鳥
此條目需要補充更多來源。 (2013年9月10日) |
| 鳥綱 化石時期:分子鐘推定冠群鳥類於1.21億年前出現
| |
|---|---|

| |
| 科學分類 | |
| 界: | 動物界 Animalia |
| 門: | 脊索動物門 Chordata |
| 演化支: | 鳥翼類 Avialae |
| 演化支: | 真鳥翼類 Euavialae |
| 演化支: | 短尾鳥類 Avebrevicauda |
| 演化支: | 尾綜骨鳥類 Pygostylia |
| 演化支: | 鳥胸骨類 Ornithothoraces |
| 演化支: | 真鳥類 Euornithes |
| 演化支: | 今鳥型類 Ornithuromorpha |
| 演化支: | 扇尾類 Ornithurae |
| 綱: | 鳥綱 Aves Linnaeus, 1758 |
| 演化支 | |
| 異名 | |
鳥或者鳥類,是鳥綱(學名:Aves)動物的通稱,是唯一存活並演化至今的鳥翼類,其全體成員均為兩足、恆溫、卵生、身披羽毛且色彩鮮艷各異、前肢特化成翅膀、具有堅硬的喙,多數可飛行,小部分純陸生或水生,骨骼中空輕盈並有強力肌肉附著,以及唯一能使用氣囊行雙重呼吸的動物。
目前全球鳥類共有萬餘種,按照世界鳥類學家聯合會的分類系統,共有11093個物種,按照國際鳥盟的分類系統則有11162個物種,更多不同分類系統的數字也大致相同。鳥類的食物多種多樣,包括植物的花蜜、種子、果實,昆蟲、魚類,蛙類、蜥蜴類、蛇類、龜類、小型哺乳動物,魷魚類、貝類、和其它鳥類,以及動物的蛋、腐肉、骨骼和人類的生活垃圾。鳥的體型大小不一,最大的鴕鳥體高可達2.5米,而最小的吸蜜蜂鳥體長僅5厘米。多數的鳥在日間活動,也有一些鳥在夜間或黃昏的時候活動(例如夜鷹、貓頭鷹等)。候鳥會進行長距離遷徙以尋找最佳棲息地,而留鳥則一般不會大幅移動。大多數鳥類都會飛行,不能飛的鳥包括古顎總目下的鴕鳥、鴯鶓、幾維鳥等多數種類,企鵝目的全部鳥類,某些生活在沒有地面捕獵者的孤立海島上的鳥類如鸚鵡目的鴞鸚鵡,也在演化過程失去了飛行能力。
現代所有鳥類在生物學上也被分類為鳥形恐龍(即鳥翼類)的一部份,過去廣義的鳥綱曾籠統地包括始祖鳥、孔子鳥、黃昏鳥等已滅絕的史前類群,但現在比較常用鳥翼類(Avialae)來包括這些不再屬於現代鳥類的史前近親[3],包括擅攀鳥龍科、反鳥類和扇尾亞綱;現代定義的鳥綱包括了現生鳥類的最近共同祖先及其後代。根據分類學的研究,證實了恐龍是鳥的祖先,因此其直系後代的鳥類也是研究已滅亡恐龍的最主要的活體材料。鳥綱用科學上最嚴謹的說法,就是「鳥翼類的冠群」[4][5]。目前所有鳥類物種,都是在1億5,000萬年前的侏羅紀~白堊紀這一段時間內,由恐龍總目獸腳亞目虛骨龍類-手盜龍類中的近鳥型恐龍分批次逐步演化而來[6][7]。
在白堊紀末滅絕事件之後,鳥類的祖先非鳥型恐龍全滅,大量鳥形恐龍和現代鳥類在事件之後的幾千萬年中滅亡,只留下化石痕跡,如加斯頓鳥形目。不過,鳥類很快在恐龍大滅絕過後的數萬年中重新佔據了地球上的各個中上層生態位維持至今。人類進入現代文明後,許多鳥類才因為人類的過度狩獵、捕捉、或農業生產破壞其棲息地而再次滅絕,如渡渡鳥、大海雀、恐鳥、象鳥、旅鴿等,不過相對來說,鳥類依靠著飛天能力、較高的智商和強健的肌肉附著力,其適應能力在脊椎動物中是非常強大的,即使在人類出現後也有大批鳥類出現數量增長,像例如多種麻雀與烏鴉高度適應人造環境;而雞、鴨與鵝等等則為人類馴化,並大量飼養。
定義
[編輯]在過去,學界對鳥綱的具體範圍有不一致定義,從包含始祖鳥到只包含現存鳥類冠群的都有,嘉克斯·高斯特指出,鳥綱的定義與用法共有四種互相衝突的定義。嘉克斯·高斯特建議採用第四個定義,並為其它定義提出替代分類:
- 具有羽毛的先進主龍類,此定義可以被鳥羽類(Avifilopluma)取代。
- 恐龍中具有羽毛且能夠飛行的,此定義可以被鳥翼類(Avialae)取代。
- 在蜥形綱之中,較接近鳥類,而離鱷魚較遠的物種,此定義可以被鳥跖類(Avemetatarsalia)取代。
- 所有現存鳥類的最近共同祖先,與最近共同祖先的所有後代,此冠群定義會讓新鳥亞綱(Neornithes;中文名也曾稱為今鳥類或今鳥亞綱)成為鳥綱的次異名[8]。
根據第四個定義,始祖鳥屬於鳥翼類,但不屬於鳥綱。嘉克斯·高斯特的建議如今已被古動物學者與鳥類學者廣泛採用,只是這些學者偶爾也會不一致地把一些鳥翼類通稱為「鳥」[9]。
形態
[編輯]鳥綱的特徵是有羽毛、喙沒有牙齒、蛋有硬殼、高代謝率、心臟有四室、輕盈但結實的骨骼。所有鳥類的前肢都進化成翼,大部份也能夠飛翔。它們有獨特的消化系統及呼吸系統,很適合飛行。一些鳥類,如鴉科及鸚鵡等是最有智慧的動物物種;有些鳥類甚至懂得製造及使用工具,很多物種也有傳遞知識的能力。
很多鳥類每年都會遷徙一段很長距離,也有不規則地移動短距離。它們會用視覺訊號及唱歌來溝通,也會進行合作繁殖及獵食。大部份鳥類都是有固「一夫一妻制」的,往往經歷整個繁殖季節,有時多年,但很少是畢生的。有些物種也會出現配對外交配;其他物種都是「一夫多妻制」或「一妻多夫制」的。它們會鳥巢中下蛋,並由雙親所孵化。大部份鳥類在孵化後都會由雙親照顧。
解剖特徵
[編輯]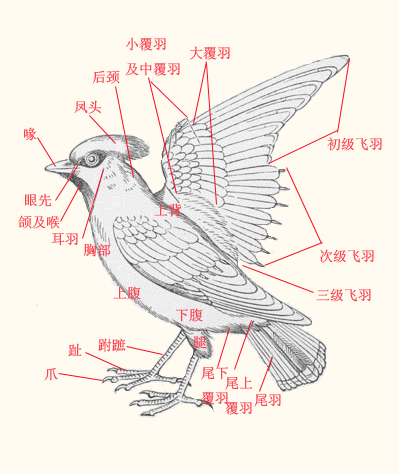
鳥類有許多特化的適應飛翔生活的解剖特徵,例如:
- 骨骼中空且高度癒合:鳥類骨骼第一特徵是硬且輕,主因密骨質較厚而呈中空,可有效減輕體重。鳥類的頸椎數量較其餘陸生動物為多,而胸椎以下骨骼癒合程度極高,如其脊椎相互癒合成若干塊聯合椎骨,不似哺乳動物椎骨間可以在一定限度內自由活動。骨骼的高度癒合有利於提高鳥類的骨強度及維持飛行姿勢,減低骨骼自重,是適應飛翔生活的特徵。
- 胸骨特別發達:除平胸目外,胸骨皆很發達,此可增加肌肉附著面積,故胸肌特別發達以利飛行。
- 體表被羽:羽毛在發生學上於爬行動物的鱗片來源相同,且構造上與哺乳動物的毛髮截然不同,簡單來說鳥類的羽毛特徵是具有分枝。羽毛的出現是鳥類適應高空飛翔寒冷環境所必須的,並且通過換羽來更換。鳥類的羽毛分正羽、副羽和絨羽。
- 呼吸系統:鳥類的呼吸系統與眾不同,其進行氣體交換的基本單元不是肺泡而是微氣管,而且具有分布於全身各處骨骼內的氣囊,吸氣時空氣經微氣管從體外進入氣囊,進行第一次氣體交換,呼氣時氣體經微氣管從氣囊排出體外,進行第二次氣體交換,因此鳥類每呼吸一次,氣體交換兩次,氣體交換效率明顯高於哺乳動物。另外氣囊的存在也進一步增加鳥類的浮力及減緩飛行壓力變化對身體的衝擊。
- 生殖和排泄系統:無論雄雌鳥類的生殖系統僅在繁殖季節膨大並產生功能,非繁殖季節其生殖系統便會萎縮,即便解剖也很難發現。鳥類以不溶於水的尿酸為主要代謝物,以上特徵可以有效地減輕鳥類自身體重。
- 鳥類全無牙齒:而演化出角質化的喙,並依採食習性的不同,而演化出多樣化的形狀,此對減輕體重有很大的幫助。
- 具有嗉囊及砂囊:為彌補缺少牙齒的劣勢,許多鳥類在食道與腺胃間具有嗉囊,主要功能用於儲存部份食物,取代部份胃的功能。鳥類的腺胃較小,食物停留時間較短。在腺胃之後,還有砂囊的存在。鳥類會吞食石塊,利用砂囊強健肌肉磨碎食物,取代牙齒及口腔咀嚼功能。
- 鳥類的消化系統明顯短於哺乳動物,這也是造成鳥類頻繁排遺的一個原因,但較短的消化系統有助於減輕鳥類的體重。
- 氮的代謝不同於哺乳動物:為了有效減輕重量,在氮的代謝,含氮物質最後轉變為毒性較低的尿酸,較哺乳動物排放的尿素相比,不需大量水份稀釋,故鳥類缺乏液態的尿液。
- 鳥類的特殊結構:羽毛、喙、蠟膜、孵卵斑、距、尾脂腺、嗉囊。
演化過程
[編輯]根據古生物學的化石記錄,可以確定鳥類是恐龍演化出來的、也是目前恐龍總目的下屬中唯一存活下來的物種,因為鳥類和非鳥恐龍這兩者在生物學上的構造極其相近,所以現今生物學界已將鳥類認定為存活至現代的恐龍後代。鳥類的第一次出現溯源於中生代侏羅紀,最早的祖先是出土於中國遼寧的傷齒龍科恐龍「始中國羽龍」,而最有名的則是德國發現的始祖鳥;他們的身體特徵都同時具有鳥類和爬行類共有的特徵,例如有牙齒,前肢(翅膀)有指爪等。在中國東北新發現的中華龍鳥和孔子鳥被認為是連接恐龍和鳥類的一環,更像是有羽毛的恐龍,比始祖鳥的年代更久遠。所以說,恐龍其實並沒有完全滅絕,恐龍中的一部分演化成了鳥類;但將鳥和恐龍定為同一類依然不被社會大眾所熟知,因為非鳥恐龍等生物存在了一億年的時間,在日常生活中「非鳥恐龍」一詞不夠簡潔有力,如果要形容三疊紀~白堊紀中存活的非鳥恐龍,一般人還是習慣直接使用「恐龍」一詞;不過無論如何,至少可以確定在生物學界的主流觀點中,鳥類的祖先必定是恐龍。[10][11][12]雖然多數學者都支持恐龍是鳥類的祖先的說法,但仍然有鳥類學家對此提出質疑,並堅持鳥類起源於三疊紀的槽齒類爬行動物假說。[13]
但不管鳥類演化出來的「成因」如何,鳥類這種生命形態一旦出現以後就開始急速進化,以非常短的時間脫離了原本非鳥恐龍的生態位並適應了陸地中高階層和獨霸天空的生態位,以爆發式的增長的速度演化出了非常多種鳥類,有些更是出現了非鳥恐龍所沒有的新生命形式(例如:企鵝目需要長時間依賴在海水中才能存活,然而沒有一種非鳥恐龍是必須依賴海水的。)據一般推測,第三紀中新世是鳥類的全盛時期,後來冰期來臨、種群衰退,據估計歷史上曾經存在過大約10萬種鳥,而在冰河期之後只有10,000種、20餘目倖存,約為之前的十分之一;不過由於鳥類對各種環境的適應力極高,所以到了現代又輻射演化出更多的新物種。
| |||||||||||||||||||||||||||||||||||||||
鳥的分類
[編輯]鳥綱分為兩個下綱:今顎下綱及古顎下綱。視乎科學分類的觀點,鳥類物種的現存數量超過12000個。一般認為鳥綱演化自白堊紀,分裂成古顎下綱及今顎下綱,並於白堊紀-第三紀滅絕事件前分裂為雞雁小綱及其他今顎下綱。其他今顎下綱的演化則有不同意見,根據分子證據是指向白堊紀演化,而化石證據則指向第三紀的演化。統合分子及化石證據的假說卻備受質疑。[15][16]
鳥綱的分類在細節上也常常受到質疑,在2010年代之前學者們對於很多目級別之間的鳥類演化關係並未有共識;不過因為2014年基於鳥類骨頭的全基因組測序分類被發表出來,大部分化石及DNA分子證據都為「鳥綱」提供了更為清晰的演化脈絡。
按照不同的分類原則可以將鳥分成不同的類別,目前主要有三大通行的分類系統:
「鄭作新」分類法
[編輯]第一個是中國鄭作新院士以鳥類形態學特徵為基礎創立的分類系統,雖然目前在國際上這個分類法已經不常用,但在中國不少鳥類研究學者中因為簡單易記、符合中文圈常用詞彙等原因仍為樂於被採用,而且在非中國大陸的港澳台地區和日韓也有少部分學者還在使用此分類。
按鳥類的生活環境和形體特徵,可以將突胸總目的鳥類分為六大生態類群:
- 游禽:可以在水面游動並主要在水面游弋的鳥類,包括鴨雁類,鷗類等。
- 涉禽:在灘涂濕地涉水活動但多不會游泳,常具有「三長」——腿長、頸長、嘴長的特徵,包括鶴類,鸛類,鴴鷸類等。
- 鳴禽:即雀形目鳥類,為鳥類中進化程度最高的一個類群。
- 攀禽:適應攀援生活的鳥類其趾形多為對趾足或轉趾足,包括啄木鳥、鸚鵡等。
- 陸禽:生活在地面的鳥類,其體態特徵適合在地面行走,常飛行能力不強,包括雉類、鶉類和鳩鴿類等。
- 猛禽:以其他動物為食物的鳥類,具有適應捕獵生活的特徵如銳利的腳爪和喙,敏銳的視覺,主要包括鷹、隼、鷂、鶚、鵟、鴞等。
「鳥類DNA」分類法
[編輯]第二個是曾經被中國之外的研究者廣泛採用的鳥類DNA分類系統。1980年代Sibley和Monroe根據DNA雜交實驗研究成果,以鳥類的進化過程為依據衡量的科和屬之間親緣關系所建立的分類系統,這個分類系統較之以前應用的鄭系統有着較大的調整,其中最為引人矚目的是擴大的鸛形目和鴉科。由於DNA-DNA雜交被證實並不適用解釋單系群的內部關係。而且鑑於1980年代中無論電腦技術、醫學、還是遺傳學都和現代有較大差距,故這一分類已被棄用,由更加新穎準確的系統代替,詳細的分類系統見鳥類DNA分類系統,不過此分類在東亞文化圈以外的地區有著強大的歷史影響力。
「全基因組測序」分類法
[編輯]第三個是2014年公布的基於全基因組測序的鳥類分類系統,此分類系統目前被廣泛採用,是目前人類科技所能達到最準確的分類法,嚴格比較基因順序而得出各個物種之間的關係為何。基於全基因組測序的鳥類分類系統[17][18]:
- 古顎下綱 Palaeognathae
- 今顎下綱 Neognathae
系統發育樹
[編輯]下圖基於國際鳥類學大會(IOC)於2019年公布的鳥類發育樹[19][20],括號內的數字為該目下的物種總數(基於IOC World Bird List, v12.1)。
| 鳥綱 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Aves |
鳥的分布
[編輯]在大部份陸地的棲息地中都可以找到鳥類,鳥類分佈在七大洲,連南極洲距海440公里的內陸都可以找到雪海燕[21]。鳥的多樣性在熱帶地區最為顯著。早期認為熱帶地區的高多樣性是高度物種形成的結果,但近來的研究發現高緯度地區也有高度物種形成,但被較熱帶地區要高的滅絕速率所抵消。[22]。許多科的鳥類可以在陸地及海洋中生活,有些海鳥上岸只是為了繁殖[23],有些企鵝可以潛到300公尺深的海中[24]。
鳥類和人類
[編輯]馴養和娛樂
[編輯]
鳥類是人類的一個重要的食物來源。通常食用的是肉和卵,主要來自鵝(雁)、雞、火雞和鴨等。其他可以作為人類食物的鳥類還包括鴯鶓、竹雞、鴕鳥、鴿、松雞、鵪鶉、鷸、鳴禽和小的燕雀類的鳥等,雞和鴨分別是每年人類屠宰的動物中數量最多和第二多的,在2013年,全球有超過六百一十億隻雞和超過二百九十六億隻鴨被宰殺。[25]天鵝和火烈鳥這些現在被保護的鳥也曾成為富人餐桌上的美味。

有許多種被馴化鳥可以為人類所利用,例如信鴿可以用來傳送信息,獵鷹可以用來打獵,魚鷹可以用來捕魚。 雞和鴿子在生物學和比較心理學方面的實驗研究中也得到廣泛應用。因為鳥對毒素特別敏感,所以在煤礦里金絲雀經常被用來判斷有毒氣體是否濃度過高,以便礦工能及時逃走。
人工飼養的鳥類還對野生種群構成另一個危害:對野生種的入侵。比如人工選種優育的猛禽往往身體強壯,體形較大,如果流入野生環境,對野生鳥類的是很大的威脅。所以歐美各國嚴禁野放人工種,並對此採取必要的監測措施。[來源請求]
把野生鳥類作為寵物飼養在中國有着很長的歷史,並發展出了各種名目。主要分聽唱和玩耍兩種類型,聽唱涉及的鳥類主要是雀形目的鳴禽,如山雀、百靈、畫眉、鷯哥、八哥等;用於各種玩耍的鳥類主要涉及部分雀形目雀科、伯勞科的鳥類以及隼形目的猛禽,如黑頭蠟嘴雀、錫嘴雀以及各種猛禽。
世界各地幾乎都有鬥雞的娛樂傳統,這種利用雞形目的鳥(也有其他目的)在發情期好鬥的特點,來進行比賽。
對鳥類的狩獵活動,伴隨着人類發展的歷史。一種狩獵方式是以鳥類為對象,有些地方由於過度狩獵,很多鳥類已經滅絕,如旅鴿。還有一些是利用鳥類捕食的能力,來幫助人抓住獵物。女真人訓練海東青捕捉天鵝取珠。[26]
非法鳥類貿易和人類活動的威脅
[編輯]
在世界各國,都有把色彩艷麗的,特別是熱帶的鳥(例如,鸚鵡和某些椋鳥科鳥類)作為寵物來飼養的習慣,這也導致一些瀕危的鳥類被走私。在抓捕和販運過程中野生鳥類死亡率極高(有說法稱最終消費者手中的每一隻野生鳥類背後有二十隻同類的屍體),因此飼養野鳥是一種對野生動物資源有着極大破壞力的陋習,它對鳥類生存的威脅從某種程度上講,甚至超過了偷獵。
由於高額利潤的驅使,鳥類貿易直接威脅着許多鳥種的生存。許多歐美國家都有馴養猛禽的許可證制度,允許人們合法飼養猛禽,接受政府的嚴格管理,但在購入渠道上仍存在非法交易。在阿拉伯世界,猛禽被視為高貴身份的象徵,許多富商不惜重金購入猛禽。
過度的狩獵活動是一些鳥類數量急劇減少以至滅絕的直接原因,如旅鴿。還有一些鳥類的滅絕或瀕危是由於棲息地被破壞、森林砍伐和農業耕作等人類活動,如朱䴉。隨着人類活動範圍的擴大和影響的加強,比如有毒化學物質在生物體內的聚集等原因,都直接威脅着鳥類的安全。
鳥害
[編輯]
由於人類活動的影響,有些鳥類在全世界範圍內泛濫成災。例如,鴿子和烏鴉就在全世界很多城市的市區過度繁殖,成為困擾各大都市的一個公共衛生問題。
另外,棲息在機場附近的野生鳥類會對起降中的飛機飛行安全造成一定威脅。這種飛鳥與飛機相撞造成的飛行安全事故稱為鳥擊,歷史上最早的鳥撞事故記錄是發生1912年的美國,飛行員卡爾洛德傑駕駛的飛機在加利福尼亞上空和一隻海鷗相撞,機毀人亡。隨着噴氣式飛機的出現和人類航空產業的發展,目前全世界機場數量飛機起降架次不斷增長,鳥撞問題日益凸現,據有關資料統計,全世界每年約有7500架次飛機受到不同程度的鳥撞,損失高達100億美元。目前解決鳥撞問題的主要方法是驅散棲息在機場附近的鳥類,具體說來,可以採取馴養猛禽驅趕、破壞棲息地、發出巨大和恐怖的聲音等不同方式。
鳥與人的共患病
[編輯]能夠傳染給人類的鳥類疾病包括:鸚鵡熱、沙門氏菌病、彎曲菌病、紐卡斯爾的疾病,分枝桿菌病(鳥的結核病)、禽流感、賈第鞭毛蟲病和隱孢子蟲病。
文化
[編輯]因其身體被羽毛覆蓋,中國古代動物學將其統稱為羽蟲。梁實秋寫過一篇《鳥》的文章:「鳥的身軀都是玲瓏飽滿的,細瘦而不乾癟,豐腴而不臃腫,真是減一分則太瘦,增一分則太肥那樣的穠纖合度,跳蕩得那樣輕靈,腳上像是有彈簧。看它高踞枝頭,臨風顧盼——好銳利的喜悅刺上我的心頭。」
參考文獻
[編輯]- ^ Field, Daniel J.; Benito, Juan; Chen, Albert; Jagt, John W. M.; Ksepka, Daniel T. Late Cretaceous neornithine from Europe illuminates the origins of crown birds. Nature. 2020-03-19, 579 (7799) [2022-10-15]. ISSN 0028-0836. doi:10.1038/s41586-020-2096-0. (原始內容存檔於2022-07-07) (英語).
- ^ Class Aves - Classification - Systema Naturae 2000. web.archive.org. [2022-10-15]. (原始內容存檔於2009-06-01).
- ^ ギル 『鳥類學』 (2009)、30頁
- ^ ギル 『鳥類學』 (2009)、626頁
- ^ 山階鳥研 (2006)、16頁
- ^ Borenstein, Seth. Study traces dinosaur evolution into early birds. AP News. 2014-07-31 [2015-03-08]. (原始內容存檔於2018-10-23).
- ^ Lee, Michael S. Y.; Cau, Andrea; Naish, Darren; Dyke, Gareth J. Sustained miniaturization and anatomical innovation in the dinosaurian ancestors of birds. Science. 2014-08, 345 (6196) [2022-10-15]. ISSN 0036-8075. doi:10.1126/science.1252243. (原始內容存檔於2022-10-15) (英語).
- ^ Gauthier, J., and de Queiroz, K. (2001). "Feathered dinosaurs, flying dinosaurs, crown dinosaurs, and the name Aves." Pp. 7-41 in New perspectives on the origin and early evolution of birds: proceedings of the International Symposium in Honor of John H. Ostrom (J. A. Gauthier and L. F. Gall, eds.). Peabody Museum of Natural History, Yale University, New Haven, Connecticut, U.S.A.
- ^ Godefroit, Pascal; Cau, Andrea; Dong-Yu, Hu; Escuillié, François; Wenhao, Wu; Dyke, Gareth. A Jurassic avialan dinosaur from China resolves the early phylogenetic history of birds. Nature. 2013-06, 498 (7454) [2022-10-15]. Bibcode:2013Natur.498..359G. ISSN 0028-0836. PMID 23719374. doi:10.1038/nature12168. (原始內容存檔於2018-05-29) (英語).
- ^ Birds are Dinosaurs - American Museum of Natural History. [2014-01-17]. (原始內容存檔於2014-03-28).
- ^ 牠是恐龍還是鳥?. [2014-01-17]. (原始內容存檔於2012-10-02).
- ^ 【專題報導】恐龍與鳥的秘密. [2014-01-17]. (原始內容存檔於2014-02-26).
- ^ 恐龍奧秘- 恐龍與鳥類關係之奧秘- 鳥類起源於恐龍的觀點已經完全確立了嗎?. [2014-01-17]. (原始內容存檔於2014-03-03).
- ^ Chiappe, Luis M. Glorified Dinosaurs: The Origin and Early Evolution of Birds. Sydney: University of New South Wales Press. 2007. ISBN 978-0-86840-413-4.
- ^ Ericson, Per G.P; Anderson, Cajsa L; Britton, Tom; Elzanowski, Andrzej; Johansson, Ulf S; Källersjö, Mari; Ohlson, Jan I; Parsons, Thomas J; Zuccon, Dario; Mayr, Gerald. Diversification of Neoaves: integration of molecular sequence data and fossils. Biology Letters. 2006-12-22, 2 (4) [2022-10-15]. ISSN 1744-9561. PMC 1834003
. PMID 17148284. doi:10.1098/rsbl.2006.0523. (原始內容存檔於2022-11-27) (英語).
- ^ Brown, Joseph W; Payne, Robert B; Mindell, David P. Nuclear DNA does not reconcile ‘rocks’ and ‘clocks’ in Neoaves: a comment on Ericson et al .. Biology Letters. 2007-06-22, 3 (3) [2022-10-15]. ISSN 1744-9561. PMC 2464679
- ^ Prum, Richard O.; Berv, Jacob S.; Dornburg, Alex; Field, Daniel J.; Townsend, Jeffrey P.; Lemmon, Emily Moriarty; Lemmon, Alan R. A comprehensive phylogeny of birds (Aves) using targeted next-generation DNA sequencing. Nature. 2015-10, 526 (7574) [2022-10-15]. ISSN 1476-4687. doi:10.1038/nature15697. (原始內容存檔於2020-04-09) (英語).
- ^ Yuri, Tamaki; Kimball, Rebecca; Harshman, John; Bowie, Rauri; Braun, Michael; Chojnowski, Jena; Han, Kin-Lan; Hackett, Shannon; Huddleston, Christopher; Moore, William; Reddy, Sushma. Parsimony and Model-Based Analyses of Indels in Avian Nuclear Genes Reveal Congruent and Incongruent Phylogenetic Signals. Biology. 2013-03-13, 2 (1) [2022-10-15]. ISSN 2079-7737. PMC 4009869
- ^ Orders of Birds – IOC World Bird List. [2022-10-18]. (原始內容存檔於2022-11-13) (美國英語).
- ^ IOCTree v2 (PDF). [2022-10-18].
- ^ Brooke, Michael. Albatrosses And Petrels Across The World. Oxford: Oxford University Press. 2004. ISBN 0-19-850125-0.
- ^ Weir, Jason T.; Schluter, D. The Latitudinal Gradient in Recent Speciation and Extinction Rates of Birds and Mammals. Science. 2007, 315 (5818): 1574–76. ISSN 0036-8075. PMID 17363673. doi:10.1126/science.1135590.
- ^ Schreiber, Elizabeth Anne; Joanna Burger. Biology of Marine Birds. Boca Raton: CRC Press. 2001. ISBN 0-8493-9882-7.
- ^ Sato, Katsufumi; Naito, Y.; Kato, A.; Niizuma, Y.; Watanuki, Y.; Charrassin, J. B.; Bost, C.-A.; Handrich, Y.; Le Maho, Y. Buoyancy and maximal diving depth in penguins: do they control inhaling air volume?. The Journal of Experimental Biology. 2002-05, 205 (Pt 9) [2022-10-15]. ISSN 0022-0949. PMID 11948196. doi:10.1242/jeb.205.9.1189. (原始內容存檔於2022-12-21).
- ^ FAOSTAT. www.fao.org. [2019-10-25]. (原始內容存檔於2017-05-11).
- ^ 《滿洲源流考》:「海東青。羽族之最鷙者,有海東青焉。身小而健,其飛極高,能擒天鵝;搏兔,亦俊於鷹鶻。」
- ^ Newton, Ian. The Speciation and Biogeography of Birds. Amsterdam: Academic Press. 2003: 463. ISBN 0-12-517375-X.
- 財團法人 台北鳥會野鳥救傷中心 官方網站
- 臺灣 阿迷購(頁面存檔備份,存於網際網路檔案館) 官方網站
外部連結
[編輯]- 延陵動物志(鳥綱) / FAUNA NGIANA (AVES) 卷一
- 中國自然網-鳥類全書
- 鳥的分類
- 鳥類的名稱(頁面存檔備份,存於網際網路檔案館)(中文)(日語)(越南文)
參見
[編輯]- 恐龍
- 觀鳥
- 鳥類環志
- 鳥類全基因組測序分類系統
- Sibley和Monroe的鳥類DNA分類系統