鸟类生理解剖学

鸟类生理解剖学,或称鸟类躯体生理学,通常研究的是鸟类所独有的适应特征,这些特征通常都是为了适应飞行所需。鸟类通常拥有轻盈的骨架、轻且强壮的肌肉、能支撑高速新陈代谢和氧气供应的循环系统及呼吸系统,還有羽毛。这些加起来,才使得鸟类能够飞行。而喙的出现则导致鸟类演化出特别的消化系统。鳥類還有較大視覺感知範圍與發達的腦部。
这些特殊的解剖学特征,使得鸟类在脊椎动物门中占据了一个独立的纲——鸟纲。
呼吸系统
[编辑]

由于飞行需要充足的氧气,鸟类发展出一种高效的呼吸系统,稱作雙重呼吸。该系统可以分为三个不同的部分:1、前气囊(分别位于锁骨、颈以及胸前部);2、肺;3、后气囊(分别位于腹部和胸后部)。鸟类的肺部只能使气流通过而不能儲存,而负责儲存空气的则是气囊。而气囊并不负责气体的交换,其作用和风箱类似,让新鲜空气以较为恒定的速度,源源不断的反覆通过肺部[1]。前气囊和后气囊加起来通常是9个,其中只有锁骨气囊是单个出现,其它的气囊都是成对出现的。有的如雀形目,气囊的数量是7个,其胸前气囊和锁骨气囊是相通的,甚至是融合到了一块。

鸟类吸气的时候,吸入的新鲜空气有一半会直接进入后气囊,另一半则经由肺部交換後进入前气囊。在呼气的时候后气囊的新鲜空气会经由肺部进行气体交换之后排出,而前气囊的低含氧量空气则不经过肺部直接排出体外。被交换过的高二氧化碳含量的气体不会再次经过肺部,而哺乳类动物则与此有很大差异:后者的肺部负责儲存和交换气体,因此会发生新鲜空气和废气混合的情况,或者说含氧气分压会逐渐发生变化。除此之外,鸟类的肺部在吸气和呼气时都会有新鲜空气以固定方向流经肺部,而哺乳类动物则只在吸气是有新鲜空气进入肺部,呼气时只是在排除废气。正是由于这两点差异,导致鸟类呼吸系统较哺乳类动物更为高效。
鸟类的肺部结构和哺乳类动物的完全不同:鸟类肺部并没有如哺乳动物肺部中的肺泡,交换气体的功能落在了称为三级支气管(又叫旁支气管)的器官上。和哺乳类动物死胡同般的肺泡不同,三级支气管两端分别与次级支气管及背支气管相连,呈蜂窝管道状。三级支气管会辐射出许多肺毛细管,管壁上布满毛细血管[2]。连同与之相连的一个三级支气管所构成的六面棱柱体称为肺小叶,气体交换即发生在此。尽管鸟类肺部弹性纤维不多,不能如哺乳动物肺泡能膨胀,但其单位体重气体交换表面积却比哺乳类动物要高得多。前者为200cm2/g,而后者仅为15cm2/g[3]。
除此之外,鸟类不具有如同哺乳動物的横膈膜,因此只能够依靠移动肋骨膨胀胸腔的方式来吸入空气,然而這點在飛行時反而可以靠著飛行動作產生的負壓和擠壓來協助體內的空氣交換。而哺乳动物则可以不膨胀胸腔,仅依靠横膈膜的运动来吸入空气(腹式呼吸)。由于气囊存在于身体各处,因此在呼气的时候,不能如哺乳类动物般仅需依靠胸部肋骨自身重力来完成该动作,而需要全身各处相关肌肉的收缩来完成。
鸟类的发声器官位于气管底部,称为鸣管。与哺乳类动物的喉咙一样,气体通过发声器官会引起震动,因此发出声音。由于存在鸣管,某些鸟类可以制造比较复杂的声音,甚至模仿人类的说话,例如鹦鹉和八哥。有些鸣声悦耳动听的鸟类,甚至可以在同一时间发出超过1种声音。
循环系统
[编辑]和绝大多数哺乳类动物,以及某些爬行类动物(比如鳄鱼)相同,鸟类的心脏分为两房两室,不過肌肉更為強健發達。这种适应性进化,使得鸟类能够更有效率的向全身传递营养物质和氧气,以支撑飞行所需要的能量和新陈代谢速度。如红宝石喉蜂鸟,在飞行的时候,心脏的跳动频率可达每分钟1200次,或者说每秒跳动20下[4]。
消化系统
[编辑]
鸟类的喙与哺乳类动物的嘴有非常大的区别,因为内部容积通常较小,无法容纳类似哺乳类动物的牙齿,因此无法进行食物的研磨而只能拾取或撕裂食物。因为这一差别,导致其消化系统与哺乳类动物相比,有一定特殊之处。
许多鸟类在食道上存在一个肌肉构成的囊袋,称为嗉囊。嗉囊的作用是软化儲存其内的食物,以及通过儲存食物来调节进入下一个消化环节的食物量。不同鸟类的嗉囊大小和形状都很不一样。鸽形目如鸽子的嗉囊较为特别,可分泌出嗉囊乳(又称鸽乳),通过反刍这种营养物质来喂养雏鸽。
除此之外,鸟类还有一个砂囊(又称为胗、肫)。砂囊由四个肌肉群组成,通过这些肌肉群的运动,将食物在砂囊内来回挤压反动,并将其磨碎。在某些鸟类的砂囊中会发现沙石粒,这些可以辅助食物的研磨,其作用类似哺乳动物的牙齿。通过遗迹化石的研究发现,这种通过吞入沙石来辅助消化的方式也存在于恐龙当中,这种被恐龙所吞食的石头称为胃石。
饮水方式
[编辑]
鸟类饮水的方式通常可分为四种。大部分的鸟类的消化道不能像人一样可以蠕动,因此无法通过这种动作来吸或者说泵水到胃里面。因此大部分的鸟类都是通过不停的低头装水到喙里面,然后抬起头来让水通过重力的作用流到胃里面。这种方式通常被描述为“啜”或者“点”[5]。因此我们很少能看到有鸟能在水边一直俯身低头饮水,如豹子羚羊一般。然而也存在一些例外情况,例如鸽形目。康拉德·洛伦兹于1939年描述道:
尽管通常来说上面的说法大致上是正确的,然而从此之后,人们逐渐发现各种特例[5][7]。
此外,进化为专食花蜜的鸟类如太阳鸟和蜂鸟,则通过可伸缩的带槽的舌头来引水入消化道,而鹦鹉则是通过舔的方式来喝水[5]。
许多海鸟在眼睛附近有专门的腺体来处理海水中多余的盐分,这些盐分通过鼻孔排出。而许多生活在沙漠的鸟类则完全通过食物来获取所需的所有水分,同时通过仅仅排泄尿酸来降低对水的需求[8]。
骨骼系统
[编辑]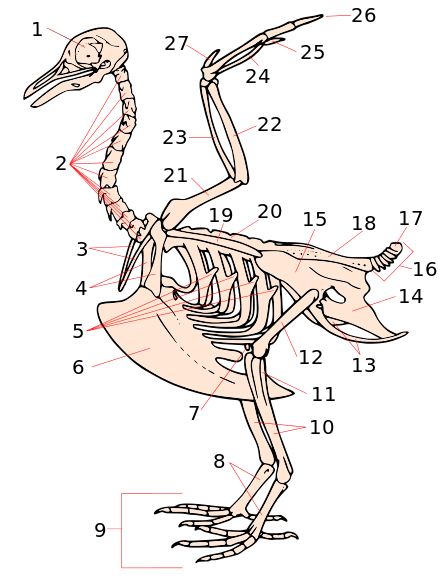
1. 颅骨
2. 颈椎
3. 叉骨
4. 鸟喙骨
5. 肋骨钩状突
6. 龙骨突
7. 髌骨
8. 跗跖骨
9. 趾骨
10. 胫骨 (胫跗骨)
11. 腓骨 (胫跗骨)
12. 股骨
13. 坐骨 (髋)
14. 耻骨 (髋)
15. 髂骨 (髋)
16. 尾椎
17. 尾综骨
18. 综荐骨
19. 肩胛骨
20. 腰椎
21. 肱骨
22. 尺骨
23. 桡骨
24. 腕骨
25. 掌骨
26. 指骨
27. 小翼羽
鸟类的骨骼进化为高度适应飞行:重量极轻而强度高,能够经受起飞至降落的整个飞行期间所产生的各种应力。其中一种适应性变化,是许多骨头会融合成单块融合骨(ossification),例如尾综骨。除此之外,鸟类也没有牙齿,甚至没有一个真正的下巴,而是进化出重量极轻的喙。正因为如此,鸟类的骨头数量比绝大多数陆生脊椎动物要少得多。许多鸟类在出生前就会在喙上长出一个称为破壳器的突出物,用于在孵化结束时在蛋壳上凿出一个洞。
鸟类的许多骨头都有空洞(气腔),中间纵横交错着可以加强结构强度的支撑柱和桁架。这种有空腔的骨头的数量在各种鸟之间不尽相同,那些可以滑翔和飞行的鸟类通常有更多这样的骨头。而这种骨头中的空腔,通常会存在由呼吸气囊所延伸出的气袋[1]。某些不会飞行的鸟类如企鹅以及鸵鸟没有这种空心的骨头,这更加证明了空心骨头是为了飞行而进化出来的[來源請求]。
鸟类颈椎的节数比很多脊柱动物要多,其中大多数由13-25块脊椎骨构成,因此脖子十分柔软灵活。鸟类是所有现存脊椎动物中唯一将锁骨融合成为叉骨,以及将胸骨融合成龙骨突的动物。用于飞行(或者如企鹅游泳)所需的胸肌非常巨大,因此需要龙骨突来提供一个更大的附着面。而那些不会飞的鸟类如鸵鸟等,其胸骨上的龙骨突并不发达。研究发现,游泳的鸟类胸骨通常较宽,飞行的鸟类长宽几乎相等,而只能在陆地上行走的鸟类,其胸骨相对较窄长[9]。
在鸟类的肋骨上,还有一种称为肋骨钩状突的结构。该结构的作用是通过覆盖后面一根肋骨,来增强整个肋骨框架的结构强度。喙头蜥也有类似的结构。和部分爬行类动物相似,鸟类的髋部较长,后肢有内跗关节。此外脊椎骨之间大量融合,胸带(锁骨、肩胛骨)愈合成上肢带骨也是一大特点。鸟类的头骨只有一个枕骨髁,此外与部分有前泪腺窝的爬行类动物一样,鸟类的头骨是双窝型的。[10]
骨骼
[编辑]

鸟类的颅骨主要由5个部分组成:顶部、枕部、面部、上喙和下喙。这几个部分均由多块骨愈合而成,由于在生长早期阶段就已开始愈合,因此已看不到骨缝。为了适应飞行,鸟类的颅骨变得非常轻,只占体重的约1%,其中部分位置还有气腔。
脊椎则可分为三大部分:颈部(13-16节),由背部至髋部等骨愈合而成的综荐骨,以及尾综骨。
胸腔正前方是叉骨和鸟喙骨,这两块骨和肩胛骨构成了上肢带骨。侧面是肋骨,肋骨在胸前与胸骨连接。
鸟的肩膀由肩胛骨、鸟喙骨以及肱骨(上臂)构成,肱骨则与尺骨和桡骨相连形成肘部。腕骨和掌骨形成了翅膀中“腕”和“掌”的部分,而指骨则互相融合成为一体。鸟类翅膀的骨头重量极轻,挥动翅膀时在自身重量上所作的功便相应减少了。
鸟的盆腔主要由三块骨头组成,分别是髂骨、坐骨 、耻骨,这三块骨头互相融合成開放性骨盆,明显演变为适合生蛋及孵化。在髋骨上有一对髋臼,下肢的第一块骨头股骨就与髋骨连接于此。 下肢的上段是股骨,在膝关节处与下方的胫跗骨相连,而胫跗骨的下端则于跗跖骨相连,趾骨则形成了爪。鸟类的脚趾形状可分为:不等趾型、对趾型、异趾型、并趾型以及前趾型等[11]。下肢骨头是鸟类最重的骨头,可使得鸟类的中心下移,以利于飞行。 鸟类所有的骨头加起来,其重量约为体重的5%,重量轻有利于飞行。
肌肉系统
[编辑]
大多数鸟类拥有约175组不同的肌肉,当中大部分用于控制翅膀、皮肤以及腿部。其中最大的是用于控制翅膀的胸大肌,在飞行鸟类中约占体重的15%至25%。在胸肌内部,有另一组称为喙上肌的肌肉。要能够飞行,则必须依赖这两组肌肉来挥动翅膀。其中喙上肌用于将翅膀升起,而胸大肌则用于将翅膀往下拍动。这两组肌肉加起来约占飞行鸟类体重的25%至35%。 鸟类皮肤上的肌肉则用于调整附着于其上的羽毛,以便帮助调整飞行姿态。 在躯干和尾部也有少量健壮且重要的肌肉,例如尾综骨上的肌肉可以控制尾部的姿态,这使得鸟类在飞行中能够迅速调整方向。
头部
[编辑]
相对于体重,鸟类大脑所占的比重较大。这导致了鸟类拥有相对较高及复杂的智力水平,因此鳥類可以做出複雜的行為和情感表現,例如築巢、育幼以及社交等等,馴養的鳥還可以學會人類的把戲。
鸟类的视觉非常敏锐,尤其是猛禽,可比人类锐利8倍。以鵟属鸟类为例,其视网膜上的视觉感受器密度为每平方毫米100万个,作为对比,人类则只有20万个,鳥的視網膜上微血管的分佈變到底下。此外,还有更多的视觉神经,使鳥類的色彩範圍也更大,可以看到紫外光範圍,人類相比之下相當於是色盲。以及其它动物不曾被发现的第二套眼部肌肉。甚至在某些品种下存在複數称为中央窝的结构,以扩大该区域的视野,就如配置望遠鏡一般。包括蜂鸟和信天翁科在内的许多鸟类,在每一只眼球内都具有两个中央窝,一個看近,一個看遠。许多鸟类可以侦测出光的偏振。鸟头骨上有一个有许多小骨头组成的围绕眼睛的环状结构,称之为巩膜眼环,眼球便容纳其中。这种特征也见于爬行类动物。還有鳥類眼中有油酯成分,與太陽眼鏡相似,可以提高在濃霧和下雨中的能見度。
许多水禽的喙部存有赫氏小体——一种类似帕西尼氏小体的机械感受器,用以感知躲藏在湿润沙砾底下的猎物对水体所造成的微小压差[12]。对于现存的所有鸟类来说,其上喙骨均可相对头盖骨进行移动。其中某些鸟类尤其明显,像鹦鹉就可以轻易被观测到这一运动[13]。
头部两侧位于眼睛和喙部之间的区域,称为眼端,這部分有一定程度上具備穿透的感光能力。该区域有时候是没有羽毛的,该处的皮肤甚至可能有鲜艳的颜色,如鸬鹚科中的许多品种便是如此。
繁殖
[编辑]
尽管大多数雄性鸟类并没有外部性器官,然而确实拥有两颗睾丸,并且在繁殖季节时会增大几百倍以生产精子[14]。雄性鸟类的两颗睾丸通常是不对称的,其左边的那颗会大得多[15]。雌性鸟类的卵巢也有类似的现象:在繁殖季节增大,并且左边的才能发挥正常功能。然而,如果左边的卵巢若是出问题了,右边的卵巢则会尝试正常发挥其功能。然而有时候右卵巢会发育成卵睾体或者睾丸,并因此发生性逆转现象[3]。
一些鸟类的雄性並無阴茎,而在交配前其精液存储于泄殖腔乳突内的储精囊内。在交配的时候,雌鸟尾部偏向一旁,而雄性或从后方,或从前方(如缝合吸蜜鸟)骑与其上,或以其它方式接近,最后泄殖腔互相接合,使得精液得以进入雌鸟的生殖道中。这一过程发生的非常快,甚至在半秒之内即可完成。交配后精液存储在雌鸟被称为精小窝的阴道腺的小管中,存储的时间根据物种的不同,从一周到一年不等。卵在离开卵巢之后会被分别受精,然后再变成蛋产于体外,并继续在雌鸟体外孵化发育。

许多水禽以及如鸵鸟、火鸡等鸟类的雄性个体拥有阴茎,该器官并非哺乳动物阴茎的同源器官。在平时,该器官收缩到肛道壁外的囊中。[3]
当蛋被孵化之后,雏鸟的父母会提供不同程度的照顾,例如提供食物和保护等。早熟鸟在孵化后的几分钟内就可以独立行动并照顾自己,而晚熟鸟则尚未睁眼、无毛且无法照顾自己,需要父母的更多照料。许多在地面筑巢的鸟,如山鹑和涉禽,在孵化后幾乎立刻就能够四处跑动,这类的鸟被称为离巢性鸟。与之相反的,在洞穴中筑巢的鸟,其雏鸟通常无法在缺乏照料的状态下成活。这样的鸟需要经历换羽的过程,并学会飞行后才能独立。
某些鸟如鸽子、鹅以及丹顶鹤等,会在一生中拥有固定的伴侣,并定期生育抚养后代,而其他鸟类则会在繁殖期间重新选择伴侣。
鳞
[编辑]鸟类在脚趾和跗跖骨上会有鳞,甚至某些鸟类在踝关节上也有鳞。这些鳞和喙、爪、距等类似,由角蛋白构成。除了翠鸟和啄木鸟之外,其它鸟的鳞并不明显。鸟鳞被认为与爬行类动物及哺乳类动物的鳞甲是同源器官[16]。
鸟类在胚胎时期皮肤是光滑的,但脚部皮肤会角质化并形成角质层(皮肤最外层),这些变厚的部分就形成了鳞。这些鳞的形式可分为:
- 松皮状:这种小片的鳞,其实只是比较厚的皮肤,上面有纵横交错的浅沟而已;
- 蜂窝状:细小但互相间不重叠的鳞,可见于鸡爪胫跖关节的正面和侧面,由α-角蛋白构成[17];
- 盾状:比板状的稍小,可见于鸡爪胫跖关节的后面;
- 板状:最大的鳞,位于跗跖骨、胫跗骨的后面,以及趾骨的上面。和爬行类动物一样,这种鳞由β-角蛋白构成[17]。
在跗跖骨正前方的一列鳞片,又被称为跗跖顶部(acrometatarsium或acrotarsium)。某些鸟腿部的羽毛和鳞片发生混杂,羽毛囊可存在于鳞片之间,甚至在鳞片之下深处的真皮层中,其中后者羽毛会直接穿过角质构成的鳞片。[16]
注释
[编辑]参考文献
[编辑]引用
[编辑]- ^ 1.0 1.1 Ritchison, Gary. Ornithology (Bio 554/754):Bird Respiratory System. Eastern Kentucky University. [2007-06-27]. (原始内容存档于2007-06-27).
- ^ Bird lungs. [2011-05-27]. (原始内容存档于2007-03-11).
- ^ 3.0 3.1 3.2 第十五章 家禽解剖. 南京农业大学. [2011-03-01].[永久失效連結]
- ^ June Osborne. The Ruby-Throated Hummingbird. University of Texas Press. 1998: 14. ISBN 0-292-76047-7.
- ^ 5.0 5.1 5.2 "Drinking Behavior of Mousebirds in the Namib Desert, Southern Africa "; Tom J. Cade and Lewis I. Greenwald; The Auk, V.83, No. 1, January, 1966pdf (页面存档备份,存于互联网档案馆)
- ^ K. Lorenz, Verhandl. Deutsch. Zool. Ges., 41 [Zool. Anz. Suppl. 12]: 69-102, 1939
- ^ "Drinking Behavior of Sandgrouse in the Namib and Kalahari Deserts, Africa"; Tom J. Cade, Ernest J. Willoughby, and Gordon L. Maclean; The Auk, V.83, No. 1, January, 1966pdf (页面存档备份,存于互联网档案馆)
- ^ Gordon L. Maclean (1996) The Ecophysiology of Desert Birds. Springer. ISBN 3540592695
- ^ Ayhan Duezler, Ozcan Ozgel, Nejdet Dursun (2006) Morphometric Analysis of the Sternum in Avian Species. Turk. J. Vet. Anim. Sci. 30:311-314
- ^ Wing, Leonard W. (1956) Natural History of Birds. The Ronald Press Company. [1]
- ^ Proctor, N.S. and P.J. Lynch. Manual of ornithology: avian structure and function. Yale Univ. Press, New Haven. 1993.
- ^ Piersma, Theunis; Renee van Aelst, Karin Kurk, Herman Berkhoudt and Leo R. M. Maas. A New Pressure Sensory Mechanism for Prey Detection in Birds: The Use of Principles of Seabed Dynamics?. Proceedings: Biological Sciences. 1998, 265 (1404): 1377–1383. doi:10.1098/rspb.1998.0445.
- ^ Zusi, R L. A Functional and Evolutionary Analysis of Rhynchokinesis in Birds.. Smithsonian Contributions to Zoology. 1984, 395.
- ^ A study of the seasonal changes in avian testes (页面存档备份,存于互联网档案馆) Alexander Watson, J. Physiol. 1919;53;86-91, 'greenfinch (Carduelis chloris)', 'In early summer (May and June) they are as big as a whole pea and in early winter (November) they are no bigger than a pin head'
- ^ Lake, PE. Male genital organs. King AS, McLelland J (编). Form and function in birds 2. New York: Academic. 1981: 1–61.
- ^ 16.0 16.1 Lucas, Alfred M. Avian Anatomy - integument. East Lansing, Michigan, USA: USDA Avian Anatomy Project, Michigan State University. 1972: 67, 344, 394–601.
- ^ 17.0 17.1 Peter R. Stettenheim (2000) The Integumentary Morphology of Modern Birds—An Overview. American Zoologist 2000 40(4):461-477; doi:10.1093/icb/40.4.461
来源
[编辑]- Peter Grant & Killian Mullarny. The New Approach to Identification, in Birding World, Vols. 1&2 ISSN 0969-6024