Hemoglobină
Hemoglobina este pigmentul respirator (pigmentul roșu), a tuturor vertebratelor, fiind prezentă în eritrocite, unde are rol în transportul oxigenului[A] (oxihemoglobină), și a CO2 (carbaminohemoglobină). Constituită dintr-o componentă proteică numită globină[B] și o componentă prostetică, colorată, hemul, un nucleu porfirinic cu 11 duble legături conjugate. Molecula hemoglobinei este un ansamblu alcătuit din 4 subunități proteice, formate dintr-un lanț proteic strâns asociat cu grupul hemic. Fiecare lanț proteic adoptă o conformație de alfa helix identică cu globina din alte proteine (mioglobina de exemplu).
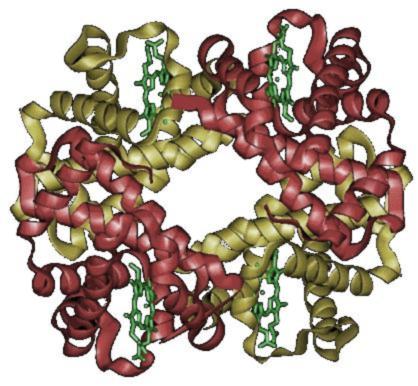

Structură
modificareGruparea hemică are un atom de fier cuplat cu inelul porfirinic. Atomul de fier realizează o legătură puternică cu proteina globulară prin intermediul nucleului imidazolic al unei molecule de histidină[C], proteină situată de obicei în planul inferior comparativ cu planul hem-ului.
Planul superior este ocupat de o legătură reversibilă cu oxigenul din heterociclu, configurația compusului format fiind cea de tip octaedrică. Atunci când oxigenul nu este legat, locul său este luat de o moleculă de apă, conformația fiind a unui octaedru neregulat. Fierul are 2 ioni : Fe2+ și Fe3+, însă Fe3+ nu poate lega O; legarea O se face și cu oxidarea de către oxigen a Fe2+ la Fe3+, deci fierul trebuie să existe în forma de oxidare +2 pentru a lega oxigenul. La om hemoglobina are o conformație de tetramer, numită hemoglobina A, formată din 2 lanțuri α și 2 lanțuri β, formate fiecare din 141, respectiv 146 aminoacizi. Hemoglobina A se numerotează α1β2, subunitățile fiind legate prin punți de sare, legături de hidrogen și interacții hidrofobe astfel: α1β1 α1β2. Ionii de Fe2+ și Fe3+ conțin respectiv 24 25 e- (configurația electronică este [Ar]3d6 4s2, [Ar]3d5 4s2d).
Configurația ionului de Fe
modificareDin punct de vedere magnetic oxihemoglobina este diamagnetică,[D] dar configurația electronilor de joasă energie atât la nivelul oxigenului cât și a fierului denotă caracter paramagnetic. Oxigenul triplet cu cea mai joasă energie, are 2 electroni în orbitali moleculari de antilegătură. Ionul de Fe2+ are tendința de a adopta o configurație de spin înalt, electronii neparticipanți aflîndu-se de asemenea în orbitali de antilegătură. Fe3+ are de asemenea electroni neparticipanți. Toate aceste molecule au caracter paramagnetic și nu diamagnetic cum ar trebui în mod teoretic. Există două explicații pentru acest lucru:
- Fe2+ se leagă de oxigenul singlet. Amândouă speciile sunt diamagnetice.
- Spinul înalt al Fe3+ detremină legarea de O22- (anionul superoxid), manifestându-se antiferomagnetismul care aliniază electronii neparticipanți, determinând proprietăți diamagnetice.
- Fe2+ cu configurație de spin jos se leagă de O22-. Amândouă speciile sunt diamagnetice.
Spectroscopia electronica cu raze X sugerează că starea de oxidare a fierului este de aproximativ 3,2, spectrul în infraroșu a legăturii O-O sugerează un tip de legătură foarte asemănătoare cu configurația ionului superoxid. Starea de oxidare corectă este +3 pentru fier și –1 pentru oxigen. Diamagnetismul acestei configurații este dat de electronii ionului superoxid care au o aliniere antiferomagnetică, în sensul opus alinierii electronilor de Fe2+, opțiune nefiind deloc surprinzătoare deoarece singletul oxigenului precum și larga separație a încărcării sunt nefavorabile stării de energie înaltă. Schimbarea valenței de Fe2+ la Fe3+ determină scăderea mărimii atomului și permite acestuia să intre în același plan cu ciclurile pirolice, putând astfel coordina restul de histidină, inițiind schimbări alosterice îm lanțul globulinic. De fapt se pare că structura adoptată este una de echilibru între aceste 3 forme.
Complexarea liganzilor
modificareAtunci când oxigenul se leagă de ionul de Fe, are loc o contracție a atomului de Fe determinând mutarea în interioriul planului porfirinic, concomitent cu îndepărtarea de atomul de oxigen și apropierea de restul histidinic. La hemoglobina umană legarea oxigenului este de tip cooperativ. Hemoglobina are afinitate pentru oxigen, afinitate crescută de saturarea moleculei cu oxigen; primul atom de oxigen legat influențează forma site-ului de legare pentru următorul atom. Acest lucru este posibil, când datorită oxigenării hemoglobinei, subunitățile proteice ale acesteia suferă modificări, modificări ce se traduc prin schimbări în întregul complex hemoglobinic determinînd ca altor subunități să le crească afinitatea pentru oxigen.
Drept consecință, curba de legare a oxigenului de hemoglobină este de tip sigmoid sau S, opusă curbei de tip hiperbolă asociată cu legarea de tip noncooperativ. Legarea oxigenului de hemoglobină este scăzută în prezența monoxidului de carbon, deoarece acesta intră în competiție pentru site-urile de legare, monoxidul legându-se de preferință în aceleași locuri ca și oxigenul. Dioxidul de carbon are un alt situs de legare, neafectând formarea complexului HbO. Sub infleunța anhidrazei carbonice, dioxidul de carbon se scindează în acid carbonic instabil care se descompune imediat punând în libertate ionul carbonat HCO3- și un proton H+.
- CO2 + H2O → H2CO3 → HCO3- + H+
Sângele încărcat cu dioxid de carbon are un pH acid, și din acest motiv poate lega protonul și dioxidul de carbon determinând transformări conformaționale în partea proteică și la nivelul capacității de legare și de eliberare a oxigenului. Protonul poate fi legat în diferite locuri de-a lungul lanțului proteic, dar dioxidul de carbon se poate lega doar de gruparea amino din poziția alpha formând ionul carbamat. Atunci când nivelul de dioxid de carbon scade (de exemplu în capilarele pulmonare), CO2 și H sunt eliberați de la nivelul hemoglobinei, crescînd afinitatea proteinei pentru O2. Acest control al hemoglobinei pentru oxigen prin legarea și eliberarea CO2 și acidului poartă denumirea de efectul Bohr. Monoxidul de carbon afectează legarea oxigenului de hemoglobină, datorită afinității de circa 200 de ori mai mare față de oxigen. Acest lucru este tradus prin aceea că o cantitate extrem de mică de CO (provenit de exemplu din fumul de țigară, eșapamentul automobilelor), reduce foarte mult capacitatea de legare a O2 de către hemoglobină. Monoxidul formează carboxihemoglobina, un compus extrem de stabil, de culoare roșu strălucitor. Asemănător cu monoxidul de carbon se comportă și ionii CN- (cian), SO (monoxidul de sulf), NO2 (dioxid de azot), S2- (sulfit). Toți acești compuși manifestă afinitate crescută pentru ionul de Fe de care se leagă fără a-i schimba starea de oxidare, și mai grav neinhibând legarea oxigenului, determinând intoxicații grave. Oxidarea Fe2+ la Fe3+ determină conversia hemoglobinei la hemiglobină sau methemoglobină, compus care nu poate lega oxigenul. În sângele normal hemoglobina este protejată printr-un sistem reducător de această transformare. Însă anumiți anioni de tipul oxidului nitros (N2O), sau dioxidului de azot (NO2), sunt capabili să transforme o fracție foarte mică de hemoglobină în methemoglobină, acest lucru neavând importanță medicală (NO2 este nociv prin alt mecanism), în timp ce oxidul nitros (N2O) este folosit în anestezii de scurtă durată, ceea ce nu permite afectarea hemoglobinei. La populațiile aclimatizate la înălțimi mari concentrația esterului glicerin-2-3-difosfat sanguin este mult crescută, ceea ce permite transportul unor cantități mai mari de oxigen la nivelul țesuturilor în condițiile unei presiuni a oxigenului scăzută. Acest fenomen în care o moleculă Y afectează legarea moleculei X pentru transportul moleculei Z.
Degradarea hemoglobinei
modificareAtunci când are loc moartea hematiilor, membrana lor se rupe, hemoglobina este hidrolizată iar Fe recuperat. Ciclul porfirinic se desface iar fragmentele sunt metabolizate de ficat. În urma procesului rezultă o moleculă de CO pentru fiecare moleculă de hemoglobină metabolizată. Procesul este unul natural și reprezintă o sursă de CO pentru corpul uman, monoxid care se elimină prin aerul expirat, sub forma de dioxid de carbon (CO2). Produsul de metabolizare final al globinei este bilirubina, un pigment de culoare galbenă a cărui nivel seric crește odată cu distrugerea excesivă a hematiilor. În hemoliză (distrugere a hematiilor mai rapidă decât sinteza), icterul asociat este cauzat de metabolitul bilirubină și de hemoglobina circulantă care poate determina blocaj renal. Dacă ritmul de degradare a hemoglobinei este prea rapid, compusii ei de degradare pot bloca anumite capilare mai ales capilarele din rinichi determinând apariția unor boli. Scăderea hemoglobinei neasociată cu scăderea numărului de hematii poartă denumirea de anemie; anemia are drept principală cauză deficiența de Fe. Ca urmare a acestei deficiențe, scade sinteza de hemoglobină, hematiile vor fi de tip hipocrom (sărace în pigment) și microcitic (mai mici decât în mod normal).
Hemoglobinopatiile sunt mutații la nivelul lanțului globinei, cele mai des întâlnite fiind anemia falciformă (siclemie) și talasemie; tot în cadrul globinopatiilor sunt și porfiriile, caracterizate de erori în sinteza hemului.
La nivel mai mic se pare că hemoglobina A se combină cu glucoza, pe care o leagă într-un anumit situs, formînd compusul HbA1c (hemoglobina glucosilată, sau glicată). Pe măsură ce concentrația glucozei crește , procentul de HbA trasnformată în HbA1c crește și el. La diabetici, unde glucoza are valori foarte mari, și compusul tinde către valori mari, procentul său fiind reprezentativ pentru nivelul glucozei sanguine datorită timpului de viață a hematiilor 120 zile.
Măsurare (teste de laborator)
modificareHemoglobina este urmărită în cadrul testelor sanguine (odată cu numărarea hematiilor și celorlalte componente sanguine celulare), rezultatele fiind exprimate în unități de concentrație masică: g/L, g/dL, mol/L (1g/dL = 0,621 mmol/L). Dacă concentrația Hb totale scade sub acest punct, este vorba de o anemie. Determinarea Hb se face prin analiză sanguină și urmărirea valorii hematocritului care reprezintă volumul ocupat de hematii în cadrul volumului sanguin. Datorită legării glucozei la Hb cu formarea complexului HbA1C (hemoglobina glicozilată), a cărui nivel este direct proporțional cu cantitatea glucozei sanguine, se poate monitoriza mai precis cantitatea glucozei sanguine care ar fi relativ greu de efectuat din cauza variațiilor orare ce ar da rezultate ce nu pot fi semnificative și pot da erori la interpretare. Pentru acest motiv se urmărește complexul HbA1C. De obicei o valoare mai mică sau egală cu 6% reprezintă un nivel normal al glucozei sanguine, în timp ce valori de peste 7% indică un nivel ridicat al glucozei (diabet). Urmărirea valorii complexului se poate face doar la subiecții sănătoși care au un ciclu al hematiilor normal. La indivizii cu dereglări ale acestui ciclu cum ar fi bolnavii cu anemie falciformă, unde se pot întâlni alte tipuri de hemoglobina (Hb S), ciclul de viață este mult mai scurt. Din acest motiv se folosește testul nivelului de fructozamină, test care are la bază măsurarea gradului de legare a glucozei de albumina serică, detereminîndu-se astfel nivelul sanguin al glucozei. Concentrația sanguină a hemoglobinei poate fi măsurată cu hemoglobinometru electronic.[E]
Tipuri de hemoglobină
modificareHemoglobină umană
modificare1.La embrion:
- Gower 1 (ξ2ε2)
- Gower 2 (α2ε2) [F]
- Hemoglobina Portland (ξ2γ2)
2.La făt:
- Hemoglobina F (α2γ2) [G]
3.La adult:
- Hemoglobina A (α2β2) [H] - Tipul cel mai des întînit.
- Hemglobina A2 (α2δ2) - Lanțurile δ se sintetizează cam din al III-lea semestru de sarcină și al adulți reprezintă cam 2,5%.
- Hemoglobina F (α2γ2) - La adulți hemoglobina F este restrînsă la o populație limitată de hematii numite celule F.
- Hemoglobina S - Tipul de hemoglobină întîlnită la hematiile în formă de seceră (anemia falciformă).
Alte tipuri de (hemo)globină
modificareMioglobina: Este identificată în mușchi, întîlnindu-se la toate vertebratele, unde colorează mușchii în roșu sau cenușiu închis. Este foarte asemănătoare cu hemoglobina, însă diferă de aceasta prin faptul ca nu prezintă unități tetramerice. De regulă stochează oxigenul pentru a fi transportat mai departe.
Hemocinina: Este al 2-lea transportor de oxigen din mediul animal, identificat la artropode și moluște, având heteroatom ionul de Cu, iar culoarea sa datorită acestui fapt este albastră atunci când leagă oxigenul.
Hemeritrina: Întâlnită la unele nevertebrate marine și anumite specii de anelide, având heteroatom ionul de Fe, culoarea sa este roz/violet când este oxigenată, incoloră când oxigenul nu este legat.
Clorocruorina: Identificată la multe anelide, asemănătoare cu eritrocruorina, însă grupare hemică este diferită. Când nu leagă oxigenul are culoarea verde, în timp ce oxigenată are culoare roșie.
Vanabina: Întâlnită la urocordate, se pare că heteroatomul este ionul de vanadiu (ipoteză neconfirmată).
Eritrocruorina: Identificată la multe anelide, este un complex proteic cu masa de peste 3,5 milioane daltoni.
Pinaglobina: Identificată doar la molusca Pinna squamosa, heteroatomul fiind ionul de Mn.
Leghemoglobina: Identificată la specii de leguminoase (lucernă, soia), bacteriile fixatoare de azot sunt protejate de această proteină care are drept heteroatom ionul de Fe.
Note
modificare- ADiagrama schimburilor gazoase în alveole și transportul oxigenului de către hemoglobină Arhivat în , la Wayback Machine. pe site-ul ZooBotanica
- BAN INTRODUCTION TO COMPLEX METAL IONS pe site-ul Chemguide
- CDiagrama legăturilor dintre atomul de Fe și hem
- DMagnetic properties of hemoglobin
- EPolicec 1988, p. 150-152
- FProtein Data Base, hemoglobina Glover2 Arhivat în , la Wayback Machine.
- GProtein Data Base, hemoglobina F Arhivat în , la Wayback Machine.
- HProtein Data Base, hemoglobina A Arhivat în , la Wayback Machine.
Bibliografie
modificare- Ion R. Baciu Homeostazia oxigenului Editura Dacia 1980
- V. Ranga, I. Teodorescu Exarcu, Anatomia și fiziologia omului, Editura Medicală, București, 1970
- A. Policec, T.D. Gligor, Gh. Ciocloda, Electronica medicală Editura Dacia, Cluj, 1983
Legături externe
modificare- http://www.ebi.ac.uk/interpro/potm/2005_10/Page1.htm
- http://www.tiscali.co.uk/reference/encyclopaedia/hutchinson/m0006815.html Arhivat în , la Wayback Machine.
- http://www.uni-bayreuth.de/departments/ddchemie/umat/eisenkomplex/eisenkomplex.htm
- http://www.fmrib.ox.ac.uk/~peterj/lectures/hbm_2/sld015.htm
- http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=1076743 Arhivat în , la Archive.is