Neoplasm
Un neoplasm este un tip de creștere anormală și excesivă, numită neoplazie, a unui țesut. Creșterea unui neoplasm este necoordonată cu cea a țesutului normal înconjurător și persistă să crească anormal, chiar dacă declanșatorul inițial este îndepărtat.[1][2][3] De obicei, această creștere anormală (dar nu întotdeauna) formează o masă. [4] Când formează o masă, poate fi numită tumoră.

ICD-10 (Clasificarea statistică internațională a bolilor) clasifică neoplasmele în patru grupe principale: neoplasme benigne, neoplasme in situ, neoplasme maligne și neoplasme de comportament incert sau necunoscut.[5] Neoplasmele maligne sunt, de asemenea, cunoscute pur și simplu sub numele de cancer și sunt în centrul oncologiei.
Înainte de creșterea anormală a țesutului, ca neoplazie, celulele suferă adesea un model anormal de creștere, cum ar fi metaplazia sau displazia.[6] Cu toate acestea, metaplazia sau displazia nu progresează întotdeauna spre neoplazie.[1] Cuvântul provine din greaca antică, din νέος- neo („nouă”) și πλάσμα plasmă („formare”, „creație”).
Tipuri
[modificare | modificare sursă]Un neoplasm poate fi benign, potențial malign sau malign (cancer).[7]
- Tumorile benigne includ: - fibroamele uterine, - osteofitele și - nevi melanocitici (alunițele pielii). Sunt circumscrise și localizate, și nu se transformă în cancer. [6]
- Neoplasmele potențial maligne includ carcinomul in situ. Sunt localizate, nu invadează și nu distrug, dar în timp, se pot transforma într-un cancer.
- Neoplasmele maligne sunt denumite frecvent cancer. Ele invadează și distrug țesutul din jur, pot forma metastaze și, dacă nu sunt tratate sau nu răspund la tratament, se dovedesc fatale.
- Neoplasmul secundar se referă la oricare dintre categoriile de tumori canceroase care sunt fie o afecțiune metastatică a unei tumori primare, fie o tumoră aparent fără legătură care crește în frecvență în urma anumitor tratamente pentru cancer, cum ar fi chimioterapia sau radioterapia.
- Rareori poate exista o neoplasmă metastatică fără un sit cunoscut al cancerului primar și aceasta este clasificată sub denumirea cancer de origine primară necunoscută.
Clonalitatea
[modificare | modificare sursă]Tumorile neoplazice sunt adesea eterogene și conțin mai multe tipuri de celule, dar inițierea și creșterea continuă depind de obicei de o singură populație de celule neoplastice. Se presupune că aceste celule sunt clonale - adică provin din aceeași celulă[8] și toate poartă aceeași anomalie genetică sau epigenetică - evidentă - a clonalității. Pentru neoplasmele limfoide, de exemplu limfomul și leucemia, clonalitatea este dovedită prin amplificarea unei singure rearanjări a genei imunoglobulinei lor (pentru leziunile celulelor B) sau a genei receptorului celulelor T (pentru leziunile celulelor T). Demonstrarea clonalității este considerată a fi necesară pentru a identifica o proliferare a celulelor limfoide ca neoplazică.[9]
Este tentant să definim neoplasmele ca proliferări celulare clonale, dar demonstrarea clonalității nu este întotdeauna posibilă. Prin urmare, nu se poate include clonalitatea în definirea neoplaziei.
Neoplazie vs. tumoră
[modificare | modificare sursă]Tumora sau tumoarea (în vorbirea curentă), însemnând în latină umflătură, unul dintre semnele cardinale ale inflamației, a însemnat inițial orice formă de umflătură (tumefacție), neoplazică sau nu. Cu toate acestea, terminologia actuală, atât medicală cât și non-medicală, folosește trmenul de tumoră ca sinonim pentru un neoplasm (o leziune chistică solidă sau plină de fluide, care poate fi sau nu formată printr-o creștere anormală a celulelor neoplazice), care apare cu o anumită mărime.
Unele neoplasme nu formează o tumoare, de exemplu leucemia și cele mai multe forme de carcinom in situ. Tumora nu este, de asemenea, sinonimă cu cancerul . În timp ce cancerul este, prin definiție, malign, o tumoră poate fi benignă, precanceroasă sau malignă.
Termenii de masă și nodul sunt adesea folosiți în mod sinonim cu tumora. În general, însă, termenul de tumoră este utilizat în mod generic, fără referire la dimensiunea fizică a leziunii.[1] Mai exact, termenul de masă este adesea utilizat atunci când leziunea are un diametru maxim de cel puțin 20 de milimetri (mm) în cea mai mare direcție, în timp ce termenul nodul este de obicei utilizat atunci când dimensiunea leziunii este mai mică de 20 de mm, în cea mai mare dimensiune.
Cauze
[modificare | modificare sursă]

Un neoplasm poate fi cauzat de o proliferare anormală a țesuturilor, care poate fi cauzată de mutații genetice. Nu toate tipurile de neoplasme determină o supraaglomerare tumorală a țesutului (vezi leucemia sau carcinomul in situ) scoțându-se în evidență asemănări între creșterile neoplazice și procesele regenerative, de exemplu, dediferențierea și proliferarea rapidă a celulelor.[10]
Recent, creșterea tumorii a fost studiată folosind matematica și mecanica mediilor continue. Tumorile vasculare (formate din vase de sânge) sunt astfel privite ca fiind amalgame ale unui schelet solid format din celule lipicioase și un lichid organic care umple spațiile în care celulele pot crește.[11] În cadrul acestui tip de model, pot fi tratate eforturi mecanice și tulpini, iar influența lor asupra creșterii tumorii, a țesutului înconjurător și a vascularizării este elucidată. Descoperirile recente din experimentele care folosesc acest model arată că creșterea activă a tumorii este limitată la marginile exterioare ale tumorii și că rigidizarea țesutului normal de bază inhibă și creșterea tumorii.[12]
Condiții benigne care nu sunt asociate cu o proliferare anormală a țesutului (cum ar fi chisturile sebacee sau ateroamele) pot fi, de asemenea, prezente ca tumori, dar nu au potențial malign. Chisturile mamare (care apar frecvent în timpul sarcinii și în alte momente) sunt un alt exemplu, la fel ca și alte umflături glandulare încapsulate (tiroidă, glanda suprarenală, pancreas).
Hematoamele încapsulate, țesutul necrotic încapsulat (de la o mușcătură de insectă, de la un corp străin sau alt mecanism nociv), keloizii (supraagregări discrete ale țesutului cicatricial) și granuloamele pot, de asemenea, să apară sub forma unor tumori.
Mărirea localizată discretă a structurilor normale (uretere, vase de sânge, conducte biliare intrahepatice sau extrahepatice, incluziuni pulmonare sau duplicări gastrointestinale ) cauzată de obstrucțiile sau îngustările de ieșire sau de conexiuni anormale, poate, de asemenea, să apară sub forma unei tumori. Exemple sunt fistulele arteriovenoase sau anevrismele (cu sau fără tromboză), fistulele biliare sau anevrisme, colangită sclerotică, cisticercoză sau chisturi hidatice, duplicări intestinale și incluziuni pulmonare, așa cum se observă la fibroza chistică.
Biopsierea unui număr de tipuri de tumori poate fi periculoasă, întrucât scurgerea conținutului lor ar putea fi catastrofală.
Când se întâlnesc astfel de tipuri de tumori, modalitățile de diagnostic, cum ar fi ecografia, tomografiile, RMN-ul, angiogramele și scanările de medicină nucleară sunt folosite înainte (sau în timpul) biopsiei sau explorării/excizării chirurgicale, în încercarea de a evita astfel de complicații severe.
Natura unei tumori este determinată de imagistică, de explorare chirurgicală sau de un patolog după examinarea țesutului dintr-o biopsie sau un specimen chirurgical.
Neoplasme maligne
[modificare | modificare sursă]Deteriorări ale ADN-ului
[modificare | modificare sursă]
Deteriorarea ADN-ului este considerată principala cauză a apariției neoplasmelor maligne cunoscute sub numele de cancer.[13][14] Rolul său central în progresia spre cancer este ilustrat în figura din această secțiune, în caseta din apropierea vârfului. (Caracteristicile centrale ale afectării ADN-ului, modificările epigenetice și reparația deficitară a ADN-ului în progresia cancerului sunt arătate în roșu.)
Deteriorarea ADN-ului este foarte frecventă. Deteriorările sau erorile ADN-ului care apar în mod natural (în mare parte datorită metabolismului celular și proprietăților ADN-ului în apă la temperaturi ale corpului) apar la o rată de peste 60.000 de erori noi, în medie, pe celulă umană, pe zi [13] [a se vedea și articolul Deteriorarea ADN-ului (care apare în mod natural)].
Deteriorări ale ADN suplimentare pot apărea din cauza expunerii la agenți exogeni. Fumul de tutun provoacă o deteriorare exogenă a ADN-ului, iar aceste deteriorări ale ADN sunt cauza probabilă a cancerului pulmonar datorat fumatului. [15] Lumina UV provenită din radiațiile solare provoacă deteriorări ale ADN care sunt importante în melanom. [16] Infecția cu Helicobacter pylori produce niveluri ridicate de specii reactive de oxigen care afectează ADN-ul și contribuie la cancerul gastric.[17] Acizii biliari, la niveluri ridicate în colonul oamenilor care consumă o dietă bogată în grăsimi, provoacă, de asemenea, daune ADN-ului și contribuie la cancerul de colon. [18] Katsurano și colab. au indicat că macrofagele și neutrofilele dintr-un epiteliu colonic inflamat sunt sursa de specii reactive de oxigen care provoacă deteriorări ale ADN care, la rândul lor, inițiază tumorageneză colonică. [19] Unele surse de deteriorare a ADN-ului sunt indicate în casetele din partea de sus a figurii din această secțiune.
Persoanele cu o mutație a liniei germinale care provoacă o deficiență în oricare din cele 34 de gene de reparare a ADN-ului (a se vedea articolul tulburări de deficiență de reparare a ADN-ului ) prezintă un risc crescut de cancer. Unele mutații ale liniei germinale din genele de reparare a ADN-ului provoacă până la 100% șanse de cancer în orice moment al vieții (de exemplu, mutațiile p53).[20] Aceste mutații ale liniei germinale sunt indicate într-o casetă din stânga figurii cu o săgeată care indică contribuția lor la deficiența de reparare a ADN-ului.
Aproximativ 70% dintre neoplasmele maligne nu au nici o componentă ereditară și sunt numite cancere sporadice.[21] Doar o minoritate de tipuri de cancer sporadice prezintă o deficiență în repararea ADN-ului din cauza mutației într-o genă de reparare a ADN-ului.
Cu toate acestea, majoritatea cancerelor sporadice au deficiență în repararea ADN-ului din cauza modificărilor epigenetice care reduc sau reduc la tăcere expresiile genelor de reparare a ADN-ului. De exemplu, din 113 cancere colorectale secvențiale, doar patru au avut o missense mutation (eroare de "traducere") în gena de reparație MGMT a ADN-ului, în timp ce majoritatea au redus expresia genei MGMT datorită metilării regiunii promotoare a genei MGMT (o modificare epigenetică). [22] Cinci rapoarte prezintă dovezi că între 40% și 90% dintre cancerele colorectale au redus expresia MGMT grație metilării regiunii promotoare a genei MGMT.[23][24][25][26][27]
În mod similar, din cele 119 cazuri de cancere colorectale deficitare de reparație care nu aveau expresia genei de reparare PMS2 a ADN-ului, gena PMS2 era deficitară în 6 cazuri din cauza mutațiilor din gena PMS2, în timp ce în 103 cazuri expresia PMS2 era deficitară, deoarece partenera sa de împerechere MLH1 a fost reprimată ddatorită metilării promotorului (proteina PMS2 este instabilă în absența MLH1).[28] În celelalte 10 cazuri, pierderea expresiei PMS2 s-a datorat probabil supraexpresiei epigenetice a microRNA, miR-155, care reglementează în jos MLH1.[29]
În alte exemple, au fost găsite defecte epigenetice la frecvențe cuprinse între 13% -100% pentru genele de reparație BRCA1, WRN, FANCB, FANCF, MGMT, MLH1, MSH2, MSH4, ERCC1, XPF, NEIL1 și ATM ale ADN-ului. Aceste defecte epigenetice au apărut în diferite tipuri de cancer (de exemplu, sân, ovar, colorectal, cap și gât). Două sau trei deficiențe de exprimare a ERCC1, XPF sau PMS2 apar simultan în majoritatea celor 49 de cancere de colon evaluate de Facista și colab.[30] Modificările epigenetice care determină expresia redusă a genelor de reparație a ADN-ului sunt prezentate într-o casetă centrală la nivelul al treilea din partea superioară a figurii din această secțiune, iar deficiența consecutivă de reparare a ADN-ului este prezentată la nivelul patru.
Atunci când expresia genelor de reparare a ADN-ului este redusă, deteriorările ADN-ului se acumulează în celule la un nivel mai mare decât normal, iar aceste deteriorări în exces provoacă frecvențe crescute de mutație sau epimutație. Ratele de mutație cresc puternic în celulele defecte în reparația nepotrivită a ADN-ului[31][32] sau în reparația omologă recombinațională (HRR). [33]
În timpul reparației pauzelor cu ADN dublu, sau repararea altor deteriorări ale ADN-ului, locurile de reparație incomplete pot provoca o amortizare a genelor epigenetice.[34][35] Deficiențele de reparare a ADN-ului (nivelul 4 din figură) determină creșterea deteriorărilor ADN-ului (nivelul 5 din figură) ceea ce duce la creșterea mutațiilor somatice și modificări epigenetice (nivelul 6 din figură).
Defectele de câmp, țesutul normal care apare cu alterații multiple (și discutate în secțiunea de mai jos), sunt precursori obișnuiți ai dezvoltării unei clone de țesut dezordonate și care proliferează necorespunzător într-un neoplasm malign. Astfel de defecte de câmp (al doilea nivel din partea de jos a figurii) pot avea mutații multiple și modificări epigenetice.
Odată format un cancer, de obicei există o instabilitate a genomului. Această instabilitate este probabil cauzată de o reparare redusă a ADN-ului sau de o deteriorare excesivă a ADN-ului. Din cauza unei astfel de instabilități, cancerul continuă să evolueze și să producă sub-clone. De exemplu, un cancer renal, eșantionat în 9 zone, a avut 40 de mutații omniprezente, care demonstrează eterogenitatea tumorii (adică prezent în toate zonele cancerului), 59 de mutații împărtășite de unele (dar nu de toate) zone și doar 29 de mutații „private” prezente într-una din zonele cancerului. [36]
Defecte de câmp
[modificare | modificare sursă]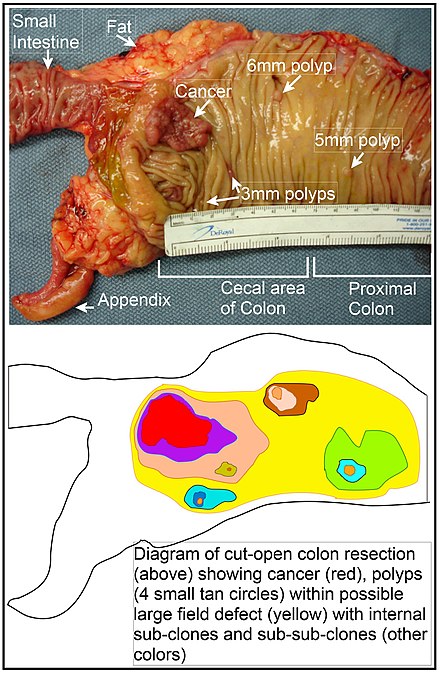
Pentru a descrie acest fenomen au fost folosiți mulți alți termeni, inclusiv „efectul de câmp”, „cancerizarea câmpului” sau „ carcinogeneza câmpului”. Termenul d „cancerizarea câmpului” a fost folosit pentru prima dată în 1953, pentru a descrie o zonă sau un „câmp” al epiteliului care a fost precondiționat prin (la acea vreme) procese în mare parte necunoscute, astfel încât să-l predispună spre dezvoltarea cancerului. [37] De atunci, termenii de „cancer de câmp” și „defect de câmp” au fost folosiți pentru a descrie țesutul pre-malign în care sunt posibile să apară noi cancere.
Defectele de câmp sunt importante în progresia cancerului.[38][39] Cu toate acestea, "majoritatea cercetărilor în domeniul cancerului, sublinia Rubin[40] au fost făcute pe tumori bine definite in vivo sau pe focare neoplazice discrete in vitro. Cu toate acestea, există dovezi că mai mult de 80% din mutațiile somatice găsite în fenotipul mutator al tumorilor colorectale umane apar înainte de debutul expansiunii clonale terminale.[41] În mod similar, Vogelstein și colab.[42] subliniază că mai mult de jumătate din mutațiile somatice identificate în tumori au avut loc într-o fază pre-neoplazică (într-un defect de câmp), în timpul creșterii celulelor aparent normale. De asemenea, modificări epigenetice prezente în tumori pot să fi apărut în defecte de câmp pre-neoplazice.
O viziune extinsă asupra efectului de câmp a fost denumită „efect etiologic de câmp”, care cuprinde nu numai modificări moleculare și patologice ale celulelor pre-neoplazice, dar și influențe ale factorilor de mediu exogeni și modificări moleculare în microambientul local asupra evoluției neoplazice de la inițierea tumorii până la moartea pacientului. [43]
În colon, un defect de câmp apare probabil prin selecția naturală a unei celule mutante sau modificate epigenetic printre celulele stem de la baza uneia dintre criptele intestinale de pe suprafața interioară a colonului. O celulă stem mutantă sau modificată epigenetic poate înlocui celelalte celule stem din apropiere prin selecție naturală. Astfel, poate apărea un petic de țesut anormal. Figura din această secțiune include o fotografie a unui segment proaspăt rezecat și deschis pe lungimea colonului care prezintă un cancer de colon și patru polipi. Sub fotografie există o diagramă schematică a modului în care s-ar fi putut forma un plasture mare de celule mutante sau alterate epigenetic, arătat de zona mare în galben din diagramă. În cadrul acestui prim plasture mare din diagrama (o clonă mare de celule), poate apărea o a doua astfel de mutație sau alterare epigenetică, astfel încât o anumită celulă stem dobândește un avantaj în comparație cu alte celule stem din interiorul patch-ului, iar această celulă stem modificată se poate extinde formând clonal un plasture secundar, sau o sub-clonă, în interiorul patch-ului original. Acest lucru este indicat în diagrama de patru patch-uri mai mici de culori diferite în zona originală mare galbenă. În cadrul acestor noi patch-uri (sub-clone), procesul poate fi repetat de mai multe ori, indicat de patchurile încă mai mici din cele patru patchuri secundare (cu culori încă diferite în diagramă) care se extind clonal, până când apar celule stem care generează fie mici polipi, fie un neoplasm malign (cancer).
În fotografie, un defect de câmp aparent în acest segment al unui colon a generat patru polipi (etichetați cu dimensiunea polipilor, 6mm, 5mm și doi de 3mm și un cancer de aproximativ 3 cm în cea mai lungă dimensiune). Aceste neoplasme sunt, de asemenea, indicate în diagrama de mai jos a fotografiei, de 4 cercuri mici arămii (polipi) și o zonă roșie mai mare (cancer). Cancerul din fotografie s-a produs în zona cecală a colonului, unde colonul se alătură intestinului subțire (etichetat) și unde apare apendicele (etichetat). Grăsimea din fotografie este exterioară peretelui exterior al colonului. În segmentul de colon prezentat aici, colonul a fost tăiat în lungime pentru a-i expune suprafața interioară și pentru a afișa cancerul și polipii care apar în mucoasa epitelială interioară a colonului.
Dacă procesul general prin care apar cancerele de colon sporadice este formarea unei clone pre-neoplazice care se răspândește prin selecție naturală, urmată de formarea subclonelor interne în cadrul clonei inițiale și sub-sub-clone în interiorul acestora, atunci cancerele de colon în general, ar trebui să fie asociate și să fie precedate de câmpurile de anomalii în creștere care reflectă succesiunea evenimentelor premaligne. Cea mai extinsă regiune de anormalitate (zona neregulată galbenă cea mai exterioară din diagramă) ar reflecta cel mai devreme eveniment în formarea unui neoplasm malign.
În cazul unor cancere, în evaluarea experimentală a deficiențelor specifice în repararea ADN-ului, multe deficiențe specifice de reparare a ADN-ului au fost, de asemenea, arătate ca aparând în defectele de câmp din jurul acestor tipuri de cancer. Tabelul de mai jos oferă exemple pentru care deficiența de reparare a ADN-ului la un cancer s-a dovedit a fi cauzată de o modificare epigenetică și de frecvențele oarecum mai scăzute cu care aceeași deficiență de reparare a ADN-ului cauzat epigenetic a fost găsită în defectul câmpului înconjurător.
| Cancer | genă | Frecvența în cancer | Frecvența în defecte de câmp | Ref. |
|---|---|---|---|---|
| colorectal | MGMT | 46% | 34% | [23] |
| colorectal | MGMT | 47% | 11% | [25] |
| colorectal | MGMT | 70% | 60% | [44] |
| colorectal | MSH2 | 13% | 5% | [25] |
| colorectal | ERCC1 | 100% | 40% | [30] |
| colorectal | PMS2 | 88% | 50% | [30] |
| colorectal | XPF | 55% | 40% | [30] |
| Cap și gât | MGMT | 54% | 38% | [45] |
| Cap și gât | MLH1 | 33% | 25% | [46] |
| Cap și gât | MLH1 | 31% | 20% | [47] |
| Stomac | MGMT | 88% | 78% | [48] |
| Stomac | MLH1 | 73% | 20% | [49] |
| Esofag | MLH1 | 77% -100% | 23% -79% | [50] |
Unii dintre micii polipi din defectul de câmp prezentat în fotografia segmentului de colon deschis pot fi neoplasme relativ benigne. Dintre polipii cu dimensiunea mai mică de 10 mm, găsiți în timpul colonoscopiei și urmați de colonoscopii repetate timp de 3 ani, 25% au fost neschimbați ca mărime, 35% au regresat sau și-au redus dimensiunile, în timp ce 40% au crescut în dimensiuni.[51]
Instabilitatea genomului
[modificare | modificare sursă]Se știe că toate cancerele prezintă o instabilitate a genomului sau un fenotip mutator.[52] ADN-ul care codifică proteinele din nucleu este aproximativ 1,5% din ADN-ul genomic total.[53] În cadrul acestui ADN care codifică proteine (numit <i>exome</i>), un cancer mediu al sânului sau al colonului poate avea aproximativ 60 până la 70 de mutații care modifică proteine, dintre care aproximativ 3 sau 4 pot fi mutații „conducătoare”, iar cele rămase pot fi mutații "pasagere".[42] Cu toate acestea, numărul mediu de mutații ale secvenței de ADN în întregul genom (inclusiv în regiunile care nu codifică proteine) în cadrul unui eșantion de țesut de cancer de sân este de aproximativ 20.000.[54] Într-un eșantion mediu de țesut de melanom (unde melanomele au o frecvență mai mare de mutație a exomeului[42]), numărul total de mutații ale secvenței de ADN este de aproximativ 80.000.[55] A se compara cu frecvența de mutație foarte scăzută, de aproximativ 70 de mutații noi, în întregul genom, între generații (de la părinte la copil) la oameni. [56] [57]
Frecvențele mari ale mutațiilor în secvențele totale de nucleotide din cadrul cancerelor sugerează că, de multe ori, o modificare timpurie a defectelor de câmp care generează un cancer (de exemplu, zona galbenă din diagrama din această secțiune) este o deficiență în repararea ADN-ului.
Defectele mari de câmp care înconjoară cancerele de colon (se extind până la aproximativ 10 cm de fiecare parte a unui cancer) au fost arătate de Facista și colab.[30] ca având frecvent defecte epigenetice în 2 sau 3 proteine de reparare a ADN-ului (ERCC1, XPF sau PMS2) în întreaga zonă a defectului de câmp.
Deficiențele în repararea ADN-ului determină creșterea ratelor de mutație.[31][32][33] O deficiență în repararea ADN-ului, poate permite acumularea de deteriorări ale ADN-ului, iar <i>sinteza de translezie</i> predispusă la erori, trecută prin aceste deteriorări, poate da naștere mutațiilor. În plus, repararea defectuoasă a acestor deteriorări acumulate ale ADN-ului poate duce la epimutații. Aceste noi mutații sau epimutații pot oferi un avantaj proliferativ, generând un defect de câmp. Deși mutațiile / epimutațiile din genele de reparare a ADN-ului nu conferă ele însele un avantaj selectiv, ele pot fi transportate ca pasageri în celule atunci când celulele dobândesc mutații / epimutații suplimentare care oferă un avantaj proliferativ.
Etimologie
[modificare | modificare sursă]Termenul „neoplasm” este sinonim cu „tumoarea”. „Neoplazia” denumește procesul de formare a neoplasmelor / tumorilor, procesul este denumit un proces „neoplazic”. Însuși cuvântul „neoplastic” provine din grecescul neo („nou”) și plastic („format, modelat”).
Termenul „tumoare” derivă din substantivul latin tumor, „umflătură” - derivat al verbului tumēre „a se umfla”.
Referințe
[modificare | modificare sursă]- ^ a b c „Type-2 pericytes participate in normal and tumoral angiogenesis”. Am. J. Physiol., Cell Physiol. 307 (1): C25–38. iulie 2014. doi:10.1152/ajpcell.00084.2014. PMC 4080181
. PMID 24788248.
- ^ Cooper GM (). Elements of human cancer. Boston: Jones and Bartlett Publishers. p. 16. ISBN 978-0-86720-191-8.
- ^ Taylor, Elizabeth J. (). Dorland's Illustrated medical dictionary (ed. 29th). Philadelphia: Saunders. p. 1184. ISBN 978-0721662541.
- ^ Stedman's medical dictionary (ed. 28th). Philadelphia: Lippincott Williams & Wilkins. . p. Neoplasm. ISBN 978-0781733908.
- ^ „II Neoplasms”. International Statistical Classification of Diseases and Related Health Problems 10th Revision (ICD-10) Version for 2010. World Health Organization. Accesat în .
- ^ a b Abrams, Gerald. „Neoplasia I”. Arhivat din original la . Accesat în .
- ^ „Cancer - Activity 1 - Glossary, page 4 of 5”. Arhivat din original la . Accesat în .
- ^ „Clone definition - Medical Dictionary definitions of popular medical terms easily defined on MedTerms”. Arhivat din original la . Accesat în .
- ^ „The association of Epstein-Barr virus with smooth-muscle tumors occurring after organ transplantation”. N. Engl. J. Med. 332 (1): 19–25. ianuarie 1995. doi:10.1056/NEJM199501053320104. PMID 7990861.
- ^ „Tumors in amphibia”. Zoological Science. 4: 411–425. .
- ^ „On the mechanics of a growing tumor”. International Journal of Engineering Science. 40 (12): 1297–316. . doi:10.1016/S0020-7225(02)00014-9.
- ^ Volokh KY (septembrie 2006). „Stresses in growing soft tissues”. Acta Biomater. 2 (5): 493–504. doi:10.1016/j.actbio.2006.04.002. PMID 16793355.
- ^ a b „DNA Damage, DNA Repair and Cancer”. New Research Directions in DNA Repair. . pp. 413–65. doi:10.5772/53919. ISBN 978-953-51-1114-6.
- ^ „DNA damage responses: mechanisms and roles in human disease: 2007 G.H.A. Clowes Memorial Award Lecture”. Mol. Cancer Res. 6 (4): 517–24. . doi:10.1158/1541-7786.MCR-08-0020. PMID 18403632.
- ^ „A novel application of the Margin of Exposure approach: segregation of tobacco smoke toxicants”. Food Chem. Toxicol. 49 (11): 2921–33. noiembrie 2011. doi:10.1016/j.fct.2011.07.019. PMID 21802474.
- ^ „Ultraviolet radiation and melanoma”. Semin Cutan Med Surg. 30 (4): 222–8. decembrie 2011. doi:10.1016/j.sder.2011.08.003. PMID 22123420.
- ^ „Redox biology and gastric carcinogenesis: the role of Helicobacter pylori”. Redox Rep. 16 (1): 1–7. . doi:10.1179/174329211X12968219310756. PMID 21605492.
- ^ „Carcinogenicity of deoxycholate, a secondary bile acid”. Arch. Toxicol. 85 (8): 863–71. august 2011. doi:10.1007/s00204-011-0648-7. PMC 3149672
- ^ „Early-stage formation of an epigenetic field defect in a mouse colitis model, and non-essential roles of T- and B-cells in DNA methylation induction”. Oncogene. 31 (3): 342–51. ianuarie 2012. doi:10.1038/onc.2011.241. PMID 21685942.
- ^ Malkin D (aprilie 2011). „Li-fraumeni syndrome”. Genes Cancer. 2 (4): 475–84. doi:10.1177/1947601911413466. PMC 3135649
- ^ „Environmental and heritable factors in the causation of cancer--analyses of cohorts of twins from Sweden, Denmark, and Finland”. N. Engl. J. Med. 343 (2): 78–85. iulie 2000. doi:10.1056/NEJM200007133430201. PMID 10891514.
- ^ „O(6)-methylguanine methyltransferase in colorectal cancers: detection of mutations, loss of expression, and weak association with G:C>A:T transitions”. Gut. 54 (6): 797–802. iunie 2005. doi:10.1136/gut.2004.059535. PMC 1774551
- ^ a b „MGMT promoter methylation and field defect in sporadic colorectal cancer”. J. Natl. Cancer Inst. 97 (18): 1330–8. septembrie 2005. doi:10.1093/jnci/dji275. PMID 16174854.
- ^ „Promoter methylation status of hMLH1, MGMT, and CDKN2A/p16 in colorectal adenomas”. World J. Gastroenterol. 16 (28): 3553–60. iulie 2010. doi:10.3748/wjg.v16.i28.3553. PMC 2909555
- ^ a b c „Promoter methylation status of hMLH1, hMSH2, and MGMT genes in colorectal cancer associated with adenoma-carcinoma sequence”. Langenbecks Arch Surg. 396 (7): 1017–26. octombrie 2011. doi:10.1007/s00423-011-0812-9. PMID 21706233.
- ^ „Promoter CpG island hypermethylation of the DNA repair enzyme MGMT predicts clinical response to dacarbazine in a phase II study for metastatic colorectal cancer”. Clin. Cancer Res. 19 (8): 2265–72. aprilie 2013. doi:10.1158/1078-0432.CCR-12-3518. PMID 23422094.
- ^ „Different patterns of DNA methylation of the two distinct O6-methylguanine-DNA methyltransferase (O6-MGMT) promoter regions in colorectal cancer”. Mol. Biol. Rep. 40 (5): 3851–7. mai 2013. doi:10.1007/s11033-012-2465-3. PMID 23271133.
- ^ „Immunohistochemical analysis reveals high frequency of PMS2 defects in colorectal cancer”. Gastroenterology. 128 (5): 1160–71. mai 2005. doi:10.1053/j.gastro.2005.01.056. PMID 15887099.
- ^ „Modulation of mismatch repair and genomic stability by miR-155”. Proc. Natl. Acad. Sci. U.S.A. 107 (15): 6982–7. aprilie 2010. doi:10.1073/pnas.1002472107. PMC 2872463
- ^ a b c d e „Deficient expression of DNA repair enzymes in early progression to sporadic colon cancer”. Genome Integr. 3 (1): 3. . doi:10.1186/2041-9414-3-3. PMC 3351028
- ^ a b „Elevated levels of mutation in multiple tissues of mice deficient in the DNA mismatch repair gene Pms2”. Proc. Natl. Acad. Sci. U.S.A. 94 (7): 3122–7. aprilie 1997. doi:10.1073/pnas.94.7.3122. PMC 20332
- ^ a b „Differing patterns of genetic instability in mice deficient in the mismatch repair genes Pms2, Mlh1, Msh2, Msh3 and Msh6”. Carcinogenesis. 27 (12): 2402–8. decembrie 2006. doi:10.1093/carcin/bgl079. PMC 2612936
- ^ a b „Disruption of Brca2 increases the spontaneous mutation rate in vivo: synergism with ionizing radiation”. EMBO Rep. 3 (3): 255–60. martie 2002. doi:10.1093/embo-reports/kvf037. PMC 1084010
- ^ Lee, Jeannie T, ed. (). „Double strand breaks can initiate gene silencing and SIRT1-dependent onset of DNA methylation in an exogenous promoter CpG island”. PLoS Genet. 4 (8): e1000155. doi:10.1371/journal.pgen.1000155. PMC 2491723
- ^ „DNA damage, homology-directed repair, and DNA methylation”. PLoS Genet. 3 (7): e110. iulie 2007. doi:10.1371/journal.pgen.0030110. PMC 1913100
- ^ „Intratumor heterogeneity and branched evolution revealed by multiregion sequencing”. N. Engl. J. Med. 366 (10): 883–92. martie 2012. doi:10.1056/NEJMoa1113205. PMC 4878653
- ^ „Field cancerization in oral stratified squamous epithelium; clinical implications of multicentric origin”. Cancer. 6 (5): 963–8. septembrie 1953. doi:10.1002/1097-0142(195309)6:5<963::AID-CNCR2820060515>3.0.CO;2-Q. PMID 13094644.
- ^ „Field defects in progression to gastrointestinal tract cancers”. Cancer Lett. 260 (1–2): 1–10. februarie 2008. doi:10.1016/j.canlet.2007.11.027. PMC 2744582
- ^ „Deficient Pms2, ERCC1, Ku86, CcOI in field defects during progression to colon cancer”. J Vis Exp (41): 1931. . doi:10.3791/1931. PMC 3149991
- ^ Rubin H (martie 2011). „Fields and field cancerization: the preneoplastic origins of cancer: asymptomatic hyperplastic fields are precursors of neoplasia, and their progression to tumors can be tracked by saturation density in culture”. BioEssays. 33 (3): 224–31. doi:10.1002/bies.201000067. PMID 21254148.
- ^ „Genetic reconstruction of individual colorectal tumor histories”. Proc. Natl. Acad. Sci. U.S.A. 97 (3): 1236–41. februarie 2000. doi:10.1073/pnas.97.3.1236. PMC 15581
- ^ a b c „Cancer genome landscapes”. Science. 339 (6127): 1546–58. martie 2013. doi:10.1126/science.1235122. PMC 3749880
- ^ „Etiologic field effect: reappraisal of the field effect concept in cancer predisposition and progression”. Mod Pathol. 28 (1): 14–29. . doi:10.1038/modpathol.2014.81. PMC 4265316
- ^ „Methylation tolerance due to an O6-methylguanine DNA methyltransferase (MGMT) field defect in the colonic mucosa: an initiating step in the development of mismatch repair-deficient colorectal cancers”. Gut. 59 (11): 1516–26. noiembrie 2010. doi:10.1136/gut.2009.194787. PMID 20947886.
- ^ „Frequent hypermethylation of DAPK, RARbeta, MGMT, RASSF1A and FHIT in laryngeal squamous cell carcinomas and adjacent normal mucosa”. Oral Oncol. 47 (2): 104–7. februarie 2011. doi:10.1016/j.oraloncology.2010.11.006. PMID 21147548.
- ^ „Increased microsatellite instability and epigenetic inactivation of the hMLH1 gene in head and neck squamous cell carcinoma”. Otolaryngol Head Neck Surg. 141 (4): 484–90. octombrie 2009. doi:10.1016/j.otohns.2009.07.007. PMID 19786217.
- ^ „Head and neck squamous cell carcinoma: mismatch repair immunohistochemistry and promoter hypermethylation of hMLH1 gene”. Am J Otolaryngol. 32 (6): 528–36. . doi:10.1016/j.amjoto.2010.11.005. PMID 21353335.
- ^ „Promoter hypermethylation of multiple genes in early gastric adenocarcinoma and precancerous lesions”. Hum. Pathol. 40 (11): 1534–42. noiembrie 2009. doi:10.1016/j.humpath.2009.01.029. PMID 19695681.
- ^ „Promoter methylation status of DNA repair gene (hMLH1) in gastric carcinoma patients of the Kashmir valley”. Asian Pac. J. Cancer Prev. 13 (8): 4177–81. . doi:10.7314/APJCP.2012.13.8.4177. PMID 23098428.
- ^ „Role of epigenetic alterations in the pathogenesis of Barrett's esophagus and esophageal adenocarcinoma”. Int J Clin Exp Pathol. 5 (5): 382–96. . PMC 3396065
- ^ „Growth of colorectal polyps: redetection and evaluation of unresected polyps for a period of three years”. Gut. 39 (3): 449–56. septembrie 1996. doi:10.1136/gut.39.3.449. PMC 1383355
- ^ „Implications of genetic heterogeneity in cancer”. Ann. N. Y. Acad. Sci. 1267: 110–6. septembrie 2012. doi:10.1111/j.1749-6632.2012.06590.x. PMC 3674777
- ^ „Initial sequencing and analysis of the human genome”. Nature. 409 (6822): 860–921. februarie 2001. doi:10.1038/35057062. PMID 11237011.
- ^ „Identification of high-confidence somatic mutations in whole genome sequence of formalin-fixed breast cancer specimens”. Nucleic Acids Res. 40 (14): e107. august 2012. doi:10.1093/nar/gks299. PMC 3413110
- ^ „Melanoma genome sequencing reveals frequent PREX2 mutations”. Nature. 485 (7399): 502–6. mai 2012. doi:10.1038/nature11071. PMC 3367798
- ^ „Analysis of genetic inheritance in a family quartet by whole-genome sequencing”. Science. 328 (5978): 636–9. aprilie 2010. doi:10.1126/science.1186802. PMC 3037280
- ^ „Estimating the human mutation rate using autozygosity in a founder population”. Nat. Genet. 44 (11): 1277–81. noiembrie 2012. doi:10.1038/ng.2418. PMC 3483378