ossements ne permettaient pas de se prononcer clairement sur la classification de l'homme de Néandertal. Des analyses comparées d'ADN nucléaire, extrait d'ossements de Néandertaliens et d'Homo sapiens anciens et modernes, publiées depuis 2010, ont largement contribué à forger un nouveau consensus[20].
En 2006, le projet génome de Néandertal, un programme de séquençage de l'ADN nucléaire de l'homme de Néandertal, a été lancé par l'Institut Max-Planck d'anthropologie évolutionniste, à Leipzig en Allemagne, en collaboration avec la société 454 Life Sciences fabriquant des séquenceurs de gènes à haut débit. L'objectif était de connaitre l'étendue du lien de parenté avec l'homme moderne et d'évaluer l'interfécondité de l'homme de Néandertal et de l'homme moderne. Ce projet a permis d'achever le séquençage du génome néandertalien dès 2009 et de publier les premières études en 2010.
Une étude de 2016 exploitant le séquençage de l'ADN nucléaire de spécimens de la Sima de los Huesos (Espagne), datés de 430 000 ans, comparé avec le génome de spécimens d'Homo sapiens, d'hommes de Néandertal et d'hommes de Denisova, a attribué les fossiles de la Sima de los Huesos à l'espèce Homo neanderthalensis, et indiqué que la séparation entre la lignée des hommes modernes et celle des humains archaïques, Dénisoviens et Néandertaliens, a eu lieu entre 550 000 et 760 000 ans avant le présent. La séparation entre Dénisoviens et Néandertaliens est quant à elle estimée entre 381 000 et 473 000 ans. Les fossiles de la Sima de los Huesos étant datés de 430 000 ans, on peut estimer cette dernière séparation à un âge d'environ 450 000 ans[1]. Pour la première fois, les liens entre différents représentants du genre Homo ont pu ainsi être établis.

Génome de Néandertal
[modifier | modifier le code]En 2018, le génome de cinq nouveaux Néandertaliens ayant vécu il y a 39 000 à 47 000 ans a pu être étudié (le nombre de Néandertaliens dont on a séquencé le génome a ainsi doublé)[21]. Ces cinq personnes avaient un génome très similaire à ceux des Néandertaliens tardifs déjà connus, conformément à ce que prédisait leur situation géographique[21]. Et bien que quatre de ces Néandertaliens aient été contemporains des premiers humains modernes en Europe, aucune trace d'apport génétique des hommes modernes n'a pu alors être décelée[21]. Des études ultérieures démontrent cependant qu'il s'est bien produit une introgression de gènes sapiens chez Néandertal, sans doute à au moins deux reprises (voir Gènes transmis par Sapiens à Néandertal).
Les apports de gènes néandertaliens chez les humains modernes outrafricains proviennent de contacts, peu après leur sortie d'Afrique, avec des Néandertaliens représentatifs des fossiles européens tardifs (leur dernier ancêtre commun datant d'environ 80 000 ans). Mais ces lignées sont sensiblement plus éloignées de celle d'un Néandertal de l'Altaï, qui a divergé il y a environ 140 000 ans et des Dénisoviens qui se sont séparés il y a au moins 400 000 ans[22].
Une étude de Johannes Krause, co-directeur du département de paléogénétique humaine de l'Institut Max-Planck d'anthropologie évolutionniste, datant de 2022, précise que Néandertal avait des prédispositions génétiques au langage. L'équipe des scientifiques a extrait et analysé des échantillons d’ADN de néandertaliens provenant de la grotte d'El Sidrón en Espagne[23]. Les généticiens ont isolé chez des Néandertal la variante humaine du gène FOXP2, responsable du développement des zones cérébrales liées au langage.
Histoire et répartition des Néandertaliens
[modifier | modifier le code]
L'apparition de l'homme de Néandertal est une question complexe qui dépend notamment de la définition qu'on choisit d'adopter. Avant les Néandertaliens classiques, les fossiles deviennent beaucoup plus rares et les datations moins précises, encourageant de nombreuses théories concurrentes. Cependant l'analyse de l'ADN nucléaire de la Sima de los Huesos en 2016 a permis de consolider une première affirmation sur les origines de Néandertal : sa lignée se sépare d'avec Homo sapiens il y a environ 660 000 ans et ses premiers fossiles seraient justement ceux de la Sima datés de 430 000 ans. Ceux-ci présentent de nombreux caractères intermédiaires mais leur dentition est déjà clairement néandertalienne, suggérant une spécialisation initiale de l'appareil masticatoire[24],[25],[4],[1]. Les débats se poursuivent sur l'attribution des autres fossiles de cette période du Pléistocène moyen : Aroeira 3 montre des caractères néandertaliens[26], mais d'autres fossiles sont d'une attribution moins claire. Jean-Jacques Hublin avance un modèle d'accrétion, où des populations successives auraient accumulé progressivement des caractères dérivés pour donner naissance au jeu de caractères commun aux Néandertaliens classiques[27].
L'Homme de Néandertal est une espèce dont l'apparition et l'évolution sont connues principalement en Europe. Les Néandertaliens découverts au Moyen-Orient, sur les territoires actuels de l'Irak, de la Syrie (grotte de Dederiyeh[28]), du Liban (Ksar Akil) et d'Israël, ainsi qu’en Asie centrale (Enfant de Teshik-Tash, en Ouzbékistan) et en Sibérie sont à ce jour moins nombreux et plus tardifs, ce qui pourrait être dû à des fouilles moins avancées. En 2007, la répartition géographique des Néandertaliens a été repoussée de 2 000 km vers l'est par rapport au site de Teshik-Tach, le plus oriental connu jusqu'alors. Des fragments osseux de la grotte Okladnikov, dans l'Altaï, jusqu'alors mal référencés, sont désormais attribués à des Néandertaliens après une analyse génétique de leur ADN mitochondrial par l'Institut Max-Planck d'anthropologie évolutionniste de Leipzig. D'après les chercheurs, l'ADN mitochondrial des Néandertaliens de l'Altaï est d'ailleurs plus proche de celui des Néandertaliens de la grotte Scladina, en Belgique, que de celui de l'Ouzbékistan, suggérant plusieurs vagues de migrations et de peuplements de la région. L'équipe du généticien Svante Pääbo a suggéré que la présence de Néandertaliens dans l'Altaï rendait envisageable une extension plus orientale, en Mongolie, voire jusqu'en Chine[29],[30].
Les estimations du nombre total de néandertaliens par les paléodémographes présentent une très grande variabilité. L'effectif maximal est évalué à 70 000 néandertaliens. La densité des populations était très faible, de l'ordre de 100 individus pour 10 000 km2, se répartissant en 2 à 3 000 clans de 20 à 35 personnes[31].
L'homme de Néandertal était nomade mais pas errant, ce qui signifie qu'il se déplaçait entre des points connus et qui comportaient un intérêt (zone de chasse, carrière de silex)[32].[source insuffisante]
Premiers peuplements européens
[modifier | modifier le code]Il y a plus d'un million d'années, quelques groupes humains sont arrivés en Europe et ont laissé des traces sous forme de fossiles et de galets taillés de type oldowayen. Les plus anciens fossiles humains européens datent de 1,2 à 1,5 million d'années et ont été mis au jour en Espagne (Sima del Elefante et Homme d'Orce) et en Bulgarie (Kozarnika). Ils sont cependant trop fragmentaires pour avoir pu être attribués à une espèce précise.
Homo heidelbergensis
[modifier | modifier le code]À partir de −700 000 ans, le peuplement de l'Europe se renouvelle avec probablement l'arrivée d'Homo heidelbergensis, porteur de l'industrie acheuléenne. À cette époque, plusieurs espèces appartenant au genre Homo coexistaient en Europe et en Asie. L'une d'elles a évolué pour donner les Néandertaliens.
Les fossiles européens de cette période sont généralement attribués à Homo heidelbergensis : c’est le cas de l’Homme de Tautavel (−450 000 ans), trouvé dans le massif des Corbières en France, de la mandibule de Mauer (−610 000 ans), trouvée près de Heidelberg en Allemagne, ou du crâne de l'Homme de Petrálona trouvé dans la grotte de Petrálona en Chalcidique (Grèce) (environ −700 000 ans).
Prénéandertaliens
[modifier | modifier le code]L’évolution qui conduit au développement d’Homo neanderthalensis, parfois appelée « néandertalisation », est un processus lent et progressif. Elle peut être suivie depuis différents fossiles, qualifiés de « pré-Néandertaliens », jusqu’aux Néandertaliens récents[33].
Un crâne daté de 400 000 ans, Aroeira 3, découvert en 2014 dans la grotte d'Aroeira au centre du Portugal, présente un mélange de caractéristiques jamais observé jusqu'alors chez les humains fossiles ; cet individu présente des traits le rendant proche des Néandertaliens mais aussi certains traits plus primitifs évoquant d'autres espèces humaines éteintes en Europe. Il pourrait contribuer à mieux comprendre les lignées d'Europe ayant évolué vers les Néandertaliens[26].
Les fossiles de Swanscombe (Kent, Angleterre, 400 000 ans), le crâne de l'Homme de Steinheim (Allemagne, 300 000 ans) et de la Sima de los Huesos à Atapuerca (Espagne, 430 000 ans) sont plus clairement attribués aux Prénéandertaliens.
Les Néandertaliens anciens
[modifier | modifier le code]Les restes de trois os longs (humérus, radius, cubitus) du bras gauche d’un individu adulte, de sexe indéterminé, datés d'environ 210 000 ans, ont été découverts en septembre 2010 sur une fouille de l'Inrap à Tourville-la-Rivière (Normandie, France). Leur étude a été publiée en octobre 2014[34],[35].
Les plus anciens fossiles de morphologie néandertalienne presque complète ont des âges compris entre - 250 000 et - 110 000 ans. Parmi eux, on peut citer le crâne de Biache-Saint-Vaast (Pas-de-Calais), vieux de 180 000 ans[36], les restes de La Chaise à Vouthon (Charente), la mandibule de Montmaurin (Haute-Garonne), les crânes de Saccopastore près de Rome en Italie (250 000 ans), ou les 800 fossiles découverts par Dragutin Gorjanović-Kramberger à Krapina en Croatie.
Les Néandertaliens classiques
[modifier | modifier le code]Les Néandertaliens les plus typiques, dont les caractères dérivés sont les plus marqués, ont des âges compris entre −100 000 et −30 000 ans, date de leur disparition.

Parmi les fossiles de Néandertaliens classiques, outre les vestiges de Néandertal même (environ −42 000 ans), il faut mentionner les squelettes de La Chapelle-aux-Saints 1, du Moustier, de La Ferrassie[37], de La Quina, de Saint-Césaire dans le Sud-Ouest de la France ou de Spy en Belgique pour ne citer que les plus complets.
Les derniers Néandertaliens connus ont été découverts notamment au Portugal, en Espagne (Zafarraya, −30 000 ans[38],[39]), en Croatie (Vindija, −32 000 ans[40],[41]) et dans le Nord-Ouest du Caucase (Mezmaiskaya, −29 000 ans). Toutes ces dates sont toutefois à considérer avec précaution, les réévaluations successives ayant tendance à vieillir les résultats obtenus par le carbone 14 pour le Paléolithique moyen[42],[43].
Des recherches conduites de 1999 à 2005 dans la grotte de Gorham à Gibraltar suggèrent que les Néandertaliens y ont vécu jusqu'à −28 000 ans, voire −24 000 ans[44],[45]. Ils auraient donc longuement cohabité avec les Homo sapiens, présents dans la région depuis 34 000 ans. Ces résultats sont toutefois fortement critiqués, par exemple par le paléoanthropologue Joao Zilhão, de l'université de Bristol[46].
En 2011, une équipe internationale publia des travaux concernant le site de Byzovaïa, près du cercle arctique en Russie, où ont été découverts des bifaces taillés typiques de la culture moustérienne, classiquement associée aux Néandertaliens en Europe occidentale. Ces outils datent d'il y a 34 000 à 31 000 ans et sont situés plus de mille kilomètres au nord du site le plus septentrional connu pour l'homme de Néandertal, remettant en question la distribution maximale de celui-ci[47],[48]. Ces conclusions ont toutefois été vivement contestées dans une publication ultérieure[49]. Les auteurs considèrent qu'en l'absence de restes fossiles constituant une preuve directe de la présence néandertalienne à cette latitude et à une date aussi récente, l'hypothèse d'un rattachement de l'industrie lithique de Byzovaya au Paléolithique supérieur demeure la plus parcimonieuse.

Caractéristiques physiques
[modifier | modifier le code]Squelette
[modifier | modifier le code]

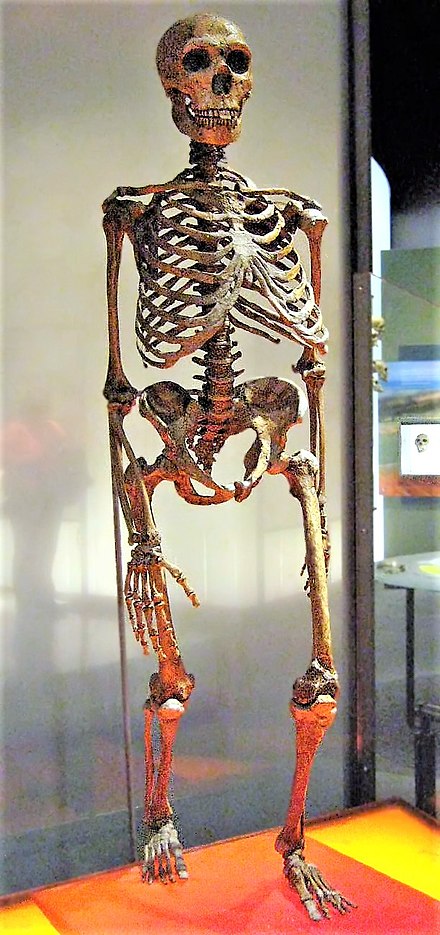
Les Néandertaliens sont de corpulence souvent très massive et robuste : 90 kg et 1,65 m en moyenne pour les hommes et 70 kg et 1,55 m pour les femmes (des individus auraient atteint 1,90 m)[50]. L'ensemble de leur structure (os épais avec corticale développée) et leurs attaches musculaires laissent supposer une grande force physique. Les règles écologiques de Bergmann (corps plus massif qui réduit la déperdition de chaleur) et d'Allen (membres courts qui réduisent également cette déperdition) s'appliquent parfaitement aux néandertaliens dont l'anatomie est une adaptation aux climats froids[51].
Les Néandertaliens présentent quelques caractères archaïques, hérités de leur prédécesseur (caractères plésiomorphes), ainsi que des caractères évolués (caractères apomorphes). Les caractères évolués peuvent être partagés avec les Homo sapiens (caractères synapomorphes) ou bien être des caractères dérivés spécifiques (caractères autapomorphes). Seuls ces derniers permettent d'identifier l'espèce lors de l'examen d'un fossile.
- Les caractères plésiomorphes des Néandertaliens sont :
- la présence d'un épaississement osseux au-dessus des orbites : ce bourrelet sus-orbitaire continu (chez les prédécesseurs des Néandertaliens, ce bourrelet présente un sillon médian, le sulcus supratoralis) forme une visière ;
- un front fuyant ;
- l'absence de menton.
- Les caractères synapomorphes des Néandertaliens sont :
- Les caractères autapomorphes des Néandertaliens sont :
- une face allongée, de forme particulière avec un fort prognathisme méso-facial ;
- des orbites hautes et arrondies ;
- une vaste cavité nasale : le nez saillant, proéminent et grand, avec de larges sinus représente un système thermorégulateur adapté à l'air froid et sec (théorie du radiateur qui réchauffe et humidifie l'air) ; des fosses nasales importantes leur permettent d'inhaler considérablement plus d'air en lien avec des besoins énergétiques plus élevés du fait de leurs corps trapus et de leur activité de chasseur[53] ;
- une arcade dentaire avancée ;
- des pommettes en retrait ;
- le trou auditif externe est ovalaire et se place dans le prolongement de la racine de l'arcade zygomatique (en dessous chez Homo sapiens). Il est fermé par un tympanal à deux versants ;
- la présence d'un espace séparant les dents du fond de la branche montante de la mandibule, dit « espace rétro-molaire » ;
- un crâne dolichocéphale au profil circulaire en vue postérieure (alors que le crâne de tous les autres Hominidés présente un profil pentagonal), l'arrière du crâne en forme de « ballon de rugby » (forme de « ballon de football » chez Homo sapiens) ;
- un os occipital formant une sorte de chignon et présentant une fosse en son centre, dite fosse sus-iniaque.
Les traits spécifiques aux Néandertaliens ont souvent été présentés comme des adaptations au froid ; les membres courts et robustes des Néandertaliens trouvent des analogues modernes dans les populations vivant dans les régions proches du pôle. Des facteurs écologiques liés aux avancées glaciaires tels que l'isolement de populations et le faible brassage génétique ont pu favoriser la fixation rapide de ces traits.
Pigmentation et couleur des cheveux
[modifier | modifier le code]
La rousseur des Néandertaliens a été évoquée à la suite de différentes études mais il semble s'agir de sur-interprétations de résultats scientifiques de la part des médias. Chez les humains, la rousseur est liée à une mutation du gène MC1R (melanocortin-1 receptor) qui régule la production de mélanine[54]. En 2000, une étude a montré que cette mutation pouvait exister depuis 100 000 ans[55]. Certains en ont conclu qu'elle pouvait être apparue chez les Néandertaliens qui l'auraient transmise aux hommes modernes[56], ce que nuancent les auteurs[57].
En octobre 2007, un article de la revue Science présente les résultats d'une étude portant sur l'extraction d'ADN fossile de deux spécimens néandertaliens, l'un découvert en Italie (Monti Lessini), l'autre en Espagne (El Sidrón 1252)[58]. Les auteurs ont amplifié et séquencé un fragment du gène MC1R et ont mis en évidence chez les deux individus une mutation inconnue chez l'homme moderne. Toutefois, il est impossible de déterminer si cette mutation était présente sur les deux allèles et donc si elle affectait le phénotype des individus en question. Pour les auteurs, la présence de ces mutations permet d'estimer qu'un pour cent environ des Néandertaliens avait une pigmentation réduite se traduisant par une peau claire et des cheveux roux[59]. Bien que cette proportion soit très limitée, certains médias ont rapporté que les Néandertaliens étaient roux[60],[61].
Alors que la plupart des musées présentent des reconstitutions de Néandertaliens avec des yeux bleus ou verts, un teint de peau clair et des cheveux roux correspondant à ce que l'on observe chez les populations modernes sous des latitudes équivalentes à celles de l'Europe, une étude génétique parue en 2012 portant sur les ossements de deux femmes néandertaliennes de Croatie suggère la possibilité parallèle d'une peau au teint plus foncé, d'yeux marron et de cheveux bruns[62].
Paléopathologie
[modifier | modifier le code]Une étude de 2024 suggère que Thorin, l'un des derniers Néandertaliens, appartenait à une lignée génétiquement isolée depuis 50 000 ans. Cette isolement et une forte consanguinité pourraient avoir contribué à l'extinction de l'espèce[63].
Les restes osseux de Néandertaliens, tant en Europe qu'en Asie occidentale, présentent parfois des anomalies qui renseignent sur les lésions organiques survenues de leur vivant et parfois responsables de leur décès. Ces anomalies peuvent être classées en quatre catégories principales[64] :
Traumatologie
[modifier | modifier le code]Les Néandertaliens semblent avoir souffert fréquemment de fractures, en particulier au niveau des côtes (Shanidar IV, « vieillard » de La Chapelle-aux-Saints 1), du fémur (La Ferrassie 1), de la fibula (La Ferrassie 2 et Tabun 1), de la colonne vertébrale (Kébara 2[65]) et du crâne (Shanidar I, Krapina, Šaľa 1). Ces fractures sont souvent ressoudées et ne montrent pas ou peu de signes d'infection, ce qui suggère que les individus étaient pris en charge au cours de leur période d'invalidité.
En relation avec des fractures, d'autres traces de traumatismes ont été signalées sur de nombreux squelettes de Néandertaliens. Ils semblent liés à des blessures perforantes, comme chez Shanidar III dont le poumon fut certainement perforé par une blessure entre les côtes 8 et 9. Il peut s'agir d'une attaque intentionnelle ou d'un accident de chasse, mais l'individu survécut à sa blessure durant quelques semaines avant d'être tué par la chute d'un bloc rocheux dans la grotte de Shanidar. D'autres traumatismes correspondent à des coups portés à la tête (Shanidar I et IV[66], Krapina[67]), tous consolidés.
Pathologie articulaire
[modifier | modifier le code]L'arthrose est particulièrement répandue chez les Néandertaliens les plus âgés. Elle concerne de façon spécifique les articulations comme les chevilles (Shanidar III), la colonne vertébrale et les hanches (« vieillard » de La Chapelle-aux-Saints[68],[69]), les bras (La Quina 5, Krapina, Feldhofer), les genoux, les doigts et les orteils, le tout en relation étroite avec les maladies articulaires dégénératives (arthrose), qui peuvent aller de la dégénérescence normale, liée à l’usure, jusqu’à la restriction des mouvements, douloureuse et handicapante, et à la déformation. C’est ce qu’on observe à des degrés divers sur les squelettes de Shanidar (I-IV).
Pathologie dentaire
[modifier | modifier le code]L'hypoplasie de l'émail dentaire est l'indicateur d'un stress survenu durant le développement des dents. Les striations et les cannelures de l'émail reflètent les périodes de pénurie alimentaire, les traumatismes ou les maladies. Une étude de 669 couronnes dentaires de Néandertaliens a montré des signes d'hypoplasie plus ou moins prononcés sur 75 % d'entre elles[70]. Les carences alimentaires en étaient la cause principale, pouvant aller jusqu'à entraîner la perte des dents. Les dents appartenant aux squelettes les plus âgés présentaient toutes une hypoplasie, particulièrement nette chez le « vieillard »[71] de La Chapelle-aux-Saints 1 et l'individu 1 de La Ferrassie.
Pathologie infectieuse
[modifier | modifier le code]On trouve occasionnellement sur des squelettes de Néandertaliens des lésions osseuses secondaires liées à une infection des tissus mous du voisinage. Shanidar I présente des traces manifestes de lésions dégénératives de même que La Ferrassie 1, où les lésions sur les deux fémurs, les tibias et les fibulas indiquent une infection systémique ou peut-être un cancer.
Langage et parole
[modifier | modifier le code]L'aptitude physique à la parole et au langage des Néandertaliens a longtemps été controversée. Les discussions portent, en particulier, sur l'aptitude physique des Néandertaliens au langage, pour laquelle la morphologie de l'os hyoïde est importante[72],[73]. L'os hyoïde est un petit os qui maintient la base de la langue. Il est présent chez tous les mammifères. Très peu d'os hyoïdes de Néandertaliens ont été mis au jour : un premier a été découvert en 1983 dans la grotte de Kébara, sur le mont Carmel en Israël (60 000 ans A.P.) et un autre dans le site d’El Sidron en Espagne (43 000 ans A.P.). Les deux os sont très peu différents de ceux des humains actuels[74],[75],[76]. Des os hyoïdes appartenant à des pré-néandertaliens ont été découverts dans le site de la Sima de los Huesos à Atapuerca en Espagne (430 000 ans A.P.) ; ils ont également des caractéristiques proches de celui des Homo sapiens[77].
En outre, au-delà de la présence ou non de l'os hyoïde, pour le préhistorien Jean-Paul Demoule, il existe plusieurs éléments permettant d'identifier dans la production d'objets des Homo erectus tardifs et des premiers Néandertaliens les preuves de la présence d'un « proto-symbolisme » indice probable d'un prélude de langage se situant entre les sons signifiants des chimpanzés et le langage de l'homme moderne[78].
Concernant le conduit vocal des Néandertaliens, Philip Lieberman a émis en 1971, sur la base d'un travail de modélisation, que ceux-ci ne disposaient pas d'un pharynx de taille suffisante pour produire tous les sons que l'on observe dans les langues du monde. Malgré de nombreuses critiques concernant cette argumentation, cette théorie s'est largement diffusée pendant une trentaine d'années. À la suite d'une longue controverse[79],[80],[81], il semble que les arguments avancés par Lieberman ne soient plus tenables à la lueur des connaissances actuelles. La reconstruction anatomique du conduit vocal qu'il avait utilisée n'était pas réaliste et ses simulations peu convaincantes. Ce n'est pas la taille du pharynx qui permet de parler mais la maîtrise des articulateurs (cordes vocales, langue, mandibule, voile du palais, lèvres). Les nouvelles simulations montrent bien que les Néandertaliens avaient la capacité physique de parler[réf. souhaitée].
Une étude publiée en 2007[82] et portant sur l'analyse de l'ADN provenant des restes de deux Néandertaliens découverts dans la grotte d'El Sidrón (Espagne) aurait permis d'y détecter la même version de la protéine Forkhead-P2 (forkhead box P2) que celle présente chez les hommes modernes. Cela pourrait plaider en faveur de l'aptitude des Néandertaliens au langage puisqu'on estime que ce gène joue un rôle important dans le développement des parties du cerveau liées à la maîtrise du langage articulé[83].
Une étude publiée en 2021 démontre que les capacités auditives des Néandertaliens étaient les mêmes que celles des humains actuels[84].
Culture et techniques
[modifier | modifier le code]
Après avoir longtemps été considéré comme un être archaïque et encore proche de l'animalité, y compris par une partie de la communauté scientifique, l'Homme de Néandertal commence à apparaître comme un être doté de capacités intellectuelles et de traditions culturelles.
Le tableau ci-dessous liste un certain nombre de comportements considérés comme modernes et courants chez Homo sapiens ainsi que la fréquence relative de leur mise en œuvre par Homo neanderthalensis[85] :
| Comportement | Fréquent | Occasionnel | Absent | Incertain |
|---|---|---|---|---|
| Art | ✓ | |||
| Utilisation de pigments | ✓ | |||
| Fabrication de parures | ✓ | |||
| Sépultures | ✓ | |||
| Rites funéraires | ✓ | |||
| Échanges à longue distance | ✓ | |||
| Production de microlithes | ✓ | |||
| Production de pointes barbelées | ✓ | |||
| Production d'outils en os | ✓ | |||
| Production de lames | ✓ | |||
| Utilisation d'aiguilles | ✓ | |||
| Exploitation des ressources marines | ✓ | |||
| Chasse aux oiseaux | ✓ |
Artisan de la pierre et de l'os
[modifier | modifier le code]
L'homme de Néandertal est l'auteur d'un outillage complexe et élaboré, et notamment des industries du Moustérien. Ses méthodes de débitage apportent en outre la preuve de ses capacités d'abstraction et d'anticipation, en particulier en ce qui concerne le débitage Levallois. Les éclats obtenus par cette méthode ou par d'autres pouvaient être utilisés bruts ou bien retouchés, légèrement modifiés sur leurs bords pour obtenir des outils plus spécialisés tels que les racloirs ou les denticulés.
Il est probable que les derniers Néandertaliens soient les auteurs du Châtelperronien, un faciès culturel de transition entre le Paléolithique moyen et le Paléolithique supérieur en Europe occidentale. Ce faciès est caractérisé par des comportements longtemps considérés comme propres aux hommes modernes : débitage de lames, utilisation de parure, fabrication d'outils en os, etc.[86],[87].
Des outils en os ont été mis au jour dans les deux sites néandertaliens de Chagyrskaya (Altaï sibérien) et de Chez-Pinaud-Jonzac (Charente-Maritime, France). À Jonzac, les outils en os sont aussi nombreux que ceux en silex, et très divers : percuteurs, couteaux, racloirs, ciseaux, lissoirs. Il s'agit d'une véritable industrie, destinée à des activités variées et nécessitant différentes mises en œuvre. Les outils ont été identifiés à partir des traces de fabrication et d'utilisation présentes en surface, mais aussi à l'intérieur par microtomographie aux rayons X. Ils ont été fabriqués essentiellement par fracturation, à la différence de ceux d'Homo sapiens, mis en forme principalement par raclage et abrasion[88],[89].
Artisan du bois
[modifier | modifier le code]Des preuves directes (traces d'adhésif naturel en bitume ou en résine[90]) ou indirectes (répartition des traces d'utilisation) montrent que certains outils en pierre étaient utilisés emmanchés. Les manches eux-mêmes, réalisés en matériaux périssables, n'ont pas été conservés. En revanche, des conditions particulièrement favorables ont permis la conservation de quelques objets en bois. Le plus spectaculaire est sans conteste un fragment d'épieu en bois d'if fiché dans le thorax d'un éléphant (Palaeoloxodon antiquus), mis au jour à Lehringen (Basse-Saxe, Allemagne) en 1948. Dans le même site, daté de l'Éémien (130 000-115 000 ans AP), sont découverts des éclats Levallois ayant servi à découper de la peau et de la viande[91],[92].
En 1995, des ouvriers d'une mine de charbon à ciel ouvert proche de Schöningen (également en Basse-Saxe) découvrent des lances en épicéa longues de deux mètres ainsi que d'autres objets en bois incrustés dans la rive d'un ancien lac. Les fouilles de Schöningen révèleront ensuite d'autres restes de bois minutieusement sculptés, notamment des bâtons de jet et d'autres instruments de chasse, ainsi que des dizaines d'outils non destinés à la chasse, datés de 300 000 ans. Le site n'a pas livré d'ossements d'hominidés mais la date est celle des premières preuves d'existence des Néandertaliens ailleurs en Europe : les fabricants d'outils étaient sans doute des représentants des premiers Néandertaliens ou peut-être des derniers Homo heidelbergensis. De nombreux ossements d'animaux ne laissent aucun doute sur le fait qu'ils chassaient des chevaux[93].
Grand chasseur
[modifier | modifier le code]
En 1981, l'archéologue Lewis Binford soutient une théorie selon laquelle les premiers hominidés (dont les Néandertaliens) jusqu'au Paléolithique moyen ne pratiquaient que la cueillette ou le charognage passif, seuls les hommes modernes pratiquant la chasse de grand gibier rapide[94]. Cette théorie est aujourd'hui abandonnée.
Rares sont les preuves directes de la pratique de chasse aux grands herbivores par les Néandertaliens telles que le fragment d'épieu en if de Lehringen et les lances de Schöningen à la pointe parfois durcie au feu[95] ou des pointes emmanchées en silex ou en os, parfois collées au bitume[90]. En revanche, les sites livrent des accumulations impressionnantes d'ossements de grands mammifères (bison à Coudoulous, Lot et à Mauran, Haute-Garonne ; saïga en Crimée ; bouquetin dans la grotte du Lazaret ; aurochs à La Borde, Lot ; cheval à Saint-Césaire, Charente-Maritime) ; elles sont interprétées comme le résultat de chasses saisonnières, parfois avec utilisation d'avens ou de fondrières comme pièges naturels (technique de « chasse à l'abîme »)[96]. De plus, les analyses biogéochimiques sur le collagène osseux des Néandertaliens et des mammifères associés[97] montrent une alimentation carnée proche de celle du loup, même en période tempérée. Chasseurs de grands mammifères, les Néandertaliens avaient des stratégies (communautés spécialisées dans la chasse de deux ou trois espèces, technique à l'approche ou à la poursuite de proies en fonction de l'âge et du sexe) qui attestent une parfaite connaissance de l'environnement et de l'éco-éthologie des animaux et un savoir-faire technique développé. Enfin les études anatomiques montrent leur adaptation à la chasse : ils étaient en effet trapus et musclés[98].
La pratique ponctuelle d'un charognage actif (accès primaire à la carcasse en écartant les prédateurs — hyène des cavernes, loup, lion des cavernes — ou en recherchant les animaux morts dans des pièges naturels) a également été évoquée, notamment pour les grands mammifères (mammouth, rhinocéros laineux)[99],[100].
Société néandertalienne
[modifier | modifier le code]
Les analyses en paléogénétique réalisées en 2010 dans la grotte d'El Sidrón suggèrent que la société néandertalienne pratique l'exogamie patrilocale[101].
Certaines constatations de comportements altruistes témoignent de manifestations d'entraide, de solidarité et d'assistance, tel le vieillard retrouvé dans le cimetière de La Chapelle-aux-Saints ou de Shanidar avec un squelette si déformé par la maladie qu'il devait probablement être infirme[102]. Les enfants s'entraînent à la taille d'outils lithiques sur les mêmes sites que les adultes expérimentés[103].
Au moins un comportement alimentaire, le bris des os pour en extraire la moelle, a révélé un apprentissage social plutôt que technique dans les niveaux du Pléistocène moyen. Une étude sur les assemblages de ces matériaux provenant de la grotte de Bolomor (Valence, Espagne, MIS 9-5e) et celle de Gran Dolina TD10-1 (Burgos, Espagne, MIS 9) : dans un même groupe, les os ne sont pas brisés aux points les plus faibles, dans une logique d'efficacité, mais suivant un comportement socialement acquis. Au sein d'une même grotte, ces comportements changent au fil des époques ; cela suggère l'existence possible d'identités culturelles ou de prédispositions comportementales dépendant des groupes, et pourrait servir de marqueur pour les zones d'extension des différents groupes[104].
Les analyses génétiques suggèrent que les Néandertaliens de Sibérie vivaient dans des populations relativement isolées de moins de 60 individus. En revanche, les Néandertaliens d'Europe et les humains modernes anciens semblent avoir vécu dans des populations de plus grande taille[105].
Un amoncellement d'ossements et d'outils découverts dans la mine de charbon de Neumark-Nord, près de Halle-sur-Saale (Saxe-Anhalt, Allemagne), montre que les Néandertaliens pouvaient se rassembler par centaines, au moins temporairement. Au cours d'environ trois siècles pendant l'Éémien, il y a environ 125 000 ans (75 000 ans avant l'arrivée des premiers Sapiens), ils ont abattu au même endroit plus de 70 éléphants (Palaeoloxodon antiquus), récoltant jusqu'à 4 tonnes de chair de chaque animal. Sur les 3 400 os d'éléphants, presque tous montrent des signes de dépeçage à l'aide d'outils tranchants, et aucun n'a été rongé par des charognards, signe qu'ils avaient été parfaitement nettoyés. La viande d'un seul éléphant a pu suffire à nourrir 350 personnes pendant une semaine, ou 100 pendant un mois s'ils étaient capables de conserver la viande aussi longtemps[106],[107].
Alimentation
[modifier | modifier le code]Les Néandertaliens ont d'abord été considérés comme exclusivement chasseurs carnivores, faisant partie des superprédateurs (mangeant de grands herbivores alors qu’Homo sapiens avait diversifié son alimentation — petit gibier, fruits de mer, poissons d'eau douce et marins). Ce régime alimentaire a parfois été considéré comme l'une des causes de leur extinction[108].
En 2010, des analyses de phytolithes piégés dans des plaques de tartre de dents fossilisées néandertaliennes provenant de différents sites ont révélé des traces de plantes fossilisées (palmier-dattier, légumineuses, rhizomes de nénuphar, graminées du genre Triticum ou Hordeum), indiquant un régime alimentaire diversifié et un comportement de chasseur-cueilleur. De plus, certains grains d'amidon retrouvés montrent des processus de cuisson, suggérant que les Néandertaliens, grâce à leur maîtrise du feu, cuisaient ces végétaux en les faisant bouillir. On pensait auparavant que seules les viandes étaient cuites (d'après l'analyse des ossements d’animaux retrouvés dans de nombreux foyers) alors que les végétaux étaient simplement grillés[109]. Les Néandertaliens ont ainsi une alimentation carnée composée essentiellement de grands mammifères mais incluant également des végétaux ou de petits animaux (lagomorphes, oiseaux, mollusques terrestres[110]) lorsque les conditions s'y prêtent. Les produits d'origine animale et végétale constituent respectivement 80 et 20 % de leur régime alimentaire[111],[112].
Des restes de coquillages trouvés dans la grotte de Bajondillo dans le Sud de l'Espagne, montrent que des Néandertaliens mangeaient des fruits de mer en Europe dès 150 000 ans avant le présent[113],[114].
En 2012 puis en 2017, l'analyse chimique puis l'identification de restes d’ADN piégés il y a environ 36 000 ans dans la plaque dentaire de Néandertaliens découverts en Belgique et en Italie montrent qu’ils mangeaient notamment du rhinocéros laineux, du mouflon et des champignons. Par contre, chez ceux d'El Sidrón (Espagne), datés d'environ 48 000 ans, seul de l’ADN végétal a été détecté. Il provient de diverses plantes et champignons (pignon de pin et mousse forestière notamment) [115]. Les Néandertaliens semblent aussi avoir utilisé des analgésiques (acide salicylique trouvé dans le saule) et des antibiotiques naturels (Penicillium)[115].
En 2012, l'analyse chimique de plaques dentaires de cinq Néandertaliens mis au jour à El Sidrón avait déjà montré la présence de composés végétaux amers pouvant provenir de l'achillée millefeuille, dont l'utilisation était déjà présumée précédemment par la présence de pollens dans une tombe néandertalienne à Shanidar, en Irak[116], et la camomille. Il s'agit de plantes sans valeur nutritive mais présentant des vertus médicinales ou pouvant servir éventuellement de coupe-faim.
En 2019, l'étude des dépôts moustériens de la grotte de Pié Lombard (Tourrettes-sur-Loup, Alpes-Maritimes, France) révèle les restes fossilisés d'au moins 225 lapins de l'espèce Oryctolagus cuniculus, de loin l'espèce animale la plus abondamment représentée sur le site[117].
Par ailleurs, les Néandertaliens chassaient des animaux marins tels que le dauphin ou le phoque. Globalement, les découvertes montrent de plus en plus que leur régime alimentaire était qualitativement aussi diversifié que celui des humains anatomiquement modernes[118],[119].
De plus, lors d'une étude de 2022, John McNabb (Centre for the Archaeology of Human Origins de l’Université de Southampton au Royaume-Uni) met en exergue la découverte du site préhistorique de la Grotte de Shanidar en Irak qui révèle que Néandertal faisait cuire ses aliments et utilisait des mélanges de diverses légumineuses avec de la viande[120].
Par ailleurs, les Néandertaliens faisaient cuire des crabes[121].
Sépultures
[modifier | modifier le code]
S'il est possible qu’Homo heidelbergensis, l'un des ancêtres probables de l'Homme de Néandertal, ait adopté un comportement particulier vis-à-vis de ses morts à Atapuerca, les premières véritables sépultures connues sont néandertaliennes[122]. Les plus anciennes datent d'environ - 100 000 ans et ont été mises au jour au Proche-Orient. Elles se multiplient ensuite et on en trouve en France (La Chapelle-aux-Saints, La Ferrassie, La Quina, Le Moustier, Saint-Césaire), en Belgique (Spy), en Israël (Kébara, Amud), au Kurdistan irakien (Shanidar), en Ouzbékistan (Teshik-Tash). Dans certains cas, elles comprennent des dépôts funéraires (outils lithiques, fragments de faune).
L'une des sépultures de Shanidar renfermait un Néandertalien enterré sous une grande dalle. Une grande quantité de pollens de plantes à fleurs était présente autour du corps[123],[124],[125],[126]. Ces pollens ont longtemps été considérés comme la preuve du dépôt de nombreuses fleurs lors de l'enfouissement. Une étude de 1999 a remis en question cette interprétation, citant des phénomènes post-dépositionnels ou l'action de rongeurs pour expliquer l'accumulation de pollens. Cette étude a cependant été fortement contestée[127].
Ces sépultures comportent souvent des fosses intentionnelles et sont pratiquement toujours associées à des habitats. Il est peu probable qu'elles n'aient eu qu'un rôle fonctionnel simplement destiné à se débarrasser d'une dépouille, même si leur interprétation en termes de religiosité est sujette à discussion.
En 2020, en Dordogne, le squelette d’un enfant de 2 ans est considéré comme ayant été enterré au cours d'un rite funéraire[128].
Les Néandertaliens ont plusieurs types de rites funéraires : sépultures, inhumations en deux temps après décomposition du corps à l'air libre (traces de désarticulation et décharnement à Shanidar VI et VIII, Krapina, Kébara[129], combe Grenal[130]), « culture des ancêtres » (analyses de taphonomie sur des crânes isolés et corps sans tête à Kébara), endocannibalisme (Moula-Guercy à Soyons, Ardèche[131],[132], Vindija et Krapina en Croatie : stries de fracturation sur os frais)[133].
Culte de l'ours
[modifier | modifier le code]Dans certains sites tels que celui du Regourdou en Dordogne, des accumulations de crânes d'ours qui semblaient disposés intentionnellement ont été interprétées comme le résultat d'un « culte de l'ours ».
Au Regourdou, un squelette d'ours brun reposait sous une dalle monolithe d'un poids de 850 kg, dans une fosse peu profonde. À proximité, le corps d'un Néandertalien était couché sur le côté gauche, la tête vers le nord, en position fœtale. Le crâne manquait, mais il restait la mandibule. L’absence du crâne a également été observée dans le cas de la sépulture néandertalienne de Kébara. D'après E. Bonifay, il s'agissait d'une véritable tombe composée d’une fosse dallée, empierrée et couverte de sable et de cendres de foyer[134],[135]. Cette interprétation a largement été remise en question depuis, les accumulations d'ossements d'ours du Regourdou pouvant être liés à des phénomènes taphonomiques liés à l'occupation de la cavité par des ours hibernants[136].
L'existence du culte de l'ours, popularisée par les romans préhistoriques de l'écrivaine américaine Jean M. Auel, est contestée par de nombreux scientifiques. Les crânes d'ours sont extrêmement résistants et peuvent être déplacés par des phénomènes naturels jusqu'à acquérir des positions évoquant une organisation volontaire mais en fait seulement due au hasard[réf. à confirmer][137].
Cannibalisme et traitement rituel des cadavres
[modifier | modifier le code]La présence de traces de désarticulation, de décharnement, de fracturation intentionnelle ou de calcination sur certains os de Néandertaliens a été interprétée comme un témoignage de la pratique du cannibalisme. Des ossements de sites tels que l'abri Moula[138], en Ardèche ou Krapina[139],[140],[141] en Croatie présentent de telles traces de découpe. Il est toutefois difficile de démontrer s'il s'agit de cannibalisme plutôt que d'un traitement post mortem des dépouilles dans le cadre d'un rite funéraire. Les fragments d'os de Krapina présentent des marques comparables à celles de sépultures secondaires d'une nécropole du XIVe siècle découverte dans le Michigan, correspondant à l'ablation de la chair sur une dépouille partiellement décomposée. Certains os crâniens du site des Pradelles à Marillac-le-Franc présentent des traces de découpe correspondant sans doute au prélèvement du cuir chevelu par scalpation.
Le crâne de Néandertalien découvert dans la grotte Guattari (Mont Circé, Italie) a longtemps été considéré comme une preuve irréfutable de rituel anthropophagique : il aurait été déposé dans un cercle de pierre après que le trou occipital avait été élargi pour consommer le cerveau. Des examens approfondis ont montré que le cercle de pierre était probablement naturel et que l'élargissement du trou occipital avait été causé par une hyène, ce que confirme la présence de traces de dents en différents points du crâne[142],[143].
Cependant, des os humains ont été découverts intentionnellement cassés dans le but d'exploiter la moelle. Ainsi dans la grotte de l'Hyène à Arcy-sur-Cure (Yonne), des os humains étaient mélangés sans aucune distinction aux os d'animaux ayant servi de nourriture, le tout encerclant le principal lieu de vie[144]. Ces découvertes permettent de retenir l'hypothèse du cannibalisme comme fort probable, sans pouvoir trancher s'il s'agit d'un endocannibalisme ou d'un exocannibalisme[145].
Arts
[modifier | modifier le code]

La reconnaissance progressive de la culture néandertalienne remet en cause la primauté culturelle de l'homme moderne : alors que l'on pensait il y a peu que la culture technique et symbolique des Néandertaliens était très nettement inférieure quantitativement et qualitativement à celle de l'Homo sapiens, les découvertes récentes font apparaître que l'Homme de Néandertal avait lui aussi développé certaines techniques évoluées (débitage de lames[146]), et développé ou adopté des traits culturels modernes (sépultures, signes gravés, parures). La thèse du rôle capital de l'arrivée de l'Homo sapiens en Europe et celle d'une corrélation entre l'évolution biologique et l'évolution culturelle expliquant le plus grand développement de l'Homo sapiens par son évolution biologique s'en trouvent donc remises en question.
Au Paléolithique moyen apparaissent les premières manifestations de préoccupations esthétiques ou symboliques :
- collecte de fossiles ou de minéraux rares ;
- utilisation d'ocre (des utilisations fonctionnelles ont été évoquées, sans preuves, mais l'utilisation sur des peaux est avérée)[147] ;
- utilisation de plumes, comme le montre la disposition des traces de découpe sur les ossements d'oiseau de la grotte de Fumane en Italie[148] ;
- gravure de traits, de lignes ou de signes géométriques simples sur des os ou des pierres[149],[150].
En 2014, la découverte dans la grotte de Gorham (Gibraltar) de formes géométriques gravées sur une paroi recouverte de sédiments datant de plus de 39 000 ans est annoncée par l'équipe de Clive Finlayson (en). Elles constituent le premier exemple connu d'art pariétal abstrait attribué aux Néandertaliens. Leur réalisation a nécessité plusieurs centaines de passages de la pointe d'un outil de pierre taillée, probablement de silex[151],[152].
En février 2018, de nouvelles datations viennent confirmer l'origine néandertalienne de créations artistiques (pourtour de main, réseau de lignes, peinture sur paroi) découvertes dans trois grottes espagnoles (grotte de La Pasiega (es), grotte de Maltravieso (es) et grotte d'Ardales (es)) : plus de 64 800 ans, soit plus de 20 000 ans avant l'arrivée en Europe des premiers hommes modernes. Des coquillages percés et teints, trouvés dans une quatrième grotte, sont encore plus vieux[153],[154],[155],[156]. Néanmoins, un article paru le 21 septembre 2018 remet en cause la datation des grottes de Maltravieso et de La Pasiega en raison du manque de corrélation entre le matériau analysé (des efflorescences de calcite) et le phénomène à dater. Quant aux aplats d'oxydes rouges trouvées à Ardales, rien ne prouve leur origine humaine[157],[158].
L'étude des pigments de la grotte du Renne à Arcy-sur-Cure (Salomon et al. 2008) permet de conclure que les Néandertaliens ont utilisé des couleurs sur les peaux (vêtements, tentes) bien avant qu'elles n'aient été utilisées pour les peintures rupestres. Les mêmes auteurs, et d'autres, notent la grande quantité de pigments remontant au Moustérien de tradition acheuléenne dans les grottes du Pech-de-l'Azé à Carsac-Aillac, Dordogne[159], et à d'autres sites de cette époque.
En 2015 un réexamen des serres d'un pygargue à queue blanche, découvertes en 1899 sur le site néandertalien de Krapina en Croatie, révèle un remodelage en vue d'en faire les plus anciens bijoux connus, datés de 130 000 ans. Il précède de 20 000 ans les plus anciens bijoux connus créé par l’Homo sapiens à Skhul en Israël. Les Néandertaliens possédaient donc une culture symbolique 80 000 ans avant l'arrivée d’Homo sapiens en Europe. Des griffes similaires qui datent de 100 000 ans avaient été trouvées au site néandertalien des grottes du Pech-de-l'Azé en France[160]. Sur le même site, des pigments étaient utilisés sur les cuirs travaillés (vêtements, tentes), prouvant que bien avant l'apparition en Europe des peintures rupestres les Néandertaliens savaient manier les matériaux colorants et qu'ils n'ont pas attendu l'Homo sapiens pour donner à leurs objets du quotidien une dimension symbolique[159].
Une controverse scientifique entoure la possible « flûte de Divje Babe », datée d'environ 43 000 ans AP, dans un environnement culturel de style et d'époque moustériens, et présentée au Musée national de Slovénie (Narodni Muzej Slovenije) à Ljubljana comme le plus vieil instrument de musique découvert à ce jour, et éventuellement attribuable à l'Homme de Néandertal. Le découvreur — ou plutôt l'inventeur — de l'objet est le paléoanthropologue Ivan Turk ; il est aussi l'auteur de l'hypothèse selon laquelle il s'agit bien d'une ancienne flûte néandertalienne[161].
Les arguments en faveur et en défaveur de cette hypothèse sont nombreux[162]. Si une nouvelle preuve décisive intervient à l'avenir dans ce débat, cet objet serait la première preuve d'une pratique musicale chez l'Homme de Néandertal. Mais à ce jour (2023), cette controverse au sujet de la nature d'artéfact humain de l'objet ou bien façonné au hasard de l'action de carnivores, ainsi que sur son caractère néandertalien, n'est pas encore tranchée.
Structures de la grotte de Bruniquel
[modifier | modifier le code]
Deux structures annulaires formées par l'accumulation de tronçons de stalagmites ont été découvertes en 1995 dans la grotte de Bruniquel, mais les campagnes de prélèvements n'ont pu avoir lieu qu'à partir de 2014. Ces structures, constituées de presque 400 « spéléofacts », ont été datées en 2016 : 176 500 ± 2 000 ans. Cet âge indique qu'elles sont vraisemblablement l’œuvre de l'homme de Néandertal, et même de néandertaliens « archaïques »[163],[164].
Fabrication de cordage
[modifier | modifier le code]En 2020, une équipe internationale a publié une étude décrivant le moulage fossilisé d’un fragment de cordelette torsadée, composée de trois brins eux-mêmes torsadés. Cette découverte a été faite dans l’abri du Maras en Ardèche, daté entre 52 000 et 41 000 ans AP. L’Homme de Néandertal ayant été le seul occupant de ce site, cet artéfact lui a été attribué. Les fibres ont été identifiées comme provenant probablement d’un conifère. C'est la plus ancienne trace de cordage trouvée, la précédente datant de 19 000 ans AP provenant du site Ohalo II[165].
Hybridations
[modifier | modifier le code]Morphologies d'apparence hybride
[modifier | modifier le code]Un squelette d'enfant découvert au Portugal a été présenté en 1999 comme un hybride Néandertal/Homo sapiens. Cette hypothèse est fortement discutée en raison des difficultés à identifier les caractères autapomorphiques chez un individu juvénile, alors que la variabilité de la population concernée est mal connue[33].
Un squelette d'enfant trouvé en contexte gravettien à Lagar Velho dans la vallée de Lapedo, au centre du Portugal, porterait des caractéristiques des deux espèces[166],[167],[168]. Cet enfant d'environ quatre ans a été inhumé dans une sépulture intentionnelle, il y a 25 000 ans. Il est donc postérieur de quelques milliers d'années aux derniers restes clairement attribuables aux Néandertaliens (entre 30 000 et 40 000 ans avant le présent[169],[170]). Cependant le caractère hybride de cet enfant est très discuté et difficile à établir : les caractères dérivés des deux taxons sont moins marqués chez les individus juvéniles que chez les adultes et la variabilité individuelle des enfants de l’époque est absolument inconnue[33].
Le réexamen (morphologie géométrique, analyse paléogénétique de l'ADN mitochondrial) en 2013 d'une mandibule dite des « amants de Vérone » suggère une hybridation entre un père sapiens et une mère néandertalienne[171].
Génétique
[modifier | modifier le code]
En 2010, le séquençage de 63 % du génome de Néandertal effectué par une équipe de l'Institut Max-Planck d'anthropologie évolutionniste coordonnée par Svante Pääbo[20],[172] montre que 1 à 4 % du génome des Homo sapiens non africains provient des Néandertaliens. Les Homo sapiens d'ascendance africaine n'ont, eux, que peu ou pas d'ADN de Néandertal. Cet apport est réparti sur 20 régions génétiques spécifiques aux Homo sapiens.
En 2010, des résultats basés sur l'analyse de 4 milliards de paires de bases d'ADN nucléaire, issus d'ossements fossiles de trois Néandertaliens, ont montré que ceux-ci étaient génétiquement plus proches des Homo sapiens eurasiatiques que de ceux d'Afrique subsaharienne[20]. Les auteurs en concluaient que les Néandertaliens auraient contribué à hauteur de 1 à 4 % (en moyenne 2,2 %) au génome des populations d'humains modernes non africaines[20].
Ces croisements entre Néandertaliens et Homo sapiens eurasiatiques auraient pu survenir il y a 100 000 à 50 000 ans au Proche-Orient[173],[174],[175].
En 2012, une étude confirme que Néandertaliens et humains modernes se sont hybridés « quand les humains modernes, porteurs de technologies proches de celles du Paléolithique supérieur, ont rencontré les Néandertaliens alors qu'ils quittaient l'Afrique »[176],[177].
Si chaque Homo sapiens n'a que 1,8 à 2,6 % du génome de Néandertal en lui, en regroupant les différentes séquences éparpillées dans le monde entier, les chercheurs sont parvenus à reconstituer environ 30 % de ce génome[178],[179] .
En 2014, l'étude du génome d'un Homo sapiens découvert à Kostenki, en Russie, et daté de 37 000 ans avant le présent, confirme encore le métissage et permet d'avancer une date à laquelle l'hybridation aurait eu lieu[180],[181].
En 2015, des analyses génétiques révèlent que le fossile Oase 1 avait un ancêtre récent néandertalien, avec un ADN autosomique néandertalien estimé de 5 à 11 %[182].
En 2017, le génome d'une seconde néandertalienne provenant de la grotte de Vindija (Croatie) a été séquencé. L'étude a permis de préciser la proportion d'ADN néandertalien chez les Eurasiens (1,8 à 2,6 %) et confirme l'implication de ces gènes néandertaliens dans la résistance au froid et la sensibilité à certaines maladies[183],[184].
En 2018, l'ADN d'un fragment osseux trouvé dans la grotte de Denisova, celui d'une adolescente morte vers 13 ans il y a environ 90 000 ans[a], montre qu'elle était l'hybride d'une mère néandertalienne et d'un père dénisovien[185]. C'est le premier cas reconnu d'une hybridation entre espèces humaines au premier degré.
Interfécondité
[modifier | modifier le code]En 2016, une information nouvelle a contribué aux débats : alors que les séquençages précédents concernaient l'ADN autosomal (non sexuel), une première description d'un chromosome Y (et donc masculin) néandertalien (provenant de la grotte d'El Sidrón, Espagne, il y a 49 000 ans) a montré de grandes différences avec le chromosome Y humain actuel[186],[187]. Même si des Néandertaliens et des humains modernes se sont hybridés il y a quelque 55 000 ans, l'ADN du chromosome Y néandertalien ne semble pas avoir été transmis aux humains modernes[187]. Ceci va dans le sens d'études antérieures constatant que si les Asiatiques et les Européens actuels ont bien hérité de 1 % à 3 % de leur ADN de leurs ancêtres via des croisements avec des Néandertaliens, leurs chromosomes Y n'en portent pas de traces. Une hypothèse explicative serait que les deux taxons n'étaient pas entièrement compatibles : le sperme des hommes Néandertaliens pourrait avoir été non fécondant pour les femmes « modernes » de leur époque[188]. On a trouvé dans le chromosome Y néandertalien d'El Sidrón des mutations de trois gènes impliqués dans le système immunitaire, dont celui qui produit des antigènes pouvant provoquer une réponse immunitaire aboutissant à une fausse-couche chez les femmes enceintes[188]. Ainsi, même si des hommes néandertaliens et des femmes « modernes » se sont sexuellement unis par le passé, ils n'ont vraisemblablement pas pu obtenir de descendance mâle suffisamment abondante ou viable, ce qui aurait pu hâter « l'extinction » des néandertaliens.
En 2019, David Reich parle des humains modernes et des Néandertaliens comme de « deux groupes à la limite de la compatibilité », l'analyse de l'ADN ancien et de l'ADN moderne d'Homo sapiens montrant une expulsion progressive des gènes néandertaliens impliqués dans la reproduction. Lors de chaque reproduction, l'héritage génétique se fait au hasard, par recombinaison des génomes paternel et maternel. Et le résultat montre que les Homo sapiens (non-africains) qui portaient le plus de traces néandertaliennes sur les chromosomes reproductifs n'ont pas eu de descendance (ou peu), donc étaient plus touchés par des problèmes de stérilité[189]. Cette stérilité partielle des hybrides est un des arguments en faveur de la définition de Néandertal comme une espèce à part entière.
Gènes transmis par Néandertal à Sapiens
[modifier | modifier le code]Dans le génome des hommes modernes européens, un gène lié à l'immunité pourrait être issu du génome de l'Homme de Néandertal[190],[191],[192]. Cette découverte est confirmée en 2016 par des études qui mettent en évidence ces cas d'introgression : gènes néandertaliens à l'origine de la stimulation immunitaire[193] et des allergies des hommes modernes[194].
Certaines séquences d'ADN acquises par Homo sapiens auprès des Néandertaliens ont pu s’avérer bénéfiques, comme celles favorisant l'adaptation au climat froid ou résistance aux maladies. Elles ont été sélectionnées positivement alors que d'autres, délétères, ont été éliminées du génome humain par sélection négative. On trouve aujourd'hui dans le génome humain des allèles dérivés de Néandertaliens qui nous rendent à la fois sensibles ou résistants à certaines maladies[Lesquelles ?][22].
Parmi les gènes que les néandertaliens nous ont transmis, certains sont associés à des traits phénotypiques ou à des maladies. Ils affectent notamment[22] :
- la concentration plasmatique de :
- cholestérol à lipoprotéines de basse densité (LDL) (rs10490626) ;
- vitamine D (rs6730714) ;
- les troubles de l'alimentation (rs74566133) ;
- l'accumulation de graisse viscérale (rs2059397) ;
- le risque de :
- polyarthrite rhumatoïde (45475795) ;
- schizophrénie (rs16977195) ;
- la réponse aux antipsychotiques (rs1459148).
Cela conforte l'idée « que l'ascendance néandertalienne influe sur le risque de maladie chez les humains actuels, en particulier en ce qui concerne les phénotypes neurologiques, psychiatriques, immunologiques et dermatologiques »[22],[195].
Gènes transmis par Sapiens à Néandertal
[modifier | modifier le code]En 2017, une étude de Cosimo Posth, analysant un fémur de Néandertalien vieux de 124 000 ans, a montré que cet os contenait déjà des gènes caractéristiques d'Homo sapiens. L'étude concluait que les premiers croisements entre Sapiens et Néandertal ont dû avoir lieu il y a 276 000 ans[196].
Une étude identifie une transmission de gènes d'Homo sapiens vers des Néandertaliens, probablement il y a plus de 100 000 ans au Moyen-Orient lors d'une première sortie d'Afrique[197].
Une étude publiée en 2020 analyse les chromosomes Y de Néandertaliens récents. Cette étude nous renseigne donc sur les lignées agnatiques. Le signal renvoyé par ces chromosomes Y diffère grandement de celui de l'ADN autosomal. Les chromosomes Y des Dénisoviens auraient divergé il y a environ 700 000 ans d'une lignée partagée par les chromosomes Y humains néandertaliens et sapiens alors que l'on sait que ce sont les Néandersoviens qui ont divergé de la lignée d'Homo sapiens vers cette date. Les lignées Y néandertalienne et sapiens se seraient séparées il y a environ 370 000 ans. On retrouve des résultats similaires avec l'ADN mitochondrial et donc la lignée cognatique, alors que les Néandertaliens anciens de la Sima de los Huesos, datés de 430 000 ans, avaient un ADN mitochondrial plus proche de celui des Dénisoviens. Ces résultats suggèrent un remplacement, chez les Néandertaliens moyens, des gènes néandertaliens ancestraux par des gènes en provenance de la lignée d'Homo sapiens, et ceci seulement pour les gènes uniparentaux, tant mitochondriaux (lignée maternelle) que du chromosome Y (lignée paternelle). Ce résultat montre des hybridations anciennes avec des Homo sapiens archaïques[198],[199].
En 2024, une étude comparée du génome des deux espèces montre que plusieurs zones de celui de l'Homme de Néandertal ont une origine attribuable à Homo sapiens (des séquences identifiées chez des Africains sans ascendance néandertalienne), et suggère qu'il y a eu au moins deux flux génétiques dans ce sens. Les néandertaliens de la grotte de Vindija et ceux de l'Altaï ont ainsi hérité 53,9 Mb de nucléotides sapiens (2,5 % du génome) et 80,0 Mb (3,7 %), respectivement[200].
Extinction
[modifier | modifier le code]
Les derniers Néandertaliens ont disparu plusieurs milliers d'années après l'arrivée d'Homo sapiens par l'est de l'Europe[201], soit probablement il y a environ 40 000 ans[202],[203]. L'extinction aurait été échelonnée dans le temps selon un gradient régional[169]. Leur disparition a suscité de nombreuses hypothèses[204],[205],[206], certaines faisant intervenir des modèles mathématiques[207] ou économiques[208],[209]. Cette disparition des Néandertaliens coïncide avec l'arrivée en Europe de groupes d'hommes modernes[b], porteurs d'une nouvelle culture matérielle, l'Aurignacien.
Causes d'extinction possibles
[modifier | modifier le code]Éruption volcanique
[modifier | modifier le code]Entre 2010 et 2017 ont été mises en relation la disparition de l'Homme de Néandertal et l'éruption campanienne, à l'origine de la caldeira des champs Phlégréens (baie de Naples, Italie)[210]. Des datations de cet événement publiées en 2003 donnent une date de 39 280 ±110 ans AP[211].
Cet événement a vu la libération dans l'atmosphère d'un volume de matières volcaniques extrêmement important, d'au moins 300 km3[211]. Le panache de cendres serait monté à plus de 30 km[212], voire 40 km[réf. nécessaire] d'altitude (stratosphère), avant de retomber en un épais manteau[213] recouvrant tout, comparable aux couches de cendres de Pompéi, sur une aire de 3 700 000 km2[212] allant de l'Italie aux steppes asiatiques, tuant toute forme de végétation sur cette zone, ainsi que toute la chaîne trophique en dépendant, grands prédateurs et Homo compris. L'atmosphère globale aurait été assombrie pendant plusieurs années.
Les derniers peuplements néandertaliens ayant survécu à cet évènement (présence constatée sur la péninsule Ibérique pendant encore plusieurs millénaires) auraient fini de disparaître en raison d'un manque de diversité génétique (garante de la viabilité d'une espèce).
Extinction due à des capacités cognitives inférieures
[modifier | modifier le code]En 2010, une équipe de paléontologues du département d'évolution humaine de l'Institut Max Planck, à Leipzig (Allemagne), a analysé l'endocrâne (l'empreinte laissée par le cerveau dans la boîte crânienne) de squelettes de néandertaliens. Les résultats ont confirmé les données sur la génétique des Néandertaliens acquises grâce au séquençage de leur ADN : le cerveau des Homo sapiens a une forme globulaire caractéristique qui n'existe pas chez Néandertal. Cette différence pourrait influencer l'organisation neuronale et synaptique du cerveau, et donc les capacités cognitives. Par une litote, Jean-Jacques Hublin, le chercheur français qui a dirigé cette étude, émet l'hypothèse que « l'homme de Néandertal ne voyait pas le monde de la même façon que nous » et que son incapacité à créer des liens sociaux complexes serait à l'origine de son extinction[214].
Extinction par stérilité de l'hybridation
[modifier | modifier le code]Selon une autre hypothèse, la disparition progressive de la population néandertalienne serait liée à la possibilité d'accouplements féconds mais donnant des hybrides stériles, au moins chez les Néandertaliennes. Une telle hypothèse, émise par le paléontologue finlandais Björn Kurtén[215], demeure difficile à tester.
Certaines études suggèrent que les enfants issus d'union entre Néandertaliens et Homo sapiens auraient été moins fertiles, ce qui aurait entraîné une diminution de la proportion de gènes hérités de Neandertal[216].
Extinction par consanguinité et petitesse de la population
[modifier | modifier le code]La diversité génétique de l'espèce semble s'être fortement réduite au cours du Pléistocène supérieur, comme le montrent les analyses faites sur les séquences connues entre 42 000 et 29 000 ans, au moment où Néandertal cohabitait avec l’Homo sapiens. Ce constat nourrit la thèse du déclin démographique de Néandertal sur cette période, déclin conduisant, par un phénomène de goulet d'étranglement de population, à la disparition progressive de certains génotypes, donc à l'appauvrissement génétique de l'espèce.
Ont également été évoqués des problèmes d'ordre génétique liés à une forte consanguinité et/ou des mutations spontanées ayant entraîné des maladies congénitales telles que l'hémophilie, le diabète insulino-dépendant ou une forme de stérilité, ayant suffisamment affecté la démographie de la population pour la faire disparaître.
Partant de l'observation que les populations néandertaliennes étaient déjà petites avant l'arrivée de l'Homme moderne, une étude publiée en 2019 met en œuvre trois facteurs que la biologie de la conservation identifie comme essentiels pour la persistance d'une petite population, à savoir la consanguinité, l'effet Allee et la stochasticité. Les résultats indiquent que la disparition des Néandertaliens peut avoir résidé dans la petitesse de leur seule population. Ainsi, même s'ils étaient identiques dans leurs traits cognitifs, sociaux et culturels, et même en l'absence de compétition interspécifique, les Néandertaliens couraient un risque d'extinction considérable. En outre, les auteurs suggérent que si les humains modernes ont contribué à la disparition des Néandertaliens, cette contribution n'aurait peut-être rien à voir avec la concurrence des ressources, mais plutôt avec la façon dont les populations entrantes ont restructuré géographiquement les populations résidentes, d'une manière qui a renforcé les effets Allee, et les effets de consanguinité et stochasticité[217]. L'analyse du génome d'un individu néandertalien tardif, surnommé « Thorin », trouvé dans la grotte Mandrin (Drôme) semble confirmer ces hypothèses : celui-ci révèle une profonde divergence avec les autres Néandertaliens tardifs, Thorin appartenant à une population de petite taille qui ne présentait aucune introgression génétique avec d'autres Néandertaliens européens tardifs connus, révélant l'isolement génétique de sa lignée bien que ces individus vécussent dans des régions voisines[218].
Coexistence avec Homo sapiens
[modifier | modifier le code]La disparition des Néandertaliens serait liée à l'arrivée des hommes modernes et à la compétition territoriale pour l'exploitation des ressources[219], voire à leur élimination physique par les hommes modernes à l'occasion de conflits violents sur les zones de contact. L'hypothèse s'appuie en particulier sur la concomitance de l'expansion d'Homo sapiens et de la disparition de Néandertal[220] ; les Homo sapiens d'Europe de l'Ouest cohabitant par exemple pendant plusieurs milliers d'années (500 à 5 400 ans suivant les régions) avec les Néandertaliens[209]. Elle se heurte toutefois à l'absence de traces de morts violentes ou de traces de cohabitation prolongée sur un même territoire[221]. En outre on peut objecter que les deux groupes ne devaient pas occuper l'ensemble du territoire européen et que les Néandertaliens avaient une meilleure connaissance de ce territoire et de ses ressources que les nouveaux arrivants[222]. La domestication du chien pour la chasse correspondant approximativement à la période de disparition des Néandertaliens, Pat Shipman a émis l'hypothèse selon laquelle cet événement aurait permis à Homo sapiens de bénéficier d'un net avantage dans sa recherche de nourriture[223],[224].
Outre cette hypothèse de l'extinction violente liée à l'homme moderne, les Néandertaliens auraient pu succomber à une maladie mortelle (épidémies) apportée par les Sapiens originaires d'Afrique[225].
La trompe d'Eustache néandertalienne, plus courte que celle des Homo sapiens, aurait favorisé des infections de l'oreille à répétition, fragilisant les Néandertaliens et participant à leur disparition[226].
L'Homme de Néandertal dans les œuvres de fiction
[modifier | modifier le code]
Les Néandertaliens ont été dépeints dans la culture populaire, y compris les apparitions dans la littérature, les médias visuels et la comédie. L'archétype des « hommes des cavernes » se moque souvent des Néandertaliens et les décrit comme des personnages primitifs, bossus, brandissant des massues, grognants, des personnages antisociaux mus uniquement par l'instinct animal. « Néandertal » peut également être utilisé comme une insulte[227].
Littérature
[modifier | modifier le code]- The Inheritors, 1955, de William Golding : chef-d'œuvre de l'auteur de Lord of the flies, ce récit met en scène la rencontre par un petit groupe de Néandertaliens d'un groupe de Cro-Magnons et son issue fatale.
- Philip José Farmer, dans son cycle de romans Le Fleuve de l'éternité, 1971-1983, met en scène un Néandertalien du nom de Kazz, comme personnage de second plan.
- The clan of the Cave Bear, 1980, de Jean M. Auel, traduit en français, d'abord sous le titre Ayla, l'enfant de la Terre, puis plus tard, sous le titre Le Clan de l'Ours des Cavernes : les aventures d’Ayla, une fillette de Cro-Magnon orpheline recueillie vers l'âge de cinq ans par une tribu de Néandertaliens.
- L'Affreux Petit Garçon, 1992 d'Isaac Asimov & Robert Silverberg : histoire d'un enfant néandertalien, ramené à l'époque contemporaine pour être étudié. Un conte scientifique et philosophique.
- Neanderthal: their time has come, 1996 de John Darnton : récit d'une rencontre entre des hommes modernes et des Néandertaliens ayant survécu jusqu'à nos jours.
- Hominids: the Neanderthal parallax, 2002 de Robert J. Sawyer : récit d'un monde imaginaire où les rôles d'Homo sapiens et de l'homme de Néandertal sont inversés.
- L'Odyssée du dernier Neandertal, 2003-2010, de Marc Klapczynski, trilogie comprenant trois romans : Aô, l'homme ancien, Le pouvoir d'Iktia et Tsinaka, l'œil de la toundra publiés aux éditions Aubéron.
- La série Thursday Next de Jasper Fforde, série science-fictive sur une Terre alternative, où les hommes de Neandertal ont été clonés.
- Neandertal, 2007-2001, de Emmanuel Roudier, série de 3 albums de BD aux éditions Delcourt.
- Au crépuscule de Néandertal, 2014, de Olivier Merle aux Éditions de Fallois.
Cinéma
[modifier | modifier le code]- The Neanderthal Man, film de Ewald André Dupont, 1953.
- La Guerre du feu, film de Jean-Jacques Annaud, 1981, d'après le roman La Guerre du feu de J.-H. Rosny aîné (1909).
- Iceman, film de science-fiction avec Timothy Hutton, Danny Glover et John Lone, 1984.
- Neandertal, film de Ingo Haeb, Jan Glaser, Jan-Christoph Glaser, 2006.
- Humains, film avec Lorànt Deutsch, sorti en France le 22 avril 2009.
- Ao, le dernier Néandertal, film de Jacques Malaterre, sorti en France le 29 septembre 2010.
- La Nuit au musée (série de films) : un groupe de Néandertaliens apparaît à plusieurs reprises dans les films.
Télévision
[modifier | modifier le code]- Korg: 70,000 B.C. (1974-1975)
Notes et références
[modifier | modifier le code]Notes
[modifier | modifier le code]- Il s'agit du fragment osseux dénommé Denisova-11. L'adolescente a été surnommée Denny.
- Le début de la migration d'Homo sapiens il y a environ 48 000 ans pourrait avoir été favorisée par un épisode climatique tempéré de la dernière glaciation.
Références
[modifier | modifier le code]- (en) Matthias Meyer, Svante Pääbo et al., « Nuclear DNA sequences from the Middle Pleistocene Sima de los Huesos hominins », Nature, 7595e série, vol. 531, , p. 504-507 (DOI 10.1038/nature17405, lire en ligne)
- (en) Christopher Brian Stringer, « The origin and evolution of Homo sapiens », Philosophical transactions of the Royal Society, (DOI 10.1098/rstb.2015.0237, lire en ligne)
- Néandertal vivait en Espagne il y a 430 000 ans, mediapart.fr, 15 mars 2016
- (en) Juan Luis Arsuaga et al., « Neandertal roots: Cranial and chronological evidence from Sima de los Huesos », Science, vol. 344, no 6190, , p. 1358-1363 (DOI 10.1126/science.1253958, lire en ligne)
- « Le changement climatique pourrait avoir aidé Homo sapiens à remplacer Néandertal en Europe », sur Maxisciences, (consulté le )
- (en) « Here's What Happened When Neanderthals And Ancient Humans Hooked Up 80000 Years Ago » (consulté le )
- (en) Emily Singer, « How Neanderthal DNA Helps Humanity », quantamagazine.org, (lire en ligne).
- Marc Groenen, Pour une histoire de la préhistoire, Éditions Jérôme Millon, , p. 485.
- (en) Alex Bridget, « Is It Neander-TAL or Neander-THAL? », sur blogs.discovermagazine.com, .
- C. J. Fuhlrott, « Menschliche Ueberreste aus einer Felsengrotte des Düsselthals. Ein Beitrag zur Frage über die Existenz fossiler Menschen » sur Google Livres, Verhandl. Naturhist. Ver. Preuss. Rheinlande Westphalen, 16, 1859, p. 131–153
- Fuhlrott, C.J. (1865) - Der fossile Mensch aus dem Neanderthal und sein Verhältniß zum Alter des Menschengeschlechts, Duisbourg, 78 S. 2 Abb.
- H. Schaaffhausen, « Zur Kentniss der ältesten Rassenschädel » sur Google Livres, dans Johannes Müller (dir.), Archiv fur Anatomie, Physiologie und wissenschaftliche Medicin, in Verbindung mit mehreren Gelehrten herausgegeben, 1858, p. 453–488
- André Leroi-Gourhan, « La grotte du Loup, Arcy-sur-Cure », Bulletin de la Société préhistorique française, vol. 47, no 5, , p. 268-280 (lire en ligne, consulté le ), p. 278, note 12.
- (en) W. King, 1864 - « The reputed fossil man of the Neanderthal », Quarterly Journal of Science, vol. 1, p. 88-97.
- M. Boule, 1911-1913 - « L’homme fossile de la Chapelle-aux-Saints », dans Annales de paléontologie, t. VI-VII-VIII.
- Fanny Defrance-Jublot, « Quand des abbés lèvent le doute sur les néandertaliens », Pour la Science, no 33, (résumé)
- Antoine Lesaint, « La semaine cléricale », La Lanterne, (lire en ligne)
- Jean-Claude Blanchet, « Néandertal Un problème humain et scientifique » (consulté le )
- Émile Gautier, « Tablettes du progrès : L'homme descend-il du singe ? », La Lanterne, (lire en ligne)
- (en) Richard E. Green et al., « A draft sequence of the Neandertal genome », Science, vol. 328, no 5979, , p. 710-722 (lire en ligne)
- (en) Mateja Hajdinjak, Qiaomei Fu et al., « Reconstructing the genetic history of late Neanderthals », Nature, vol. 555, , p. 652-656 (DOI 10.1038/nature26151).
- (en) Kay Prüfer et al., A high-coverage Neandertal genome from Vindija Cave in Croatia, Science Vol. 358, Issue 6363, p. 655-658, 2017 DOI 10.1126/science.aao1887
- « Néandertal, un grand bavard de la préhistoire ? », sur hominides.com, (consulté le )
- (en) María Martinón-Torres, José María Bermúdez de Castro, Aida Gómez-Robles, Leyre Prado-Simón et Juan Luis Arsuaga, « Morphological description and comparison of the dental remains from Atapuerca-Sima de los Huesos site (Spain) », Journal of Human Evolution, 1re série, vol. 62, , p. 7-58 (DOI 10.1016/j.jhevol.2011.08.007, lire en ligne)
- Marina Martínez de Pinillos, María Martinón-Torres, Matthew M.Skinner, Juan Luis Arsuaga, Ana Gracia-Téllez, Ignacio Martínez, Laura Martín-Francésa et José María Bermúdez de Castro, « Expression des crêtes du trigonide de molaires inférieures à Atapuerca-Sima de los Huesos : expression morphologique interne et externe et inférences évolutionnistes », Comptes-rendus Palévol, 3e série, vol. 13, , p. 205-221 (DOI 10.1016/j.crpv.2013.10.008, lire en ligne)
- (en) Joan Daura, Montserrat Sanz, Juan Luis Arsuaga, Dirk L. Hoffmann, Rolf M. Quam, María Cruz Ortega, Elena Santos, Sandra Gómez, Angel Rubio, Lucía Villaescusa, Pedro Souto, João Mauricio, Filipa Rodrigues, Artur Ferreira, Paulo Godinho, Erik Trinkaus et João Zilhão, « New Middle Pleistocene hominin cranium from Gruta da Aroeira (Portugal) », Proceedings of the National Academy of Sciences of the United States of America, 13e série, vol. 114, , p. 3397-3402 (DOI 10.1073/pnas.1619040114, lire en ligne)
- (en) Jean-Jacques Hublin, « The origin of Neandertals », Proceedings of the National Academy of Sciences of the United States of America, 38e série, vol. 106, , p. 16022-16027 (DOI 10.1073/pnas.0904119106, lire en ligne)
- (en) Ohta Shoji, Nishiaki Yoshihiro, Abe Yoshito, Mizoguchi Yuji, Kondo Osamu, Dodo Yukio, Muhesen Sultan, Akazawa Takeru, Oguchi Takashi et Haydal Jamal, « Neanderthal infant burial from the Dederiyeh cave in Syria », Paléorient, vol. 21, no 2, , p. 77-86 (DOI 10.3406/paleo.1995.4619, lire en ligne).
- (en) Krause, J., Orlando, L., Serre, D., Viola, B., Prufer, K., Richards, M. P., Hublin, J.-J., Hanni, C., Derevianko, A. P. et Paabo, S. (2007) - « Neanderthals in central Asia and Siberia », Nature, 449, 7164, p. 902-904. (résumé)
- « Des néandertaliens complètement à l'est ! », sur hominidés.com.
- (en) Jean-Pierre Bocquet-Appel et Anna Degioanni, « Neanderthal Demographic Estimates », Current Anthropology, vol. 54, no 8, , p. 202-213 (DOI 10.1086/673725)
- « À la rencontre de Néandertal - Regarder le documentaire complet », sur ARTE (consulté le )
- Jean-Jacques Hublin, « Origine et évolution des Néandertaliens », dans Yves Coppens et Pascal Picq (dir.), Aux origines de l'humanité, vol. 1, Fayard, .
- (en) Jean-Philippe Faivre, Bruno Maureille, Priscilla Bayle, Isabelle Crèvecœur, Matthieu Duval et al., « Middle Pleistocene Human Remains from Tourville-la-Rivière (Normandy, France) and Their Archaeological Context », PLoS one, vol. 9, no 10, , e104111 (DOI 10.1371/journal.pone.0104111).
- Nathaniel Herzberg, « Le bras d’un pré-Néandertalien trouvé au bord de la Seine », Le Monde, (lire en ligne)
- Demoule, J.-P., « « De l’archéologie de sauvetage à l’archéologie préventive »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?) », préface de La France archéologique, éditions Hazan/Inrap.
- Jean-Louis Heim, Les Hommes fossiles de La Ferrassie, Paris, Masson, 1976.
- Hublin J.J., Barroso Ruiz C., Medina Lara P., Fontugne M., Reyss J.-L., 1995 - The Mousterian site of Zafarraya (Andalucia, Spain): dating and implications on the palaeolithic peopling processes of Western Europe. C. R. Acad. Sc. Paris. 321 (IIa): 931-937.
- Hublin J.-J et E.Trinkaus, 1998 - The Mousterian human remains from Zafarraya (Andalucia, Spain). American Journal of Physical Anthropology suppl. 26 : 122-123.
- Smith, F.H., Trinkaus, E. Pettitt, P.B., Karavanic', I. et Paunovic, M., (1999) « Direct radiocarbon dates for Vindija G1 and Velika Pećina Late Pleistocene hominid remains », PNAS, Vol. 96, numéro 22, 12281-12286, 26 octobre 1999.
- Higham, T., Ramsey, C.B., Karavanic, I., Smith, F.H. et Trinkaus, E., (2006), « Revised direct radiocarbon dating of the Vindija G1 Upper Paleolithic Neandertals », PNAS, 17 janvier 2006, vol. 103, no. 3, p. 553-557.
- Higham, T., Brock, F., Peresani, M., Broglio, A., Wood, R. et Douka, K. (2009) - « Problems with radiocarbon dating the Middle to Upper Palaeolithic transition in Italy », Quaternary Science Reviews, 28, p. 1257-1267.
- Maroto, J., Vaquero, M., Arrizabalaga, Á., Baena, J., Baquedano, E., Jordá, J., Julià, R., Montes, R., van der Plicht, J., Rasines, P. et Wood, R. (2012) - « Current issues in late Middle Palaeolithic chronology: New assessments from Northern Iberia », Quaternary International, 247, p. 15-25.
- Finlayson, C. et al. (2006) « Late survival of Neanderthals at the southernmost extreme of Europe »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?), Nature, advanced online publication, 13 septembre 2006.
- Brill, D. (2006) « Neanderthal's last stand », news@nature.com, 13 septembre 2006.
- Zilhão, J. et Pettitt, P., (2006), « On the New Dates for Gorham's Cave and the Late Survival of Iberian Neanderthals »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?), Before Farming, 2006/3, article 3, p. 95-122
- (en) Ludovic Slimak, John Inge Svendsen, Jan Mangerud, Hugues Plisson, Herbjørn Presthus Heggen, Alexis Brugère, Pavel Yurievich Pavlov, « Late Mousterian Persistence near the Arctic Circle », Science, vol. 332, no 6031 p. 841-845 du 13 mai 2011.
- L’Homme de Neandertal, près du cercle polaire ? Le site russe de Byzovaya émission Le Salon noir du 22 juin 2011 sur France Culture.
- Zwyns, N., Roebroeks, W., McPherron, S.P., Jagich, A. et Hublin, J.-J. (2012) - « Comment on “Late Mousterian Persistence near the Arctic Circle” », Science, vol. 335, no 6065, p. 167.
- Bernard Vandermeersch et Maureille, B. (2007) - Les Néandertaliens, biologie et cultures, Paris, Éditions du Comité des travaux historiques et scientifiques, Documents préhistoriques 23, 342 p.
- (en) Trenton W. Holliday, « Postcranial evidence of cold adaptation in European Neandertals », American Journal of Physical Anthopology, vol. 104, no 2, , p. 245–258.
- « Le cerveau humain depuis le paléolithique », sur hominides.com (consulté le )
- (en) Stephen Wroe, William C. H. Parr, Justin A. Ledogar, Jason Bourke, Samuel P. Evans, Luca Fiorenza, Stefano Benazzi, Jean-Jacques Hublin, Chris Stringer, Ottmar Kullmer, Michael Curry, Todd C. Rae, Todd R. Yokley, « Computer simulations show that Neanderthal facial morphology represents adaptation to cold and high energy demands, but not heavy biting », Proceedings of the Royal Society B, Royal Society, vol. 285, no 1876, (ISSN 0962-8452, PMID 29618551, DOI 10.1098/rspb.2018.0085).
- Rees, J. L. (2000) - « The Melanocortin 1 Receptor (MC1R): more than just red hair », Pigment Cell Res, 13, p. 135–140.
- Harding, R.M., Healy, E., Ray, A.J., Ellis, N.S., Flanagan, N., Todd, C., Dixon, C., Sajantila, A., Jackson, I.J., Birch-Machin, M.A. et Rees, J.L. (2000) - « Evidence for variable selective pressures at MC1R », American Journal of Human Genetics, 66, 4, p. 1351-1361.
- « Red-Heads and Neanderthals »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?)
- « Red hair genes 100,000 years old »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?).
- Lalueza-Fox, C., Römpler, H., Caramelli, D., Stäubert, C., Catalano, G., Hughes, D., Rohland, N., Pilli, E., Longo, L., Condemi, S., de la Rasilla, M., Fortea, J., Rosas, A., Stoneking, M., Schöneberg, T., Bertranpetit, J. et Hofreiter, M. (2007) - « A melanocortin 1 receptor allele suggests varying pigmentation among Neanderthals », Science, mise en ligne 25 octobre 2007. résumé.
- « Certains hommes de Néandertal avaient les cheveux roux, selon leur ADN »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?).
- Paul Rincon, « Neanderthals 'were flame-haired' », BBC News.uk, (lire en ligne)
- Stéphane Foucart, « L'homme de Neandertal, ce rouquin au teint pâle », Le Monde, (lire en ligne)
- (en) Caio C S Cerqueira, Vanessa R Paixão-Côrtes, Francis M B Zambra, Francisco M Salzano, Tábita Hünemeier et Maria-Cátira Bortolini, « Predicting homo pigmentation phenotype through genomic data : From neanderthal to James Watson », American Journal of Human Biology, vol. 24, no 5, , p. 705-709 (DOI 10.1002/ajhb.22263)
- Trust My Science, « ADN des Néandertaliens : forte consanguinité et isolement génétique depuis 50 000 ans », sur trustmyscience.com, (consulté le )
- Tropel, Ch. (2004) « Approche synthétique de la pathologie des Néanderthaliens », Paléobios, 13. « texte intégral »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?).
- Bar Yosef O. et Bernard Vandermeersch (1991) Le squelette moustérien de Kébara 2, Cahiers de Paléoanthropologie, CNRS, 197 p.
- Trinkaus, E. (1983) The Shanidar Neandertals, New York : Academic Press, 502 p.
- Montet-White, A. (1996) Le Paléolithique en ancienne Yougoslavie, Jérôme Millon, 268 p.
- Tappen N.C. (1985) « The dentition of the « Old man » of la Chapelle-aux-Saints and inferences concerning Neandertal behavior », Am. J. Phys. Anthropol., 67 : p. 43-50.
- Trinkaus E. (1985) « Pathology and the posture of the la Chapelle-aux-Saints Neandertal », Am. J. Phys Anthropol., 67 : p. 19-41.
- Ogilvie M.D., Curvan B.K. et Trinkaus E. (1989) « Incidence and patterning of dental enamel hypoplasia among the Neandertals », Am. J. Phys. Anthropol., 79 : p. 25-41.
- L'individu aurait vécu jusqu'à l'âge de 50 à 60 ans. « Homme de La Chapelle-aux-Saints - Homo neanderthalensis », sur hominides.com.
- Arensburg, B. et Tillier, A.-M. (1990) « Le langage des néandertaliens », La Recherche, vol. 21, no 224, p. 1084-1086.
- Heim, J.-L, Boë, L.J. et Abry, Ch. (2002) « La parole à la portée du conduit vocal de l'homme de Neandertal. Nouvelles recherches, nouvelles perspectives », Comptes Rendus Palevol, Volume 1, Issue 2, p. 129-134
- (en) B. Arensburg, « The vertebral column, thoracic cage and hyoid bone », dans Ofer Bar-Yosef et Bernard Vandermeersch (éds.), Le squelette moustérien de Kébara 2, Paris, France, Éditions du CNRS, , p. 113-146.
- (en) B. Arensburg, A. M. Tillier, B. Vandermeersch, H. Duday, L. A. Schepartz et Y. Rak, « A Middle Palaeolithic human hyoid bone », Nature, vol. 338, , p. 758-760 (DOI 10.1038/338758a0).
- (es) L. Rodríguez, L.L. Cabo et J.E. Egocheaga, « Breve nota sobre el hioides Neandertalense de Sidrón (Piloña, Asturias) », dans M. P. Aluja, A. Malgosa et R. M. Nogués (éds.), Antropología y Diversidad, vol. 1, Barcelone, Espagne, Edicions Bellaterra, , p. 484-493.
- (en) I. Martinez, L. Arsuaga, R. Quam, J. M. Carretero, A. Gracia et L. Rodrìguez, « Human hyoid bones from the middle Pleistocene site of the Sima de los Huesos (Sierra de Atapuerca, Spain) », Journal of Human Evolution, vol. 54, no 1, , p. 118-124 (DOI 10.1016/j.jhevol.2007.07.006).
- Jean-Paul Demoule, Naissance de la figure : L'art du paléolithique à l'âge du fer, Paris, Gallimard, coll. « Folio Histoire », , 308 p. (ISBN 9782072696756), p. 28.
- Boë, L.-J., Heim, J.-L., Honda, K. Maeda, S. (2002) « The potential of Neandertal vowel space was as large as that of modern humans », Journal of Phonetics, 30, 465-484.
- Lieberman, P. (2007) « Current views on Neanderthal speech capabilities: A reply to Boë et al. (2002) », Journal of Phonetics, 35, 552-563.
- Boë, L.J., Heim, J.L., Honda, K., Maeda, S., Badin, P., Abry, C. (2007) « The vocal tract of newborn humans and Neanderthals: Acoustic capabilities and consequences for the debate on the origin of language. A reply to Lieberman », Journal of Phonetics, 35, 564–581.
- Krause, J., Lalueza-Fox, C., Orlando, L., Enard, W., Green, R.E., Burbano, H.A., Hublin, J.-J., Hänni, C., Fortea, J., de la Rasilla, M., Bertranpetit, J., Rosas, A. et Pääbo, S. (2007) - « The derived FOXP2 variant of modern humans was shared with Neandertals »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?), Current Biology, vol. 17, 21, p. 1908-1912.
- Néandertal, un grand bavard de la Préhistoire ?
- Camille Gaubert, « Néandertal aurait perçu le son et la voix aussi bien que l'humain », Sciences et Avenir, (lire en ligne, consulté le ).
- Kate Wong, « Le crépuscule de l'homme de Néandertal », Pour La Science, no 386, décembre 2009.
- Baffier D., Les derniers Néandertaliens, le Châtelperronien, La Maison des roches, Paris, 1999.
- Pelegrin J., Technologie lithique : Le Châtelperronien de Roc-de-Combe (Lot) et de La Côte (Dordogne), Cahiers du Quaternaire, CNRS, Paris, 1995.
- « Une industrie passée inaperçue : quand Néandertal transforme l’os en outil », sur CNRS, (consulté le ).
- (en) Malvina Baumann, Hugues Plisson, Serge Maury, Sylvain Renou, Hélène Coqueugniot et al., « On the Quina side: A Neanderthal bone industry at Chez-Pinaud site, France », PLOS One, (DOI 10.1371/journal.pone.0284081
).
- (en) Éric Boëda, Stéphanie Bonilauri, Jacques Connan, Dan Jarvie, Norbert Mercier, Mark Tobey, Hélène Valladas and Heba al Sakhel, « New Evidence for Significant Use of Bitumen in Middle Palaeolithic Technical Systems at Umm el Tlel (Syria) around 70,000 BP », Paléorient, vol. 34, no 2, , p. 67-83.
- J. Jaubert et A. Delagnes, « De l'espace parcouru à l'espace habité au Paléolithique moyen », dans Bernard Vandermeersch et B. Maureille, Les Néandertaliens. Biologie et cultures, Éditions du CTHS, , p. 263-281.
- Gérard Cordier, « Blessures préhistoriques animales et humaines avec armes ou projectiles conservés », Bulletin de la Société préhistorique française, t. 87, nos 10-12, , p. 462-482 (DOI 10.3406/bspf.1990.9929, lire en ligne).
- (en) Andrew Curry, « Rare wooden artifacts showcase the smarts of early Neanderthals », Science, vol. 384, no 6691, (DOI 10.1126/science.z5hff6o).
- Lewis Binford, Bones, Ancient Men and Modern Myths, Academic Press, 1981.
- (de) Hartmut Thieme, Ein angekohlter Holzstab vom altpalàolithischen Fundplatz Schôningen (Un bâton de bois carbonisé du site paléolithique ancien de Schôningen), In Den Bogen spannen: Festschrift fiir Bernhardt Gramsch, Beier & Beran, Archaeologische Fachliteratur, (ISBN 9783930036356), volume 1, p. 15-27
- Silvana Condemi et François Savatier, Néandertal, mon frère, Flammarion, (lire en ligne), p. 74
- (en) Hervé Bocherens, « Isotopic biogeochemistry as a marker of Neandertal diet » [« La biogéochimie isotopique comme marqueur de l'alimentation des Néandertaliens »], Anthropologischer Anzeiger, vol. 55, no 2, , p. 101-120.
- Cyril Bousquet, « Une spécialiste de Néandertal « pessimiste sur notre avenir » », sur nouvelobs.com, .
- Brugal, J.-Ph. et Jaubert, J. (1991) - « Les gisements paléontologiques pléistocènes à indices de fréquentation humaine : un nouveau type de comportement de prédation ? », Paléo, no 3, p. 15-41.
- Science & Vie junior, Chasseurs ou charognards ?, Hors série, no 22, octobre 1995, p. 68–72.
- (en) Carles Lalueza-Fox et al., « Genetic evidence for patrilocal mating behavior among Neandertal groups », PNAS, vol. 108, no 1, , p. 250-253 (DOI 10.1073/pnas.1011553108, lire en ligne).
- Marylène Patou-Mathis, Préhistoire de la violence et de la guerre, Odile Jacob, , p. 57.
- (en) Dick Stapert, « Neanderthal children and their flints », PalArch’s Journal of Archaeology of Northwest Europe, vol. 1, no 2, , p. 1-25 (lire en ligne, consulté le ), p. 6.
- (en) « Learning by Heart: Cultural Patterns in the Faunal Processing Sequence during the Middle Pleistocene », PLOS-one, (lire en ligne, consulté le ).
- (en) Fabrizio Mafessoni et al., « A high-coverage Neandertal genome from Chagyrskaya Cave », biorxiv.org, (consulté le ).
- (en) Andrew Curry, « Neanderthals lived in groups big enough to eat giant elephants », Science, vol. 379, no 6631, (DOI 10.1126/science.adg9641
- (en) Sabine Gaudzinski-Windheuser, Lutz Kindler, Katharine MacDonald et Wil Roebroeks, « Hunting and processing of straight-tusked elephants 125.000 years ago: Implications for Neanderthal behavior », Science Advances, vol. 9, no 5, (DOI 10.1126/sciadv.add8186
- (en) Peter Ungar, Evolution of the human diet : the known, the unknown, and the unknowable, New York, Oxford University Press, Inc., , 413 p., relié (ISBN 978-0-19-518346-7 et 0-19-518346-0, LCCN 2005036120, lire en ligne).
- (en) Amanda G. Henry et coll, « Microfossils in calculus demonstrate consumption of plants and cooked foods in Neanderthal diets (Shanidar III, Iraq; Spy I and II, Belgium) », Proceedings of the National Academy of Sciences, (lire en ligne).
- Agusti, B., Alcalde, G., Güell, A., Juan-Muns, N., Nebot, J, Rueda, J.M. et Terradas, X. (1992) - « Caracoles terrestres en la cueva 120 - Aportacion al conocimiento de la dieta alimenticia durante el Paleolitico medio », Revista de Arqueologia, Ano XII, no 131, p. 8-11.
- (en) El Zaatari, S., Grine, F.E., Ungar, P.S., Hublin, J.-J., Ecogeographic variation in Neandertal dietary habits: « Evidence from occlusal molar microwear texture analysis », Journal of Human Evolution, 61(4), 2011, p. 411-424
- (en) Wißing, C., Rougier, H., Crevecoeur, I., Germonpré, M., Naito Y.I., Semal, P., Bocherens, H., « Isotopic evidence for dietary ecology of Neanderthals in North-Western Europe », Quaternary International, Vol. 411, 8 août 2016, p. 327-345
- Cortés-Sánchez, Miguel, Morales-Muñiz, Arturo, Simón-Vallejo, M.D., Lozano-Francisco, M.C., Vera-Peláez, J.L., Finlayson, C., Rodríguez-Vidal, J., Delgado-Huertas, A., Jiménez-Espejo, F.J., Martínez-Ruiz, F., Martínez-Aguirre, M.A., Pascual-Granged, A.J., Bergadà -Zapata, M.M., Gibaja-Bao, J.F., Riquelme-Cantal, J.A., López-Sáez, J.A., Rodrigo-Gámiz, M., Sakai, S., Sugisaki, S., Finlayson, G., Fa, D.A. et Bicho, N.F. (2011) - « Earliest known use of marine resources by Neanderthals », PLoS ONE, 6, 9, p. e24026.
- avec AFP, « Les néandertaliens mangeaient des fruits de mer il y a 150 000 ans », Le Monde.fr, (lire en ligne)
- (en) Sid Perkins (2017) Neandertals ate woolly rhinos and mushrooms, may have used painkillers and antibiotics, Science magazine, 8 mars 2017.
- (en) Donald G. Barceloux, Medical Toxicology of Natural Substances : Foods, Fungi, Medicinal Herbs, Plants, and Venomous Animals, John Wiley & Sons, , p. 421.
- (en) Maxime Pelletier, Emmanuel Desclaux, Jean-Philip Brugal et Pierre-Jean-Texier, « The exploitation of rabbits for food and pelts by last interglacial Neandertals », Quaternary Science Reviews (en), vol. 224, , article no 105972 (DOI 10.1016/j.quascirev.2019.105972).
- C.B. Stringer, J.C. Finlayson, R.N.E. Bartond, Y. Fernández-Jalvo, I. Cáceres, R.C. Sabin, E.J. Rhodes, A.P. Currant, J. Rodríguez-Vidal, F. Giles-Pacheco, et J.A. Riquelme-Cantal, « Neanderthal exploitation of marine mammals in Gibraltar », PNAS, vol. 105, no 38, , p. 14319-14324 (lire en ligne)
- Neanderthal diet like early modern human's Preuve de chasse maritime & composition du régime de Néandertal, sur le site du Natural History Museum, vidéo & texte.
- Hominidés
- Géo
- Bruno Maureille, Les premières sépultures, Le Pommier / Cité des sciences et de l'industrie, .
- Arlette Leroi-Gourhan, « Le Néanderthalien IV de Shanidar », Bulletin de la Société préhistorique française, vol. 65, no 3, , p. 79-83 (lire en ligne, consulté le ).
- (en) Arlette Leroi-Gourhan, « The Flowers Found with Shanidar IV, a Neanderthal Burial in Iraq », Science, vol. 190, no 4214, , p. 562-564 (DOI 10.1126/science.190.4214.562, lire en ligne, consulté le ).
- Arlette Leroi-Gourhan, « Shanidar et ses fleurs », Paléorient, vol. 24, no 2, , p. 79-88 (lire en ligne, consulté le ).
- (en) Ralph S. Solecki, Shanidar, the first flower people, New York, A. Knopf, .
- (en) D. J. Sommer, « The Shanidar IV 'Flower Burial' : a Re-evaluation of Neanderthal Burial Ritual », Cambridge Archaeological Journal, vol. 9, no 1, , p. 127-129 (lire en ligne, consulté le ).
- Florian Bardou, « En Dordogne, et par-delà, l’homme de Néandertal enterrait bien ses morts », sur liberation.fr, (consulté le ).
- Ofer Bar-Yosef, Henri Laville, Liliane Meignen, Anne-Marie Tillier, Bernard Vandermeersch, Baruch Arensburg, Anna Belfer-Cohen, Paul Goldberg, Yoel Rak et Eitan Tchernov, « La Sépulture Néandertalienne de Kébara (Unité XII) », dans Marcel Otte, L'Homme de Neandertal, vol. 5 : La Pensée (Actes du colloque international de Liège, 4-7 décembre 1986), Études et Recherches Archéologiques de l'Université de Liège, , 17-24 p. (lire en ligne).
- Maria Dolores Garralda et Bernard Vandermeersch, « Les Néandertaliens de la grotte de Combe-Grenal (Domme, France) », Paléo, no 12, , p. 213-259 (lire en ligne).
- François Savatier, « Des Néandertaliens cannibales dans la vallée du Rhône », sur pourlascience.fr, Pour la science, (consulté le ).
- (en) Alban R. Defleur et Emmanuel Desclaux, « Impact of the last interglacial climate change on ecosystems and Neanderthals behavior at Baume Moula-Guercy, Ardèche, France », Journal of Archaeological Science, vol. 104, , p. 114-124 (DOI 10.1016/j.jas.2019.01.002, lire en ligne).
- Bernard Vandermeersch et Bruno Maureille, Les Néandertaliens, biologies et cultures, chap. « Les sépultures néandertaliennes », éd. Comité des travaux historiques et scientifiques, 2007, 342 p.
- Bonifay, E. (1965) - « Un ensemble rituel moustérien à la grotte du Regourdou (Montignac, Dordogne) », in: Actes du IVe Congrès de l'UISPP, Rome, vol. II, p. 136-140.
- Bonifay, E. (2002) - « L'Homme de Neandertal et l'ours (Ursus arctos) dans la grotte du Regourdou (Montignac-sur-Vézère, Dordogne, France) », in: L'ours et l'Homme, Tillet, T. et Binford, L., (Éds.), Liège, ERAUL 100, p. 247-254.
- Cavanhié, N. (2007) - Étude archéozoologique et taphonomique des grands carnivores du site paléolithique moyen de Régourdou (Montignac, Dordogne), Université de Toulouse II le Mirail, Mémoire de Master 2.
- « Le Regourdou aujourd’hui… », sur pole-prehistoire.com.
- A. Defleur, T. White, P. Valensi, L. Slimak et E. Crégut-Bonnoure, « Neanderthal cannibalism at Moula-Guercy, Ardèche, France » [« Cannibalisme néandertalien à Moula-Guercy, Ardèche, France »], Science, vol. 286, , p. 128-131 (résumé).
- Gorjanoviv-Kramberger, D. (1906), Der diluviale Mensh von Krapina in Kroatia, Ein Betrag zur Paläoanthropologie, Kreidel, Wiesbaden.
- Fred Hines Smith, The Neandertal remains from Krapina. A descriptive and comparative study [« Les vestiges néa,dertaliens de Kapina. Une étude descriptive et comparative »] (thèse d'anthropologie), Knoxville, University of Tennessee, , 359 p. (présentation en ligne).
- T.D. White, Prehistoric cannibalism at Mancos [« Cannibalisme préhistorique à Mancos 5MTUMR-2346 »], Princeton University Press, , 462 p. (présentation en ligne).
- T.D. White et N. Toth, « The Question of Ritual Cannibalism at Grotta Guattari », Current Anthropology, vol. 32, no 2, , p. 118-138.
- cf. également les références de l'article « Grotte Guattari ».
- Catherine Girard, Les Industries moustériennes de la grotte de l'Hyène à Arcy-sur-Cure, Yonne, Paris, CNRS, , 224 p. (lire en ligne), p. 27.
- Silvana Condemi et François Savatier, Néandertal, mon frère, Flammarion, (lire en ligne), p. 98.
- S. Révillion et A. Tuffreau, Les industries laminaires du Paléolithique moyen, Éditions du CNRS, , 193 p. (présentation en ligne).
- M. Soressi et F. d’Errico, « Pigments, gravures, parures : les comportements symboliques controversés des Néandertaliens », dans Bernard Vandermeersch et Bruno Maureille, Les Néandertaliens, biologie et culture, CTHS, (lire en ligne), p. 297-309.
- (en) M. Peresani, I. Fiore, M. Gala, M. Romandini et A. Tagliacozzo, « Late Neandertals and the intentional removal of feathers as evidenced from bird bone taphonomy at Fumane Cave 44 ky B.P., Italy », Proceedings of the National Academy of Sciences, vol. 108, no 10, , p. 3888-3893 (lire en ligne, consulté le ).
- Lorblanchet, M. (1999) La naissance de l'Art. Genèse de l'art préhistorique, Paris, Éd. Errance, 304 p.
- Jacques Jaubert, Chasseurs et artisans du Moustérien, Paris, La Maison des Roches, coll. « Histoire de la France préhistorique », , 152 p. (ISBN 2912691052)
- Joaquín Rodríguez-Vidal, Francesco d'Errico, Francisco Giles Pacheco et al., « A rock engraving made by Neanderthals in Gibraltar », Proceedings of the National Academy of Sciences, (résumé).
- « Découverte de la première gravure néandertalienne dans une grotte à Gibraltar », sur culturebox.francetvinfo.fr, (consulté le ).
- (en) Tim Appenzeller, « Europe's first artists were Neandertals », Science, vol. 359, no 6378, , p. 852-853 (DOI 10.1126/science.359.6378.852, lire en ligne).
- (en) D. L. Hoffmann, C. D. Standish, M. García-Diez, P. B. Pettitt, J. A. Milton et al., « U-Th dating of carbonate crusts reveals Neandertal origin of Iberian cave art », Science, vol. 359, no 6378, , p. 912-915 (DOI 10.1126/science.aap7778, lire en ligne, consulté le ).
- (en) Paul Rincon, « Neanderthals were capable of making art », BBC News, (lire en ligne, consulté le ).
- (en) Ian Sample, « Neanderthals – not modern humans – were first artists on Earth, experts claim », sur The Guardian, (consulté le )
- Ludovic Slimak, Jan Fietzke, Jean-Michel Geneste, Roberto Otanon, « Néandertal, l'homme qui oublia de peindre les grottes ? », Archéologia, n°570, novembre 2018, pp.4-5
- Ludovic Slimak, Jan Fietzke, Jean-Michel Geneste, Roberto Otanon, « Comment on U-th dating of carbonate crusts reveal Neandertal origin of Iberian cave art », Science, n° 361, doi.org/10.1126/science.aau1371
- Hélène Salomon et al., « Les matières colorantes au début du Paléolithique supérieur : caractérisation chimique et structurale, transformation et valeur symbolique », PNRC, , p. 20 (lire en ligne [PDF], consulté le ).
- Les plus anciens bijoux de Néandertaliens découverts en Croatie, L'Express, 23/3/2015
- ce serait alors une flûte de type kena : droite, tubulaire, à encoche (sans canal d'insufflation) et à trous de jeu, donc codée 421.111.12 dans la nomenclature de la classification du Système Hornbostel-Sachs (voir la section « organologie » de l'article sur la kena). Plusieurs flûtes en os d'animaux divers, de même type, indiscutables mais un peu plus tardives et plutôt attribuées à Homo sapiens, ont été retrouvées. Voir notamment les flûtes de Geißenklösterle et de Hohle Fels, ou de la grotte d'Isturitz dans les Pyrénées-Atlantiques, datées jusqu'à environ 35 000 à 40 000 ans (AP), ainsi que l'article consacré à la Flûte au Paléolithique.
- Pour le détails des arguments échangés dans ce débat, ainsi que pour ses références scientifiques, consulter l'article qui est consacré à cet objet archéologique.
- (en) Jacques Jaubert, Sophie Verheyden, Dominique Genty, Michel Soulier, Hai Cheng, Dominique Blamart, Christian Burlet, Hubert Camus, Serge Delaby, Damien Deldicque, R. Lawrence Edwards, Catherine Ferrier, François Lacrampe-Cuyaubère, François Lévêque, Frédéric Maksud, Pascal Mora, Xavier Muth, Édouard Régnier, Jean-Noël Rouzaud, Frédéric Santos, « Early Neandertal constructions deep in Bruniquel Cave in southwestern France », Nature, no 534, , p. 111-114 (DOI 10.1038/nature18291).
- Jacques Jaubert, « Que faisait Néandertal dans la grotte de Bruniquel ? », Pour la Science, no 465, , p. 26-35 (résumé).
- (en) Hardy, B.L., Moncel, M., Kerfant, C. et al., « Direct evidence of Neanderthal fibre technology and its cognitive and behavioral implications », Sci Rep, vol. 10, (DOI 10.1038/s41598-020-61839-w, lire en ligne).
- Zilhão, J. et Trinkaus, E. (2002), Portrait of the Artist as a Child : the Gravettian Human Skeleton from the Abrigo do Lagar Velho and its Archeological Context, Trabalhos de Arqueologia, vol. 22, 609 p.
- Plagnol V., Wall J.D., (2006), « Possible ancestral structure in human populations », PLoS Genetics, 2 : e105 ; citation : ..strong evidence for ancient admixture in both a European and a West African population (p ≈ 10−7), with contributions to the modern gene pool of at least 5%. While Neanderthals form an obvious archaic source population candidate in Europe.
- Duarte, C., Maurício, J., Pettitt, P.B., Souto, P., Trinkaus, E., van der Plicht, H. et Zilhão, J., (1999), « The early Upper Paleolithic human skeleton from the Abrigo do Lagar Velho (Portugal) and modern human emergence in Iberia », Proceedings of the National Academy of Sciences, vol. 96, no 13, p. 7604-7609, citation : […] the persistence of a morphological mosaic for that time period, indicates that this child was not the result of a rare Neandertal/early modern interbreeding but the descendant of extensively admixed populations.
- Higham, T., Douka, K., Wood, R., Ramsey, C.S., Brock, F., Basell, L., Camps, M., Arrizabalaga, A., Baena, J., Barroso-Ruíz, C., Bergman, Ch., Boitard, C., Boscato, P., Caparrós, M., Conard, N.J., Draily, Ch., Froment, A., Galván, B., Gambassini, P., Garcia-Moreno, A., Grimaldi, S., Haesaerts, P., Holt, B., Iriarte-Chiapusso, M.-J., Jelinek, A., Jordá Pardo, J.F., Maíllo-Fernández, J.M., Marom, A., Maroto, J., Menéndez, M., Metz, L., Morin, E., Moroni, A., Negrino, F., Panagopoulou, E., Peresani, M., Pirson, S., de la Rasilla, M., Riel-Salvatore, J., Ronchitelli, A., Santamaria, D., Semal, P., Slimak, L., Soler, J., Soler, N., Villaluenga, A., Pinhasi, R. et Jacobi, R. (2014) - « The timing and spatiotemporal patterning of Neanderthal disappearance », Nature, vol. 512, p. 306-309.
- Archéologia, no 525, octobre 2014, p. 8.
- (en) Silvana Condemi, Aurélien Mounier, Paolo Giunti, Martina Lari, David Caramelli, Laura Longo, « Possible Interbreeding in Late Italian Neanderthals? New Data from the Mezzena Jaw (Monti Lessini, Verona, Italy) », PLOS ONE, vol. 8, no 3, (DOI 10.1371/journal.pone.0059781).
- Hervé Morin, « Il y a du Neandertal en nous », Le Monde, (lire en ligne)
- On a tous quelque chose de Neandertal, Rachel Mulot, Sciences et Avenir, juin 2010, page 30.
- Science, Comparative Genomics : the Neandertal Genome
- « Welcome to the family, Homo sapiens neanderthalensis », New Scientist.com, (lire en ligne)
- Sankararaman, S., Patterson, N., Li, H., Pääbo, S. et Reich, D. (2012) - « The date of interbreeding between Neandertals and Modern Humans », PLoS Genetics, 8, (10).
- Hybridation Sapiens et Néandertal, confirmation de la première étude
- Quel est l’apport génétique de Néandertal sur l’humain moderne ?
- Benjamin Vernot, Joshua M. Akey, Resurrecting Surviving Neandertal Lineages from Modern Human Genomes, Science
- Seguin-Orlando, A., Korneliussen, T.S., Sikora, M., Malaspinas, A.-S., Manica, A., Moltke, I., Albrechtsen, A., Ko, A., Margaryan, A., Moiseyev, V., Goebel, T., Westaway, M., Lambert, D., Khartanovich, V., Wall, J.D., Nigst, P.R., Foley, R.A., Lahr, M.M., Nielsen, R., Orlando, L. et Willerslev, E. (2014) - « Genomic structure in Europeans dating back at least 36,200 years », Science, 2014/11/06 [online].
- L'homme de Kostenki précise le métissage Sapiens-Néandertal
- (en) Qiaomei Fu, Mateja Hajdinjak, Oana Teodora Moldovan, Silviu Constantin, Swapan Mallick, Pontus Skoglund, Nick Patterson, Nadin Rohland, Iosif Lazaridis, Birgit Nickel, Bence Viola, Kay Prüfer, Matthias Meyer, Janet Kelso, David Reich & Svante Pääbo, « An early modern human from Romania with a recent Neanderthal ancestor », Nature, no 524, , p. 216-219 (DOI 10.1038/nature14558).
- François Savatier, « L'ADN d'une deuxième femme de Néandertal séquencé », Pour la science, no 482, , p. 16.
- (en) Kay Prüfer, Cesare de Filippo, Steffi Grote, Fabrizio Mafessoni et al., « A high-coverage Neandertal genome from Vindija Cave in Croatia », Science, , article no eaao1887 (DOI 10.1126/science.aao1887).
- (en) Matthew Warren, « Mum’s a Neanderthal, Dad’s a Denisovan: First discovery of an ancient-human hybrid - Genetic analysis uncovers a direct descendant of two different groups of early humans », Nature, vol. 560, , p. 417-418 (DOI 10.1038/d41586-018-06004-0).
- (en) Ann Gibbons, « Modern human females and male Neandertals had trouble making babies ; Here’s why », Science News.org, (lire en ligne).
- (en) Fernando L. Mendez , G. David Poznik, Sergi Castellano et Carlos D. Bustamant, « The Divergence of Neandertal and Modern Human Y Chromosomes », AJHG, vol. 98, no 4, , p. 728-734 (lire en ligne).
- (en) Ann Gibbons, « Neandertals and Moderns Made Imperfect Mates », Science, vol. 343, no 6170, , p. 471-472 (DOI 10.1126/science.343.6170.471, résumé).
- David Reich, Comment nous sommes devenus ce que nous sommes, Quanto, , 380 p. (ISBN 978-2-88915-308-4), p. 74-80.
- Un gène de l'immunité qui nous vient de l’Homme de Néandertal, futura-sciences.com
- Laurent Abi-Rached, The Shaping of Modern Human Immune Systems by Multiregional Admixture with Archaic Humans, Science
- (en) Sebastian Temme, Martin Zaccharias, Jürgen Neumann, Sebastian Wohlfromm, Angelika König, Nadine Temme, Sebastian Springer, John Trowsdale et Norbert Koch « A novel family of human lymphocyte antigen class II receptors may have its origin in archaic human species », Journal of Biological Chemistry].
- (en) Michael Dannemann, Aida M. Andrés, Janet Kelsocorrespondence, « Introgression of Neandertal- and Denisovan-like Haplotypes Contributes to Adaptive Variation in Human Toll-like Receptors », American Journal of Human Genetics, vol. 98, no 1, , p. 22–33, (DOI 10.1016/j.ajhg.2015.11.015).
- (en) Quach, Rotival, Pothlichet, Loh et al., « Genetic Adaptation and Neandertal Admixture Shaped the Immune System of Human Populations », Cell, vol. 167, no 3, , p. 643-656 (DOI 10.1016/j.cell.2016.09.024).
- « Africains et Européens ont des systèmes immunitaires génétiquement différents… et l’Homme de Néandertal y est pour quelque chose », Institut Pasteur, (consulté le )
- (en) Posth, C. et al. Deeply divergent archaic mitochondrial genome provides lower time boundary for African gene flow into Neanderthals. Nat. Commun. 8, 16046, 2017
- (en) Michael Price, « Africans, too, carry Neanderthal genetic legacy », Science, (lire en ligne).
- Gautier Cariou, « Un grand-père moderne pour les Néandertaliens », sur CNRS, (consulté le ).
- (en) Martin Petr, Mateja Hajdinjak, Qiaomei Fu, Elena Essel, Hélène Rougier6 et al., « The evolutionary history of Neanderthal and Denisovan Y chromosomes », Science, vol. 369, no 6511, , p. 1653-1656 (DOI 10.1126/science.abb6460).
- (en) Liming Li, Troy J. Comi, Rob F. Bierman et Joshua M. Akey, « Recurrent gene flow between Neanderthals and modern humans over the past 200,000 years », Science, vol. 385, no 6705, (DOI 10.1126/science.adi17).
- (en) D. Mylopotamitaki et al., « Homo sapiens reached the higher latitudes of Europe by 45,000 years ago », Nature, vol. 626, , p. 341-346 (lire en ligne)
- Erin Blakemore, « Qui étaient vraiment les Néandertaliens ? », sur nationalgeographic.fr, (consulté le )
- Le Monde avec AFP, « La disparition de l’homme de Néandertal serait plus ancienne que ce que l’on croyait », sur lemonde.fr, (consulté le )
- Pettitt. P.B. (1999) « Disappearing from the World: An Archeological Perspective on Neanderthal Extinction », Oxford Journal of Archeology, 18, p. 217-239.
- Guk, Y. (1997) « Why's the Neanderthal Gone? », Science in Russia, 4, p. 50-54.
- Gelly, R. (1992) « L'énigme de Néandertal », Sciences et Avenir, no d'avril, p. 64-69.
- Flores, J. C. (1998) « A Mathematical Model for Neanderthal Extinction », Journal of Theoretical Biology, Volume 191, numéro 3, p. 295-298.
- R.D. Horan, E. Bulte et J.F. Shogren, « How trade saved humanity from biological exclusion: an economic theory of Neanderthal extinction », Journal of Economic Behavior & Organization, vol. 58, no 1, , p. 1-29.
- (en) Tom Higham, « The timing and spatiotemporal patterning of Neanderthal disappearance », Nature, no 512, , p. 306–309 (DOI 10.1038/nature13621)
- (en) « Did a volcanic cataclysm 40,000 years ago trigger the final demise of the Neanderthals? », Geological Society of America, ScienceDaily, (lire en ligne)
- Francesco G. Fedele, Biagio Giaccio, Roberto Isaia et Giovanni Orsi, « The Campanian Ignimbrite eruption, Heinrich Event 4, and Palaeolithic change in Europe: A high-resolution investigation », dans Volcanism and Earth's atmosphere, AGU Book, (lire en ligne).
- Benjamin A. Black, Ryan R. Neely et Michael Manga, « Campanian Ignimbrite volcanism, climate, and the final decline of the Neanderthals », Geology, vol. 43, no 5, , p. 411-414 (lire en ligne, consulté le ), p. 411.
- Documentaire Science grand format, L'apocalypse de Néandertal, diffusé sur France 5 le 29 novembre 2018
- (en) « Press Release : December 2010 », sur harvard.edu (consulté le ).
- Hypothèse présentée dans le roman Dance of the Tiger. A novel of the Ice Age (1978) et explicitée dans une note de l'auteur à la fin de l'ouvrage.
- (en) Ann Gibbons, « Neandertals and Moderns Made Imperfect Mates », Science, vol. 343, , p. 471-472 (ISSN 0036-8075 et 1095-9203, PMID 24482455, DOI 10.1126/science.343.6170.471, lire en ligne, consulté le )
- (en) Krist Vaesen et al., Inbreeding, Allee effects and stochasticity might be sufficient to account for Neanderthal extinction, PLoS ONE 14(11), 27 novembre 2019
- (en) Ludovic Slimak, Tharsika Vimala, Andaine Seguin-Orlando et al., A late Neanderthal reveals genetic isolation in their populations before extinction, biorxiv.org, 10 avril 2023, doi.org/10.1101/2023.04.10.536015
- D’Errico, F. et Sánchez Goñi, M.F. (2003), « Neanderthal extinction and the millennial scale climatic variability of OIS 3 », Quaternary Science Reviews, 22, p. 769–788.
- « L'homme de Néandertal n'est pas mort de froid », sur sciencepresse.qc.ca, .
- « 10 scénarios pour une disparition » in: Neandertal : enquête sur une disparition, Les Dossiers de la Recherche, no 24, août-octobre 2006.
- Finlayson, C., Fa, D.A., Finlayson, G., Pacheco, F.G. et Vidal, J.R., (2004), « Did the moderns kill off the Neanderthals? A reply to F. D’Errico and Sánchez Goñi », Quaternary Science Reviews, Volume 23, numéros 9-10, p. 1205-1209.
- La domestication du chien aurait-elle aidé sapiens à supplanter Néandertal ?, Maxisciences, 16 mai 2012.
- Shipman, P., « Dog domestication may have helped humans thrive while Neandertals declined », American scientist, mai-juin 2012, vol. 100, no 3, p. 198
- Marcel Otte, L'Homme de Néandertal. L'extinction, Université de Liège, , p. 27
- (en) Anthony Santino Pagano, Samuel Márquez, Jeffrey T. Laitma, « Reconstructing the Neanderthal Eustachian Tube: New Insights on Disease Susceptibility, Fitness Cost, and Extinction », The Anatomical Record, (DOI 10.1002/ar.24248)
- (en) Dimitra Papagianni et Michael A. Morse, Neanderthals Rediscovered : How Modern Science Is Rewriting Their Story, Thames & Hudson, , 208 p. (ISBN 978-0-500-77311-6, lire en ligne)
Voir aussi
[modifier | modifier le code]Articles connexes
[modifier | modifier le code]- Châtelperronien
- Micoquien
- Moustérien
- Paléolithique moyen
- Projet génome de Néandertal
- Hybridation entre les humains archaïques et modernes
Bibliographie
[modifier | modifier le code]- « Neandertal : enquête sur une disparition », Les Dossiers de La Recherche, no 24, (ISSN 1772-3809, présentation en ligne).
- Collectif, Neandertal, Paris, Tallandier, 2006 [version mise à jour des textes figurant dans le no 24 des Dossiers de la Recherche] (ISBN 978-2-84734-366-3).
- Collectif, Les Néandertaliens : biologie et cultures, dir. Bernard Vandermeersch et Bruno Maureille, coll. Documents préhistoriques, Paris, CTHS, 2007 (ISBN 978-2-7355-0638-5).
- Collectif, Néandertal l'Européen, dir. Stéphane Pirson et Michel Toussaint, 2e éd. revue et corrigée, Namur, Institut du patrimoine wallon, 2011 [catalogue d'une exposition organisée à Namur du 4 septembre 2010 au 3 avril 2011] (ISBN 978-2-87522-069-1) (BNF 42609978).
- Collectif, Neandertal : notre nouvel ancêtre : la preuve par l'ADN, Les Dossiers de la Recherche, no 45, octobre 2011 (ISSN 1772-3809).
- Collectif, Néandertal réhabilité, Dossiers d'archéologie, no 345, mai-juin 2011 (ISSN 1141-7137).
- Collectif, L'homme de Neandertal et l'invention de la culture, Dossier pour la science, no 76, juillet 2012 (ISSN 1246-7685).
- Juan Luis Arsuaga, Le collier de Néandertal : nos ancêtres à l'ère glaciaire, Paris, Odile Jacob, 2001 [l'original espagnol a paru en 1999] (ISBN 978-2-7381-1422-8).
- Claudine Cohen, Un Néandertalien dans le métro, coll. Science ouverte, Paris, Éd. du Seuil, 2007 (ISBN 978-2-02-092557-0).
- Jean-Jacques Hublin et Bernard Seytre, Quand d'autres hommes peuplaient la terre : nouveaux regards sur nos origines, coll. Champs Sciences, Paris, Flammarion, 2011 [rééd. revue d'un ouvrage paru en 2008] (ISBN 978-2-08-125242-4).
- Jacques Jaubert, Chasseurs et artisans du Moustérien, coll. Histoire de la France préhistorique, Paris, La Maison des Roches, 1999 (ISBN 978-2-912691-05-7).
- Bruno Maureille, Les premières sépultures, Paris, Le Pommier / Cité des sciences et de l'industrie, 2004 (ISBN 978-2-7465-0203-1).
- Bruno Maureille, Qu'est-il arrivé à l'homme de Néandertal ?, coll. Les petites pommes du savoir, Paris, Le Pommier, 2008 (ISBN 978-2-7465-0389-2).
- Svante Pääbo, Néandertal : à la recherche des génomes perdus, Les liens qui libèrent, 2015 (ISBN 979-10-209-0321-1).
- Marylène Patou-Mathis, Néandertal de A à Z, Allary Éditions, , 640 p. (ISBN 978-2-37073-160-9, présentation en ligne, lire en ligne).
- Thomas Wynn et Frederick L. Coolidge, How to think like a Neandertal, New York, Oxford University Press, 2012 (ISBN 978-0-19-974282-0).
- Silvana Condemi et François Savatier, Néandertal, mon frère, Flammarion, , 263 p. (lire en ligne).
- David Reich, Comment nous sommes devenus ce que nous sommes, Quanto, , 380 p. (ISBN 9782889153084).
Filmographie
[modifier | modifier le code]- Neanderthal, docu-fiction télévisé britannique (durée 100 min) réalisé par Tony Mitchell en 2000 (diffusion en 2001) (OCLC 47173335)
- Il était une fois… l'Homme : l'Homme de Néandertal, dessin animé de Albert Barillé et Fabrice Matéo
- Interview de Jean-Louis Heim et Marylène Patou-Mathis au sujet de l'Homme de Néandertal (2007 ?)
Liens externes
[modifier | modifier le code]- « Neandertal, une histoire belge ? », Carbone 14, le magazine de l'archéologie, France Culture, 16 mars 2024.
- Le musée de l'Homme de Néandertal de La Chapelle-aux-Saints, en France
- Exposition sur l'Homme de Néandertal, au Musée de l'Homme, à Paris (2018)
- (en) Le musée de Néandertal, en Allemagne, sur le site originel de 1856
- (en) New Portuguese skull may be an early relative of Neandertals – article de Ann Gibbons sur Science, 13 mars 2017
- (en) In Neanderthal DNA, Signs of a Mysterious Human Migration – article de Carl Zimmer, NY Times, 4 juillet 2017
- (en) Neanderthal or Neandertal ?
- (en) Vidéo Homo sapiens vs Neandertals – The Evolution of Language, aussi sur Epic History TV (2016, 18')
- Site de l'homme de Néandertal de Krapina (Croatie) sur culture.ec.europa.eu/fr/
- Kraneamus, visite du musée Néandertal de Krapina, sur mkn.mhz.hr/en
Bases de données et dictionnaires
[modifier | modifier le code]
- Ressources relatives au vivant :
- Ressource relative à la santé :
- Notices dans des dictionnaires ou encyclopédies généralistes :