Genetik
| Denna artikel ingår i en artikelserie |
| Genetik |
|---|
 |
| Nyckelämnen |
| Kromosom · DNA · RNA · Genom · Ärftlighet · Mutation · Nukleotid |
| Relaterade ämnen |
| Evolution · Populationsgenetik · Kvantitativ genetik · Molekylär genetik |
| Forskning och tillämpningar |
| Genetiker · DNA-sekvensering · Genteknik |
|
|


Genetik, ärftlighetslära, är en vetenskap inom biologin som studerar hur egenskaper nedärvs, hur genomet (arvsmassan) är uppbyggt och fungerar, hur förändringar av generna (arvsanlagen) uppstår, samt den biologiska variationen. Alltsedan förhistorisk tid har människorna förbättrat husdjur och odlade växter genom att använda indirekt kunskap om hur egenskaper ärvs från föräldrar till avkomman för att genomföra mer eller mindre systematisk avel. Men det vetenskapliga studiet av de genetiska mekanismerna tog sin början först med Gregor Mendel vid mitten av 1800-talet. Mendel kände inte heller till de grundläggande molekylära genetiska mekanismerna. Men genom systematiska kontrollerade experiment och noggrann statistik kunde han klarlägga generella principer för hur nedärvningen går till, till exempel att det som ärvs är ett antal enskilda särdrag, arvsanlag, som ärvs oberoende av varandra, det som senare kom att kallas gener.
Genetiken tillhandahåller viktiga verktyg som används i den moderna forskningen för att undersöka funktionen hos specifika gener, till exempel genom kartläggning av genetisk interaktion. I organismerna är den genetiska informationen vanligtvis lagrad i den kemiska strukturen hos specifika DNA-molekyler, som i sin tur finns i kromosomer. Arvsanlagen finns i oerhört långa molekyler, DNA, som vardera består av en kedja av nukleotider. Dessa kedjor sitter ihop parvis och bildar något som ser ut som en spiralskruvad stege. Nukleotiderna sticker ut åt sidan från vardera kedjan. I DNA används fyra olika nukleotider. De är olika långa, men matchar varandra kort mot lång, så att varje steg i stegen blir ungefär lika långt. Den ordningsföljd som nukleotiderna har i kedjan utgör den genetiska informationen. Det här är lite likt hur ordningsföljden mellan bokstäverna på en boksida gör att vi kan utläsa ett meddelande.
Generna motsvarar delsträckor längs kedjan. Sekvensen av nukleotider i genen översätts i två steg till en sekvens av aminosyror som bildar ett protein. En grupp om tre nukleotider motsvarar en aminosyra. Denna motsvarighet kallas den genetiska koden. Aminosyrasekvensen avgör vilken form proteinet får och därmed vilken funktion det kan fylla. I stort sett alla uppgifter som behöver utföras i den levande cellen sköts av proteiner.
Genetiken har stor betydelse för vilken yttre form och vilket beteende en organism ska få. Men den är inte allenarådande. Miljön som organismen lever i påverkar också hur den blir. Slutresultatet formas av ett samspel mellan arv och miljö. Som exempel bestäms den längd ett människobarn uppnår i vuxen ålder både av de gener det får av sina föräldrar och av hur mycket näring det får, vilken näring det får, vilka sjukdomar det råkar ut för och hur mycket det motionerar.
Molekyler och grundläggande kemiska processer
[redigera | redigera wikitext]DNA och kromosomer
[redigera | redigera wikitext]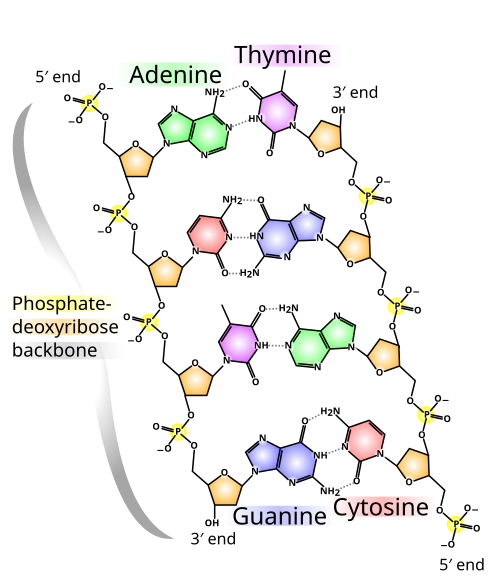
Den genetiska informationen och generna finns i oerhört långa molekyler som kallas DNA, deoxyribonukleinsyra. DNA är kedjor av nukleotider. En nukleotid består av en fosfat, en socker och en kvävebas. Det finns fyra olika kvävebaser: adenin (A), cytosin (C), guanin (G) och tymin (T). Den genetiska informationen utgörs av den ordning (sekvens) i vilken nukleotiderna placerats längs kedjan. Varje gen är en liten delsträcka i en stor DNA-molekyl.[1] (Vissa virus utgör ett undantag från denna princip — i dessa virus lagras den genetiska informationen i RNA, en molekyl som är mycket lik DNA.[2]
DNA förekommer normalt i par om två kedjor som löper parallellt och tillsammans följer en korkskruvsform, en dubbel helix. När DNA-kedjorna bildar par kommer varje nukleotid mitt för en nukleotid i den andra kedjan. Nukleotiderna passar då olika bra ihop, så att bindningen blir starkast om A paras ihop med T och C paras ihop med G. I naturligt DNA har sammanlänkningen nästan alltid gjorts på detta optimala sätt. Genom detta förhållande finns hela sekvens-informationen i båda delkedjorna. Om man vet vilken sekvens som finns i den ena kedjan, kan man från denna räkna ut exakt vilken sekvens den andra kedjan har. Detta förhållande utgör den kemiska grunden för nedärvning: vid DNA-replikering kopieras den genetiska informationen genom att dubbelkedjan delas upp och genom att varje delkedja används som mall vid skapandet av en ny par-kedja.[3]
Den långa dubbla DNA-kedjan kallas också kromosom. En kromosom innehåller normalt ett stort antal gener. Generna ligger då i en viss ordning längs kedjan, med större eller mindre mellanliggande nukleotidsekvenser som inte ingår i gener. En bakterie innehåller normalt en enda kromosom som är formad som en ring, kedjan har inga ändpunkter. Eukaryota celler har nästan alltid flera kromosomer. Dessa har ändpunkter. Människocellens längsta kromosom är 247 miljoner baspar lång.[4] Det finns ett stort antal proteiner som binder sig till DNA-kedjan och bidrar till att hålla ordning på den långa kedjan, ge den en hanterbar fysisk form och kontrollera vilka andra molekyler som kan bearbeta den. DNA-kedjan och proteinerna bildar tillsammans materialet kromatin.[5] Den fullständiga uppsättningen genetiskt material i en organism (vanligtvis den sammanlagda sekvensen av alla dess kromosomer) kallas organismens genom.
I många organismer förekommer kromosomerna i analoga par, det vill säga att för nästan varje dubbelkedja finns en annan dubbelkedja med nästan samma innehåll. Sådana organismer sägs vara diploida.[6] Organismer som inte har sådana kromosompar sägs vara haploida. De två allelerna för en gen är lokaliserade på analoga positioner på var sin analog kromosom. Den ena allelen har ärvts från den ena föräldern och den andra allelen från den andra föräldern.

Könskromosomerna utgör ett undantag genom att dessa könsbestämmande kromosomer ofta är mycket olika. Men de förekommer ändå oftast i par.[7] Hos människan och många andra djur finns en så kallad Y-kromosom som är mycket liten och innehåller mycket få gener. Denna kromosom innehåller den gen som sätter igång de processer som utvecklar de specifikt manliga egenskaperna. Y-kromosomen paras ihop med en så kallad X-kromosom. Eftersom Y-kromosomen gör att individen blir en man förekommer den endast i män. Män har alltså en X-kromosom och en Y-kromosom, medan kvinnor har två X-kromosomer. X-kromosomen är hos människan en av de största kromosomerna.
Reproduktion
[redigera | redigera wikitext]När celler delar sig kopieras hela genomet och vanligtvis erhåller varje dottercell en kopia. Celldelning förekommer i två huvudvarianter. Mitosen är den enklaste och vanligaste formen av reproduktion. Den utgör grunden för asexuell reproduktion. Asexuell reproduktion förekommer också hos flercelliga organismer. Avkomman får då hela sitt genom från en enda förälder. Avkomma som är genetiskt identisk med sin förälder kallas klon.
För eukaryota organismer är sexuell reproduktion vanligast. Avkomman får då en kombination av genetiskt material från två föräldrar. Vid sexuell reproduktion alternerar cellerna mellan en form som innehåller en enkel kromosomuppsättning (haploid) och en form med dubbel kromosomuppsättning (diploid).[6] Haploida celler kan smälta samman och kombinera sina genetiska material så att det bildas en diploid cell. Haploida celler uppstår genom att diploida celler delar sig utan att DNA kopieras. Dottercellerna erhåller då en slumpvis vald kromosom från varje kromosompar. De flesta djurarterna och många växter är diploida under större delen av livscykeln och den haploida fasen är för dessa begränsad till en enda cell, gamet, till exempel en spermie eller en äggcell.
Bakterier har inte sexuell reproduktion med övergångar mellan haploida och diploida faser. Men de har andra sätt att skapa nya kombinationer av genetiskt material. Genom konjugation kan de till exempel överföra små ringformade DNA-kedjor mellan sig.[8]
Rekombination och genetisk koppling
[redigera | redigera wikitext]
Vid sexuell reproduktion kan gener som befinner sig på olika kromosomer i ett kromosompar fördela sig oberoende av varandra i avkomman. På detta sätt kan nya genkombinationer uppstå. Gener på samma kromosom kan i princip inte omkombineras. Men genom överkorsning kan gener flyttas mellan kromosomer. Vid överkorsning utbyts en delsträcka av DNA-kedjan mellan två kromosomer, så att två analoga avsnitt byter plats.[9] Denna process förekommer framför allt under reduktionsdelningen eller meios, den specialiserade typ av celldelning som skapar haploida celler (gameter).
Sannolikheten för att överkorsning ska inträffa mellan ett visst par av positioner på kromosomen beror på avståndet mellan punkterna. För gener på långt avstånd från varandra är sannolikheten för överkorsning så stor att generna blir lika lite kopplade till varandra som två gener på olika kromosomer. Men för gener som befinner sig närmare varandra finns en betydande sannolikhet att de ska följa varandra, att de ska vara genetiskt kopplade till varandra och ärvas tillsammans. Genom att studera dessa sannolikheter för att gener ska följas åt kan man göra en karta som visar ungefär var längs kromosomen respektive gen befinner sig.[10]
Arvsmassan verkar genom att styra hur proteinerna blir
[redigera | redigera wikitext]Introduktion
[redigera | redigera wikitext]Generna spelar sin huvudsakliga roll när cellen tillverkar proteiner. Genen innehåller den information som avgör hur proteinerna ska bli. Cellerna är de allra minsta självständiga delarna i organismer.
Den principiella arbetsuppdelningen i celler är enkel - generna innehåller instruktionerna för arbetet och proteinerna utför arbetet. Uppgifterna kan till exempel bestå i att bygga en ytterligare kopia av cellen eller att reparera skador i den. Varje proteintyp är en utpräglad specialist och utför endast ett visst speciellt jobb. Generna för alla proteinerna finns alltid tillgängliga.
Protein består av långa kedjor av aminosyror. En aminosyra i taget läggs till i ena änden. Varje gång en aminosyra ska läggas till finns 20 olika att välja mellan. Det är genen som avgör vilken aminosyra som läggs till. Ordningsföljden mellan nukleotiderna i DNA bestämmer ordningsföljden mellan aminosyrorna i proteinet. Den tredimensionella form som proteinet då får bestäms av den ordningsföljd som aminosyrorna sitter i längs kedjan och det är denna form som i sin tur bestämmer vad proteinet kan göra, vad det har för funktion.

Om sekvensen av nukleotider förändras så förändras också genen. Detta kan få till följd att den motsvarande sekvensen av aminosyror i proteinet blir annorlunda. Om en del av genen tas bort blir proteinet kortare och kanske inte fungerar. Detta är förklaringen till att organismer med olika alleler kan ha olika egenskaper, motsvarande proteiner fungerar olika. Som exempel beror hårfärgen på hur mycket melanin (mörkt pigment) som byggs in i håret när det växer fram. En person med normal uppsättning gener för tillverkning av melanin får mörkt hår. Men om en av allelerna har en sekvens som är så annorlunda att motsvarande protein inte fungerar, då produceras mycket mindre melanin, och personen blir ljushårig.
Den genetiska koden
[redigera | redigera wikitext]
Genernas funktion är vanligtvis att de bestämmer utseendet för ett protein som produceras. Proteiner är kedjor av aminosyror och sekvensen av nukleotider i genens DNA används för att bestämma sekvensen av aminosyror i proteinet. Vägen från DNA börjar med att cellen tillverkar en RNA-molekyl som motsvarar genens DNA-sekvens. Denna process kallas transkription. RNA som har denna roll kallas budbärar-RNA.
RNA-molekylen används sedan som mall vid tillverkning av en kedja av aminosyror. Denna process kallas translation, det engelska ordet för översättning. Vid translationen används en genetisk kod som pekar ut en aminosyra för en grupp om tre nukleotider. En sådan triplett kallas kodon.[11] Flödet av information går åt ett håll, från nukleotid-sekvens i DNA till aminosyra-sekvens i protein. Francis Crick kallade detta för molekylärbiologins centrala dogm.[12]
Aminosyrornas ordningsföljd avgör vilken tredimensionell form proteinet får när kedjan vecklar ihop sig. Denna form är i sin tur avgörande för den funktion proteinet kan ha i cellens processer.[13][14] En del, så kallade strukturella proteiner, bildar stommen i cellen. Det kan vara fibrer som kollagen. Andra proteiner kan binda till enkla molekyler och andra proteiner, ofta som enzymer som underlättar kemiska reaktioner mellan de molekyler som proteinet bundit till (utan att proteinet självt förändras). Proteinets struktur är dynamisk; proteinet hemoglobin böjs till något olika former vilket underlättar infångandet, transporten och frigörandet av syre-molekyler i däggdjurs-blod.
Utbyte av en enda nukleotid i DNA kan medföra att en aminosyra i motsvarande protein byts ut. Eftersom proteinets struktur är avhängig av aminosyrasekvensen kan ett sådant utbyte få dramatiska följder för proteinets egenskaper genom att strukturen blir instabil eller genom att ytan förändras så att proteinets interaktion med andra proteiner eller molekyler förändras. Ett exempel är sickelcellsanemi som är en genetisk sjukdom som uppkommer på grund av att en enda nukleotid bytts ut i sekvensen som kodar för β-globin i hemoglobin. Detta gör att en aminosyra blir utbytt och hemoglobinet får andra fysiska egenskaper.[15] Sickle-cell-versioner av hemoglobin fastnar i varandra så att de staplas på varandra och bildar långa fibrer som förstör de röda blodkropparnas form. Dessa celler som får formen av en skära (engelska sickle) har därför mycket svårare att ta sig fram genom de minsta blodkärlen, utan tenderar att klumpa ihop sig och förstöras, och därmed orsaka de kända symptomen för denna sjukdom.
Vissa gener kopieras till RNA utan att detta RNA i sin tur blir använt som mall för proteinsyntes. Sådana RNA-molekyler kallas icke-kodande RNA. En del av dessa RNC vecklas ihop till former som bidrar till viktiga processer is cellen. De ingår till exempel ribosomer och fungerar som transfer-RNA. RNA kan också ha reglerande funktioner genom interaktion med andra RNA-molekyler genom hybridisering.
Genreglering
[redigera | redigera wikitext]En organisms genom innehåller tusentals gener. Men alla generna behöver inte vara aktiva samtidigt, så cellen producerar vid varje tillfälle endast mRNA och proteiner bara från ett urval av sina gener. I cellen finns många processer som reglerar vilka gener som är aktiva och hur mycket protein som tillverkas från dem. Transkriptionsfaktorer är reglerande proteiner som binder till tidiga delar av den DNA-del-sekvens som utgör genen.[16] Transkriptionsfaktorn möjliggör eller stimulerar transkription av cellen.
I genomet för bakterien Escherichia coli finns till exempel en serie gener som är nödvändiga för produktion av aminosyran tryptofan. Men om cellen har tillräcklig mängd tryptofan så behövs inte dessa geners proteiner. Närvaron av troptofan påverkar därför dessa geners aktivitet genom att tryptofan-molekyler binds till ett protein som kallas tryptofan-repressor (en transkriptions-faktor). Detta ändrar tryptofan-repressorns form så att den binder till genen. Tryptofan-repressorn blockerar då transkription av generna och därmed produktionen av motsvarande proteiner.[17] Detta är ett exempel på negativ återkoppling.

Skillnader i genuttryck är särskilt tydliga i flercelliga organismer, där alla celler innehåller samma genom men ändå har mycket olika struktur och beteende, beroende på att de uttrycker olika delmängder av generna. Alla cellerna i en flercellig organism härstammar från en enda cell. De förändras till olika gentyper som ett svar på olika inverkan från omvärlden och olika signaler från andra celler och gradvis etablerar de olika genuttrycksmönster. Det finns inte en ensam gen som är orsak till utvecklingen av olika strukturer i den flercelliga organismen. Dessa mönster uppkommer genom komplex interaktion mellan många celler.
I eukaryota celler finns kromatin-strukturer som påverkar transkriptionen av gener. Dessa består ofta av modifieringar av DNA och kromatin som kan ärvas till dotterceller på ett stabilt sätt.[18] Dessa strukturer sägs vara epigenetiska, eftersom de tillkommer utöver och genom modifiering av den information som finns i DNA-sekvensen och eftersom de överför information från en cell till nästa generation. Epigenetiska strukturer kan göra att olika celltyper som odlas i samma medium kan ha mycket olika egenskaper. Trots att epigenetiska strukturer i allmänhet är dynamiska under utvecklingen kan en del, till exempel genom paramutation, ärvas över flera generationer och utgöra mycket ovanliga undantag till principen att arvet förmedlas genom överföring av DNA.[19]
Arvets och miljöns relativa inflytande
[redigera | redigera wikitext]
Trots att generna innehåller all den information en organism använder för att fungera spelar också miljön en viktig roll för hur fenotypen blir. Organismens fenotyp beror på ett samspel mellan genetiken och miljön. Ett exempel på detta är temperatur-känsliga mutationer. Ofta kan utbytet av en enda aminosyra göra att ett protein blir mindre stabil till sin form, utan att dess beteende eller interaktion med andra molekyler förändras. Vid hög temperatur, när molekylerna rör sig snabbare och kolliderar hårdare, kan ett sådant protein förändras till sin form i så hög grad att det förlorar sin funktion. Samma protein kan alltså fungera normalt vid lägre temperaturer. En sådan mutation gör att siamesiska katter har pigment på kroppsdelar som sticker ut och kyls mer, men inte på bålen, där den är varmare.[20]
Miljön har också en avgörande betydelse för den genetiska sjukdomen fenylketonuri.[21] Den mutation som orsakar fenylketonuri förstör kroppens förmåga att bryta ner aminosyran fenylalanin. Detta gör att giftiga halter byggs upp av en molekyl som utgör ett mellansteg i den normala nedbrytningen. Denna molekyl orsakar kramper och allvarliga försämringar av de mentala funktionerna. Men om en person som har mutationen för fenylketonuri följer en strikt diet som inte alls innehåller fenylalanin så fungerar denne normalt och är frisk.
En populär metod att avgöra arvets respektive miljöns relativa inflytande är att studera hur enäggstvillingar skiljer sig åt i egenskaper och jämföra det med vilka skillnader som finns i andra tvillingpar. Vetenskapsmännen jämför då hur ofta enäggstvillingar har samma brist med hur ofta andra tvillingpar har samma brist.
Nedärvning
[redigera | redigera wikitext]Autosomal nedärvning sker oberoende av kön. Förändringen sitter inte i könskromosomen.
Intermediär nedärvning är när ingen av generna i ett genpar dominerar över den andra utan båda bidrar till organismens fenotyp.
Polygen nedärvning innebär att många gener tillsammans ger en egenskap, i lägre eller högre grad. Varje gen var för sig, ger en ganska liten eller ingen effekt alls, men som tillsammans ger stor effekt.
Nedärvning och Mendels lagar
[redigera | redigera wikitext]
På den mest grundläggande nivån sker nedärvning i organismer i diskreta särdrag, så kallade gener.[22] Denna egenskap hos nedärvning observerades först av Gregor Mendel, som studerade uppdelning av ärftliga egenskaper hos ärtplantor.[23][24] I de experiment där han studerade egenskapen blomfärg, observerade Mendel att blommorna på en viss planta var antingen lila eller vit, men aldrig hade någon mellanliggande färg. Dessa olika och klart åtskilda versioner av samma gen kallas alleler.
För ärtan, som är diploid, har varje plant-individ två alleler av varje gen, en från varje förälder.[6] Många arter, däribland människan, har detta nedärvningsmönster. Diploida organismer med två kopior av samma allel för en viss gen sägs vara homozygota för denna gen, vid detta locus, det vill säga på denna lilla delsekvens av DNA-kedjan. Individer som har två olika alleler av en viss gen sägs vara heterozygota för denna gen.
Hela uppsättningen alleler hos en individ kallas dess genotyp, medan alla dess observerbara egenskaper (särdrag) kallas dess fenotyp. När en individ är heterozygot för en gen är det ofta så att den ena allelens egenskaper är helt bestämmande för fenotypen. Denna allel sägs vara dominant. Den andra allelen, som inte ger någon observerbar egenskap, sägs vara recessiv. En del alleler har mellanliggande effekter.[25]
När två individer reproducerar sig sexuellt, ärver avkomman slumpvis en av de två allelerna från respektive förälder. Dessa observationer av diskret nedärvning och uppdelning (segregation) av alleler är tillsammans kända som Mendels första lag.
De grundläggande lagarna inom klassisk mendelsk genetik innefattar nedärvningsmönstren:
Autosomal dominant
Autosomal recessiv
X-länkad dominant
X-länkad recessiv
Mitokondriell nedärvning[26]
Diagram och speciella notationssätt
[redigera | redigera wikitext]
Genetiker använder diagram och symboler för att beskriva nedärvning. En gen representeras av en eller flera bokstäver. Ofta används ett plus-tecken ('+') för den vanliga o-muterade allelen av en gen, den så kallade vild-typen.[27]
Vid experiment med befruktning och avel, särskilt när man diskuterar Mendels lagar, representeras föräldragenerationen ofta med "P" och första generationen avkomma med "F1". Avkomman av generation F1 kallas i sin tur "F2". Punnet-diagrammet används ofta för beräkning av förväntat resultat av kors-avel.
När genetiker studerar genetiska sjukdomar hos människan använder de ofta stamtavlor för att klargöra nedärvningen av särdrag.[28] Stamtavlan visar nedärvning av särdrag i ett familjeträd.
Samspel mellan flera gener
[redigera | redigera wikitext]
Organismer har tusentals gener, och för organismer med sexuell reproduktion kan generna normalt omkombineras fritt. Detta innebär att ärvandet av en allel för gul eller grön ärta inte är beroende av ärvandet av alleler för vit eller violett blomma. Så alleler för olika gener bildar andra kombinationer i avkomman än hos föräldrarna och ett oerhört stort antal kombinationer kan bildas. Undantaget att vissa par av gener tenderar att följas åt kallas genetisk koppling.
I många fall samverkar flera olika gener på ett sätt som påverkar samma särdrag. Hos växten Omphalodes verna finns till exempel en gen med alleler som bestämmer blommornas färg: blå eller magenta. Men en annan gen kontrollerar om blommorna blir vita eller får färg. När en individ har två kopior av den vita allelen blir dess blommor vita oavsett om den första genen innebär blå eller magenta. Ett sådant samspel kallas epistasis. I detta fall är den andra genene (vit eller färg) är epistatisk relativt den första.[29]
Många särdrag varierar på ett i stort sett kontinuerligt sätt, till exempel människors längd och hudfärg. Här är det inte som med O. verna, där varje blomma har en av endast tre olika färger. De kontinuerligt varierande egenskaperna är resultatet av ett samspel av många olika gener.[30] Dessa geners inflytande förmedlas i varierande grad av den miljö som organismen upplever. Hur mycket individens egenskaper beror på genetiskt arv kallas arvbarhet.[31] Arvbarheten kan uttryckas med en siffra mellan 0 och 1 (eller 0 och 100 %) där 1 betyder att en viss egenskap enbart beror på genotypen och inte alls på miljön. Den högsta arvbarhet man funnit är 0,97. Det gäller för fingeravtryck hos enäggstvillingar. Kroppsmått har ofta en hög grad av arvbarhet, medan livslängd och antal avkomma har låg arvbarhet. Mätning av arvbarhet är relativ. I en mer varierande miljö har miljön större påverkan på den totala variationen av egenskapen. Som exempel är det en komplex process som bestämmer hur långa människor blir. I USA har denna egenskap en arvbarhet på 89 %. Men i Nigeria, där tillgången på näringsriktig mat varierar mer, är arvbarheten för längd bara 62 %.[32]
Evolution
[redigera | redigera wikitext]Mutationer
[redigera | redigera wikitext]
Under DNA-replikering inträffar ibland fel i polymeriseringen av den nya kedjan. Dessa fel kallas mutationer och kan påverka organismens fenotyp, särskilt om de inträffar i en gens kodande sekvens. Felfrekvensen är oftast mycket låg, ett fel per 10–100 miljoner baspar. Felen hålls tillbaka av den korrekturläsningsfunktion som ingår i DNA-polymeras.[33][34] Utan korrekturläsningsfunktionen skulle felfrekvensen vara tusenfalt högre. Eftersom många virus använder sig av DNA och RNA-polymeras som saknar korrekturläsningsfunktion har de den högre mutationsfrekvensen. Processer som ökar frekvensen av DNA-förändringar sägs vara mutagena. Mutagena kemikalier stimulerar felkopiering vid DNA-replikering, ofta genom att störa sammanparandet av baspar. UV-strålning introducerar mutationer genom att direkt förändra DNA-strukturen.[35] Kemiska skador på DNA kan också uppkomma utan externa störningar. Celler använder mekanismer för DNA-reparation för att reparera skillnader och avbrott i DNA-kedjan. Men reparerandet misslyckas då och då med att återställa den ursprungliga sekvensen.
Naturligt urval
[redigera | redigera wikitext]Mutationer förändrar organismernas genotyp och en del av dessa förändringar påverkar också fenotypen. De flesta mutationerna har obetydlig inverkan på fenotypen, hälsan eller förmågan till reproduktion. De mutationer som har inverkan påverkar oftast till det sämre, men ibland innebär mutationen en förbättring. Studier av bananflugan Drosophila melanogaster visar att av de mutationer som ändrar det protein som genen kodar för är 70% skadliga medan resten är neutrala eller svagt fördelaktiga.[36]

Populationsgenetik studerar hur genetiska skillnader fördelas i populationer och hur fördelningarna förändras över tiden.[37] Förändringar i allel-frekvenser i en population påverkas framför allt av naturligt urval, om en allel ger en reproduktionsfördel för organismen,[38] men också av andra processer, som genetisk drift, artificiellt urval och genflöde.[39]
När dessa processer verkar över många generationer så förändras organismernas genom på ett signifikant sätt. Det är dessa förändringar som är evolutionen. Urval av motationer som är fördelaktiga kan göra att arter utvecklas till former som har bättre förmåga att överleva i artens miljö. Detta kallas anpassning.[40] Artbildning, utvecklande av nya arter, inträffar oftast genom att populationer blir geografiskt isolerade från varandra så att de inte kan utbyta gener.[41] Den vetenskapsrörelse som framför allt på 40-talet tillämpade genetiska principer för att studera populations-biologi och evolution kallas den Moderna syntesen.
Genom att jämföra olika arters motsvarande, homologa, DNA-avsnitt kan man uppskatta det evolutionära avståndet mellan dem, det vill säga när de blev olika arter. Detta kallas den molekylära klockan.[42] Evolutionära avstånd mellan arter kan användas för att konstruera ett evolutionärt träd. Dessa träd visar det gemensamma ursprunget och hur arterna har utskiljt sig från varandra över tiden. Men de visar inte vilket genetiskt material som överförts mellan obesläktade arter. Sådan överföring kallas horisontell genöverföring och är vanligast bland bakterier.
Forskning och utveckling
[redigera | redigera wikitext]Modellorganismer
[redigera | redigera wikitext]
I början studerade genetikerna nedärvning i ett stort antal organismer. Men med tiden kom de att specialisera sig på ett ganska litet antal. När det redan fanns en betydande mängd forskningsresultat för en viss organism blev detta en motivation för andra forskare att välja samma organism för vidare undersökningar.[43] Bland de vanliga ämnena för genetiska undersökningar av modellorganismer ingår genreglering och genernas inverkan på utveckling och cancer.
I viss mån valdes modellorganismer för att de är praktiska att arbeta med, till exempel för att de har korta generationscykler eller är lätta att utföra genmanipulering på. Förutom bananflugan ingår bland de mycket brett använda modellorganismerna tarmbakterien Escherichia coli, den lilla växten backtrav, jäst-arten Saccharomyces cerevisiae, den en millimeter långa masken Caenorhabditis elegans och husmusen.
Tillämpningar
[redigera | redigera wikitext]Det finns många tillämpningar av genteknik, bland annat genetisk släktforskning, genterapi, genmodifikation och kloning.
Historia
[redigera | redigera wikitext]Människor har i alla tider undrat över hur exempelvis människobarn kommer att likna sina föräldrar, utan att vara exakta kopior av dem.
Redan i förhistorisk tid började människan använda genetisk kunskap för domesticering och förädling av växter och djur. Friedrich Miescher (1844-1895) beskrev första gången 1869 en substans som han kallade ”nuklein”, som han funnit i cellkärnor. Något senare lyckades han framställa detta ämne i ren form genom att utgå från laxsperma, och 1889 fick ämnet namnet ”nukleinsyra” av Mieschers elev Richard Altman. Man fann att ämnet endast existerade i kromosomerna.
Varken nukleinsyrans eller cellkärnas funktion var dock klarlagd vid denna tid. När Gregor Mendel upptäckte ärftlighetsprinciperna på 1860-talet, och när Mendels resultat återupptäcktes i början av 1900-talet, var det oklart var i cellerna arvsanlagen fanns, och vilka molekyler som var bärare av dem. Inte förrän 1865 publicerade Gregor Mendel sina rön om slumpmässig kombination av ärftlighetsanlag i avkomma, och de uppmärksammades inte förrän i början av 1900-talet.
När Jean-Baptiste de Lamarck och Charles Darwin lade fram sina teorier om evolution visste egentligen ingen av dem hur egenskaper nedärvs. Båda trodde till exempel att förvärvade egenskaper, som exempelvis muskelstyrka, ärvdes, vilket orättvist kommit att förknippas enbart med lamarckism.
På 1930-talet genomförde Max Delbrück med flera experiment som visade att man genom att utsätta celler för röntgenstrålar kunde förändra de ärftliga egenskaperna hos cellerna. Det föreslogs att kromosomernas kemiska struktur på något sätt bestämde dessa ärftliga egenskaper. Precis hur denna kemiska struktur kunde påverka en organisms egenskaper och beteende föreföll oförklarligt vid denna tidpunkt. De kemiska undersökningarna av olika nukleinsyrepreparat gav alltid samma resultat i form av de fyra typerna av nukleotider i ungefär samma proportioner. Kromosomernas kemiska uppbyggnad föreföll alltså enkel och likformig vilket stod i stark kontrast till de levande organismernas komplexitet, mångfald och variation.
På 1950-talet pågick forskning om DNA-molekylens struktur endast på några få ställen. En grupp forskare i USA leddes av Linus Pauling. I England intresserade sig två grupper för problemet. Vid University of Cambridge fanns bland andra Francis Crick och James Watson och vid King's College i London arbetade Maurice Wilkins och Rosalind Franklin med att med hjälp av röntgendiffraktion fastställa DNA-molekylens struktur. 1948 hade Pauling upptäckt att många proteiner hade en helixstruktur, och de första undersökningarna med röntgendiffraktion antydde att även DNA hade en sådan struktur, men någon detaljerad förståelse av molekylens uppbyggnad hade man ännu inte.

Crick och Watson försökte konstruera rimliga modeller utgående från kända fakta, men antalet möjligheter var fortfarande många. Ett genombrott skedde när den österrikiske kemisten Erwin Chargaff besökte Cambridge och beskrev ett av sina experiment. Han hade fastställt att prover av DNA inte alltid hade samma proportioner av de olika nukleotiderna, men att de alltid hade lika koncentration av adenin som av tymin och lika koncentration av guanin som av cytosin. Crick och Watson började fundera på strukturer som innefattade två trådar med kompletterande nukeotidbaser bundna till varandra. Med hjälp av information från Rosalind Franklins röntgendiffraktionsbilder lyckades de finna en modell som stämde med all kända fakta. Den hade en helixstruktur med två nanometers tjocklek och en höjd av cirka 3,4 nanometer per varv omfattande cirka 10 baspar. De skyndade sig att publicera sina idéer innan Franklin själv hade offentliggjort några av sina resultat.
Det har efteråt blivit en kontroversiell fråga hur mycket Watson och Crick varit beroende av Franklins data för att komma fram till sin modell, och många har anklagat dem för att inte ge henne tillräckligt erkännande av hennes betydelse i upptäckten av DNA-molekylens struktur. Mest omdebatterat är det faktum att Wilkins tydligen visat Franklins bilder för Watson och Crick när Franklin inte själv var närvarande. Wilkins, Watson och Crick fick nobelpriset i medicin 1962 för sina upptäckter. Vid denna tidpunkt hade Franklin avlidit.
Watsons och Cricks modell väckte stor uppmärksamhet när den publicerats. Efter att ha kommit fram till sin modell 21 februari 1953, gjorde de sina första uttalanden den 28 februari. Den 25 april publicerades deras artikel ”A structure for Deoxiribose Nucleic Acid”. Forskningen om genetikens och molekylärbiologins grundvalar tog sedan fart. I en föreläsning 1957 redogjorde Crick för sina idéer om kopplingen mellan DNA, RNA och proteiner: ”DNA ger RNA ger protein”, något som har kommit att kallas molekylärbiologins ”centrala dogm”. Crick och hans medarbetare fortsatte sedan under slutet av 1950-talet med arbetet med att knäcka den genetiska koden.
Trots att DNA:s struktur gav en tydlig fingervisning om hur själva nedärvningen går till visste man ändå inte hur DNA påverkar cellernas funktion. Under de följande åren kartlade vetenskapsmännen hur DNA styr de processer som producerar proteiner. Man upptäckte att DNA används som en mall för att skapa en motsvarande RNA-sträng, budbärar-RNA. Nukleotidsekvensen i budbärar-RNA används som mall vid skapandet av en sekvens av aminosyror i ett protein. Denna översättning mellan nukleotider och aminosyror kallas den genetiska koden.
Dessa insikter i den molekylära grunden för nedärvning blev grogrunden för en närmast explosiv utveckling av forskningen. En viktig utveckling var Frederick Sangers metod för DNA-sekvensering, som kom 1977. Med denna metod kan man avläsa nukleotidsekvensen i DNA-molekylen.[44] År 1983 utvecklade Kary Banks Mullis polymeraskedjereaktionen, som snabbt kan isolera och mångfaldiga ett specifikt önskat avsnitt utifrån enblandning av DNA.[45] Genom de samordnade insatserna i Human Genome Project och det parallella arbetet i företaget Celera Genomics kunde man med hjälp av dessa och andra tekniker kartlägga i stort sett hela människans genom.[46]
Se även
[redigera | redigera wikitext]- Evolution
- Gen
- Allel
- Genteknik
- Genterapi
- GMO
- Monohybrid klyvning
- Dihybrid klyvning
- Intermediär klyvning
- Polyhybrid klyvning
- Återkorsning
- Överkorsning
- Mem
- Translokation
- Epigenetik
- Personlighetsgenetik
- Humangenetik
- Arv (genetik)
- Hereditet
- Heritabilitet
Källor
[redigera | redigera wikitext]- Den här artikeln är helt eller delvis baserad på material från engelskspråkiga Wikipedia, Genetics, tidigare version.
- ^ Pearson, H (12 november 2006). ”Genetics: what is a gene?”. Nature "441" (7092): ss. 398–401. doi:. ISSN 0028-0836. PMID 16724031.
- ^ Prescott, L (1993). Microbiology. Wm. C. Brown Publishers. ISBN 0697013723
- ^ Griffiths, William M.; Miller, Jeffrey H.; Suzuki, David T. et al., reds (2000). ”Mechanism of DNA Replication”. An Introduction to Genetic Analysis (7th). New York: W. H. Freeman. ISBN 0-7167-3520-2
- ^ Gregory, SG (12 november 2006). ”The DNA sequence and biological annotation of human chromosome 1.”. Nature "441" (7091): ss. 315–21. doi:. PMID 16710414.
- ^ Alberts et al. (2002), II.4. DNA and chromosomes: Chromosomal DNA and Its Packaging in the Chromatin Fiber
- ^ [a b c] Griffiths, William M.; Miller, Jeffrey H.; Suzuki, David T. et al., reds (2000). ”Mendelian genetics in eukaryotic life cycles”. An Introduction to Genetic Analysis (7th). New York: W. H. Freeman. ISBN 0-7167-3520-2
- ^ Griffiths, William M.; Miller, Jeffrey H.; Suzuki, David T. et al., reds (2000). ”Sex chromosomes and sex-linked inheritance”. An Introduction to Genetic Analysis (7th). New York: W. H. Freeman. ISBN 0-7167-3520-2
- ^ Griffiths, William M.; Miller, Jeffrey H.; Suzuki, David T. et al., reds (2000). ”Bacterial conjugation”. An Introduction to Genetic Analysis (7th). New York: W. H. Freeman. ISBN 0-7167-3520-2
- ^ Griffiths, William M.; Miller, Jeffrey H.; Suzuki, David T. et al., reds (2000). ”Nature of crossing-over”. An Introduction to Genetic Analysis (7th). New York: W. H. Freeman. ISBN 0-7167-3520-2
- ^ Griffiths, William M.; Miller, Jeffrey H.; Suzuki, David T. et al., reds (2000). ”Linkage maps”. An Introduction to Genetic Analysis (7th). New York: W. H. Freeman. ISBN 0-7167-3520-2
- ^ Berg JM, Tymoczko JL, Stryer L, Clarke ND (2002). ”I. 5. DNA, RNA, and the Flow of Genetic Information: Amino Acids Are Encoded by Groups of Three Bases Starting from a Fixed Point”. Biochemistry (5th). New York: W. H. Freeman and Company
- ^ Crick, F (12 november 1970). ”Central dogma of molecular biology.”. Nature "227" (5258): ss. 561–3. doi:. PMID 4913914. http://www.nature.com/nature/focus/crick/pdf/crick227.pdf.
- ^ Alberts et al. (2002), I.3. Proteins: The Shape and Structure of Proteins
- ^ Alberts et al. (2002), I.3. Proteins: Protein Function
- ^ ”How Does Sickle Cell Cause Disease?”. Brigham and Women's Hospital: Information Center for Sickle Cell and Thalassemic Disorders. 11 april 2002. http://sickle.bwh.harvard.edu/scd_background.html. Läst 23 juli 2007.
- ^ Brivanlou, AH (12 november 2002). ”Signal transduction and the control of gene expression.”. Science "295" (5556): ss. 813–8. doi:. PMID 11823631.
- ^ Alberts et al. (2002), II.3. Control of Gene Expression – The Tryptophan Repressor Is a Simple Switch That Turns Genes On and Off in Bacteria
- ^ Jaenisch, R (12 november 2003). ”Epigenetic regulation of gene expression: how the genome integrates intrinsic and environmental signals.”. Nature genetics "33 Suppl": ss. 245–54. doi:. PMID 12610534.
- ^ Chandler, VL (12 november 2007). ”Paramutation: from maize to mice.”. Cell "128" (4): ss. 641–5. doi:. PMID 17320501.
- ^ Imes, DL (12 november 2006). ”Albinism in the domestic cat (Felis catus) is associated with a tyrosinase (TYR) mutation.”. Animal genetics "37" (2): ss. 175–8. doi:. PMID 16573534.
- ^ ”MedlinePlus: Phenylketonuria”. NIH: National Library of Medicine. http://www.nlm.nih.gov/medlineplus/phenylketonuria.html. Läst 15 mars 2008.
- ^ Griffiths, William M.; Miller, Jeffrey H.; Suzuki, David T. et al., reds (2000). ”Patterns of Inheritance: Introduction”. An Introduction to Genetic Analysis (7th). New York: W. H. Freeman. ISBN 0-7167-3520-2
- ^ Blumberg, Roger B.. ”Mendel's Paper in English”. http://www.mendelweb.org/Mendel.html.
- ^ Griffiths, William M.; Miller, Jeffrey H.; Suzuki, David T. et al., reds (2000). ”Mendel's experiments”. An Introduction to Genetic Analysis (7th). New York: W. H. Freeman. ISBN 0-7167-3520-2
- ^ Griffiths, William M.; Miller, Jeffrey H.; Suzuki, David T. et al., reds (2000). ”Interactions between the alleles of one gene”. An Introduction to Genetic Analysis (7th). New York: W. H. Freeman. ISBN 0-7167-3520-2
- ^ Merriam, J. (2001-01-01). Sydney Brenner, Jefferey H. Miller. red. Encyclopedia of Genetics. Academic Press. sid. 1180. doi:. ISBN 9780122270802. http://www.sciencedirect.com/science/article/pii/B0122270800008156. Läst 4 september 2019
- ^ Cheney, Richard W.. ”Genetic Notation”. Arkiverad från originalet den 3 januari 2008. https://web.archive.org/web/20080103021518/http://faculty.users.cnu.edu/rcheney/Genetic%20Notation.htm. Läst 18 mars 2008.
- ^ Griffiths, William M.; Miller, Jeffrey H.; Suzuki, David T. et al., reds (2000). ”Human Genetics”. An Introduction to Genetic Analysis (7th). New York: W. H. Freeman. ISBN 0-7167-3520-2
- ^ Griffiths, William M.; Miller, Jeffrey H.; Suzuki, David T. et al., reds (2000). ”Gene interaction and modified dihybrid ratios”. An Introduction to Genetic Analysis (7th). New York: W. H. Freeman. ISBN 0-7167-3520-2
- ^ Mayeux, R (12 november 2005). ”Mapping the new frontier: complex genetic disorders.”. The Journal of clinical investigation "115" (6): ss. 1404–7. doi:. PMID 15931374.
- ^ Griffiths, William M.; Miller, Jeffrey H.; Suzuki, David T. et al., reds (2000). ”Quantifying heritability”. An Introduction to Genetic Analysis (7th). New York: W. H. Freeman. ISBN 0-7167-3520-2
- ^ Luke, A (12 november 2001). ”Heritability of obesity-related traits among Nigerians, Jamaicans and US black people.”. International journal of obesity and related metabolic disorders "25" (7): ss. 1034–41. doi:. PMID 11443503.
- ^ Griffiths, William M.; Miller, Jeffrey H.; Suzuki, David T. et al., reds (2000). ”Spontaneous mutations”. An Introduction to Genetic Analysis (7th). New York: W. H. Freeman. ISBN 0-7167-3520-2
- ^ Freisinger, E (12 november 2004). ”Lesion (in)tolerance reveals insights into DNA replication fidelity.”. The EMBO journal "23" (7): ss. 1494–505. doi:. PMID 15057282.
- ^ Griffiths, William M.; Miller, Jeffrey H.; Suzuki, David T. et al., reds (2000). ”Induced mutations”. An Introduction to Genetic Analysis (7th). New York: W. H. Freeman. ISBN 0-7167-3520-2
- ^ Sawyer, SA (12 november 2007). ”Prevalence of positive selection among nearly neutral amino acid replacements in Drosophila.”. Proceedings of the National Academy of Sciences of the United States of America "104" (16): ss. 6504–10. doi:. PMID 17409186.
- ^ Griffiths, William M.; Miller, Jeffrey H.; Suzuki, David T. et al., reds (2000). ”Variation and its modulation”. An Introduction to Genetic Analysis (7th). New York: W. H. Freeman. ISBN 0-7167-3520-2
- ^ Griffiths, William M.; Miller, Jeffrey H.; Suzuki, David T. et al., reds (2000). ”Selection”. An Introduction to Genetic Analysis (7th). New York: W. H. Freeman. ISBN 0-7167-3520-2
- ^ Griffiths, William M.; Miller, Jeffrey H.; Suzuki, David T. et al., reds (2000). ”Random events”. An Introduction to Genetic Analysis (7th). New York: W. H. Freeman. ISBN 0-7167-3520-2
- ^ Darwin, Charles (1859). On the Origin of Species (1st). London: John Murray. sid. 1. ISBN 0801413192. http://darwin-online.org.uk/content/frameset?itemID=F373&viewtype=text&pageseq=16. Related earlier ideas were acknowledged in Darwin, Charles (1861). On the Origin of Species (3rd). London: John Murray. sid. xiii. ISBN 0801413192. http://darwin-online.org.uk/content/frameset?itemID=F381&viewtype=text&pageseq=20
- ^ Gavrilets, S (12 november 2003). ”Perspective: models of speciation: what have we learned in 40 years?”. Evolution; international journal of organic evolution "57" (10): ss. 2197–215. doi:. PMID 14628909.
- ^ Wolf, YI (12 november 2002). ”Genome trees and the tree of life.”. Trends in genetics "18" (9): ss. 472–9. doi:. PMID 12175808.
- ^ ”The Use of Model Organisms in Instruction”. University of Wisconsin: Wisconsin Outreach Research Modules. Arkiverad från originalet den 13 mars 2008. https://web.archive.org/web/20080313023531/http://www.loci.wisc.edu/outreach/text/model.html. Läst 15 mars 2008.
- ^ Sanger, F (12 november 1977). ”DNA sequencing with chain-terminating inhibitors”. Proceedings of the National Academy of Sciences of the United States of America "74" (12): ss. 5463–7. doi:. PMID 271968.
- ^ Saiki, RK (12 november 1985). ”Enzymatic amplification of beta-globin genomic sequences and restriction site analysis for diagnosis of sickle cell anemia.”. Science "230" (4732): ss. 1350–4. doi:. PMID 2999980.
- ^ ”Human Genome Project Information”. Human Genome Project. Arkiverad från originalet den 15 mars 2008. https://web.archive.org/web/20080315062131/http://www.ornl.gov/sci/techresources/Human_Genome/home.shtml. Läst 15 mars 2008.
Vidare läsning
[redigera | redigera wikitext]- Bengtsson, Bengt Olle (1999) Genetik och politik. Berättelser om en vetenskap mitt i samhället. Lund: Arkiv förlag
| ||||||||||||||
| |||||