Microsporidia
| Microsporidiomycota Microsporidia | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Classificação científica | |||||||||||
| |||||||||||
| Classes e ordens[2] | |||||||||||
| |||||||||||
| Sinónimos | |||||||||||
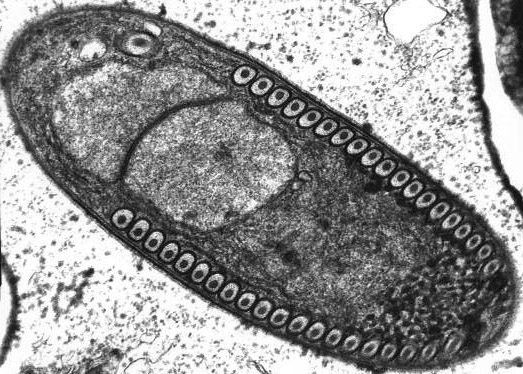
Microsporidia é um clado de organismos filogeneticamente próximos dos fungos, entre os quais se encontra classificado ao nível de divisão.[7] Com distribuição cosmopolita, todos os membros conhecidos são parasitas intracelulares obrigatórios de organismos eucarióticos. Estão descritas cerca de 1300 espécies, repartidas por cerca de 160 géneros, o que é uma pequena parte da diversidade real do grupo, uma vez que um grande número de potenciais hospedeiros não foi examinado para infecção por microsporídios. Esses patógenos são comuns entre animais de quase todos os grupos sistemáticos, desde protozoários até vertebrados superiores, incluindo humanos, nos quais são causadores de diversas formas de microsporidiose. Os mais numerosos e diversos são os microsporídios que parasitam crustáceos e insectos.[8]
Descrição
[editar | editar código-fonte]Definição do grupo
[editar | editar código-fonte]A divisão Microsporidia tem sofrido profundas modificações taxonómicas nas últimas décadas, não existindo consenso pleno quanto à sua taxonomia interna. O grupo era inicialmente um conjunto de parasitas unicelulares de animais, formadores de esporos e de estrutura muito simples, pelo que foi incluído entre os protozoários. Contudo, após estudos genéticos, descobriu-se que eram organismos próximos dos fungos, principalmente dos fungos do antigo filo Zygomycota, o que motivou a sua transferência para a actual posição taxonómica.[9]
Os termos «microsporidium» (pl. «microsporidia») e «microsporídios» são usados como nomes vernaculares para os membros do grupo. O nome Microsporidium Balbiani, 1884[10] também é usado como um nome genérico para membros incertae sedis do agrupamento.[11]
Características gerais
[editar | editar código-fonte]Os microsporídios possuem uma série de características únicas que demonstram o extremo grau de especialização deste táxon para o parasitismo intracelular. Os esporos desses patógenos contêm um complexo único de organelas, conhecido apenas por microsporídios, um aparato de extrusão projetado para infectar a célula hospedeira perfurando sua membrana e lançando o embrião diretamente no citoplasma. Este método de transferência do embrião do esporo para a célula do animal hospedeiro não é conhecido para outros protozoários.[8]
O genoma dos microsporídios é o menor entre os eucariotos e praticamente carece de íntrons, sendo neles a mitose uma forma de pleuromitose intranuclear fechada. Os seus ribossomas são semelhantes aos dos organimos procarióticos e possuem um coeficiente de sedimentação de 70S. A célula dos microsporídios carece de cinetossomas e seus derivados (flagelos e centríolos), lisossomas e reserva de grânulos de nutrientes. Por muito tempo, a ausência de mitocôndrias nos microsporídios foi considerada primária, mas recentemente pequenos organelos de duas membranas, os mitossomas, foram encontradas nas suas células, e a sua origem mitocondrial foi comprovada.[12]
Morfologia e fisiologia
[editar | editar código-fonte]
Os Microsporidia são um grupo de parasitas intracelulares unicelulares formadores de esporos. Não apresentam mitocôndrias, tendo em seu lugar mitossomas. Também são desprovidos de estruturas móveis, como flagelos. Estas características levaram a que fossem considerados protozoários ou protistas, mas agora são reconhecidos como fungos,[14] ou como um grupo irmão dos fungos.[15]
Esses micróbios fúngicos são parasitas eucarióticos obrigatórios que usam um mecanismo único para infectar as células hospedeiras.[16] Comprovou-se recentemente, com um estudo publicado em 2017, que o grupo infecta coleópteros em grande escala. Até agora, foram validamente descritas cerca de 1500 espécies de um total de provavelmente mais de um milhão de espécies extantes.[17]
A distribuição dos microsporídios está restrita à distribuição dos animais hospedeiros, e todos os principais grupos taxonómicos de animais hospedam microsporídios. A maioria infecta insetoss, mas também são responsáveis por doenças comuns de crustáceos e peixes. As espécies conhecidas de microsporídios geralmente infectam apenas uma espécie hospedeira ou um grupo de táxons intimamente relacionados. Aproximadamente 10 % das espécies são parasitas de vertebrados e várias espécies, a maioria das quais são oportunistas, podem infectar humanos, nos quais podem causar microsporidiose.
Após a infecção, os microsporídios influenciam os seus hospedeiros de várias maneiras e todos os órgãos e tecidos podem ser invadidos, embora geralmente por diferentes espécies de microsporídios especializados. Algumas espécies são letais e algumas são usadas no controle biológico de insetos-praga. Nos insectos, a castração parasitária, o gigantismo ou a mudança do sexo do hospedeiro são efeitos potenciais do parasitismo microsporidiano. Nos casos mais avançados de parasitismo, o microsporídio domina completamente a célula hospedeira e controla o seu metabolismo e reprodução, formando um xenoma.[18]
Esporos
[editar | editar código-fonte]A replicação ocorre dentro das células do hospedeiro, que são infectadas por meio de esporos unicelulares. Estes variam de 1 a 40 μm de comprimento, o que os coloca entre os menores eucariontes conhecidos. Os microsporídios que infectam mamíferos têm comprimentos de 1,0 a 4,0 μm.[19] O grupo também apresenta os menores genomas eucarióticos conhecidos.
Os esporos, altamente resistentes e capazes de sobreviver fora do hospedeiro por vários anos, apresentam um aparato de extrusão, destinado a facilitar a sua saída da célula do hospedeiro, constituído por um tubo polar enrolado que termina num disco de ancoragem na parte apical do esporo.[20] A morfologia dos esporos é útil na distinção entre diferentes espécies. Os esporos da maioria das espécies são ovais ou piriformes, mas esporos em forma de bastonete ou esféricos não são incomuns. Algumas gerações produzem esporos de forma única para o género. O esporo é protegido por uma parede, composta por três camadas: (1) um exósporo externo, com uma camada densa; (2) um endósporo mediano, largo e aparentemente sem estrutura, contendo quitina; e (3) uma fina membrana plasmática interna.
Na maioria dos casos existem no esporo dois [[núcleo celular|núcleos celulares] intimamente associados, formando um diplokaryon, mas por vezes há apenas um núcleo.
A metade anterior do esporo contém um aparelho semelhante a um arpão, com um longo filamento polar em forma de fio, que é enrolado na metade posterior do esporo. A parte anterior do filamento polar é circundada por um polaroplasto, uma lamela de membranas. Atrás do filamento polar existe um vacúolo posterior.[18]
Genoma
[editar | editar código-fonte]Os microsporídios têm os menores genomas eucarióticos (nucleares) conhecidos. O estilo de vida parasitário dos microsporídios levou à perda de muitos genes mitocondriais e do aparelho de Golgi, e até mesmo o seu RNA ribossómico está reduzido em tamanho em comparação com os da maioria dos eucariontes.
Como consequência, os genomas dos microsporídios são muito menores do que os de outros eucariotas. Os genomas de microsporídios atualmente conhecidos têm tamanho de 2,5 a 11,6 Mb, codificando de 1848 a 3266 proteínas, o que está na mesma faixa de muitas bactérias.[21]
Eventos de transferência horizontal de genes (HGT) parecem ter ocorrido muitas vezes no microsporídios. Por exemplo, os genomas de Encephalitozoon romaleae e Trachipleistophora hominis contêm genes que derivam de animais e bactérias, e alguns até de fungos.[21]
Infecção e seus efeitos
[editar | editar código-fonte]No caso da infecção de hospedeiros multicelulares com estrutura digestiva diferenciada, quando o esporo atinge o intestino do hospedeiro germina e aumenta a pressão osmótica no seu interior até que a sua parede rígida se rompe no ponto mais fino, situado no ápice. Simultaneamente, o material do filamento é rearranjado para formar um tubo que funciona como uma agulha hipodérmica e penetra no epitélio intestinal. Formada a estrutura de injecção, o vacúolo posterior incha, forçando o filamento polar a ejetar rapidamente o conteúdo infeccioso para o citoplasma do hospedeiro potencial.
Uma vez dentro da célula hospedeira, forma-se um esporoplasma, dividindo-se ou formando um plasmódio multinucleado antes de produzir novos esporos. O ciclo de vida varia consideravelmente. Alguns têm um ciclo de vida assexuado simples,[22] enquanto outros têm um ciclo de vida complexo, envolvendo múltiplos hospedeiros e reprodução assexuada e reprodução sexual.
Diferentes tipos de esporos podem ser produzidos em diferentes estágios, provavelmente com diferentes funções, incluindo autoinfecção (transmissão dentro de um único hospedeiro).
Implicações para a saúde do hospedeiro
[editar | editar código-fonte]Nos hospediros animais, incluindo os humanos, os microsporídios geralmente causam doenças crónicas e debilitantes, em vez de infecções letais. Os efeitos sobre o hospedeiro incluem redução da longevidade, fertilidade, peso e vigor geral. A transmissão vertical de microsporídios é frequentemente relatada.
No caso dos cinsetos, a transmissão vertical geralmente ocorre como transmissão transovariana, onde os parasitas microsporídeos passam dos ovários da fêmea hospedeira para os ovos e eventualmente se multiplicam nas larvas infectadas. As espécies Amblyospora salinaria, que infecta o mosquito Culex salinarius Coquillett, e Amblyospora californica, que infecta o mosquito Culex tarsalis Coquillett, fornecem exemplos típicos de transmissão transovariana de microsporídios.[23][24][25][26]
As infecções de humanos por microsporídios por vezes causam uma doença chamada microsporidiose. Pelo menos 14 espécies de microsporídeos, distribuídas por oito géneros, foram reconhecidas como patógenos humanos. Estes incluem Trachipleistophora hominis, o patógeno humano mais comum.[27]
Controlo da malária
[editar | editar código-fonte]Membros do agrupamento Microsporidia, especificamente a espécie Vavraia culicis, que infecta mosquitos, está a ser testada como um possível método de controle da malária 'à prova de evolução'.[28]
A infecção de Anopheles gambiae, o principal vetor da malária Plasmodium falciparum, por microsporídeos reduz a infecção por malária dentro do mosquito e encurta a sua vida útil.[29]
Como a maioria dos mosquitos infectados com malária morre naturalmente antes que o parasita da malária esteja maduro o suficiente para transmitir a doença, qualquer aumento na mortalidade do mosquito por meio da infecção por microsporídios pode reduzir a transmissão da malária para humanos. Em maio de 2020, pesquisadores relataram que Microsporidia MB, um simbionte no intestino médio e ovários de Anopheles arabiensis, prejudicava significativamente a transmissão de P. falciparum, não teve "nenhum efeito evidente" na aptidão dos mosquitos hospedeiros e sendo transmitido verticalmente (por herança).[30]
Hiperparasitismo
[editar | editar código-fonte]
Os Microsporidia podem infectar uma grande variedade de hospedeiros, incluindo hospedeiros que são eles próprios parasitas. Nesse caso, a espécie microsporídica é um hiperparasita, ou seja, um parasita de um parasita. Como exemplo, são conhecidas mais de dezoito espécies que parasitam digeneanos (parasitas platelmintas). Esses organismos são eles próprios parasitas de vários vertebrados e moluscos. Oito dessas espécies pertencem ao género Nosema.[31]
Da mesma forma, a espécie microsporídica Toguebayea baccigeri é um parasita de um Digenea, o faustulídeo Bacciger israelensis, ele próprio um parasita intestinal de um peixe marinho, a boga Boops boops (Teleostei, Sparidae).[32]
Classificação e filogenia
[editar | editar código-fonte]O primeiro género microsporidiano descrito, Nosema, foi inicialmente colocado por Carl Nägeli no grupo de fungos Schizomycetes, juntamente com algumas bactérias e leveduras.[33][34] Por algum tempo os microsporídios foram considerados eucariotas muito primitivos, colocados no grupo dos protozoários Cnidospora.[1]
Mais tarde, principalmente pela ausência de mitocôndrias, os membros do agrupamento Microsporidia foram colocados, em conjunto com outros Protozoa como os diplomonados, parabasalídeos e Archamoebae, no grupo de protozoários Archezoa, organismos que terão divergido antes da origem da mitocôndria.[35] Investigação mais recente demonstrou que era falsa a teoria da origem primitiva (para todos eles). Em vez disso, os microsporídios são propostos como organismos altamente desenvolvidos e especializados, que apenas dispensaram funções biológicas que deixaram de ser necessárias, porque são fornecidas pelo hospedeiro.[36] Além disso, os organismos formadores de esporos em geral têm um sistema complexo de reprodução, tanto sexual quanto assexuado, que parece longe de ser primitivo.
Desde meados da década de 2000, os microsporídios foram colocados dentro do reino Fungi ou, em alternativa, considerados como um grupo irmão dos Fungi partilhando um ancestral comum.[37][38][39][40] Tendo em conta a grande diversidade de organismos, o facto de serem microscópicos, ubíquos e, na sua vasta maioria, parasitas obrigatórios, durante muito tempo, o trabalho para identificar os clados foi amplamente baseado no habitat e no hospedeiro. Nesse contexto, foram propostas três classes de Microsporidia por Vossbrinck e Debrunner-Vossbrinck, com base no habitat: Aquasporidia, Marinosporidia e Terresporidia.[41] A partir de 2007 o clado foi integrado como uma divisão (ou filo) no reino Fungi,[7] solução que tem vindo a ganhar consenso entre os micologistas.
Uma possível árvore filogenética para o grupo, incluindo outros grupos de fungos e opistocontos, é a seguinte:[42][43][44]
| Opisthokonta |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
O status nomenclatural de Microsporidia é ambíguo. O clado foi tratado como um filo sob as Código Internacional de Nomenclatura Zoológica (ICZN), pois ao tempo o agrupamento estava atribuído a Sporozoa, mas a citação do autor não é clara. Também é questionável se o nome é válido de acordo com o Código Internacional de Nomenclatura Botânica (ICN), que passou a aplicar-se por força da transferência para o reino Fungi, pois a filiação dos Microsporidia entre os fungos e, portanto, também a responsabilidade do ICN, apenas surgiu em 2007. A atribuição da autoridade ao embriologista francês Édouard-Gérard Balbiani[45] é, portanto, provisória,[7] embora de uso comum na actualidade.
Uma nova classificação, proposta por Cavalier-Smith em 1993, incluía os seguintes agrupamentos de alto nível taxonómico:[46]
- Subfilo Rudimicrospora Cavalier-Smith 1993
- Classe Minisporea Cavalier-Smith 1993
- Ordem Minisporida Sprague, 1972
- Classe Metchnikovellea Weiser, 1977
- Ordem Metchnikovellida Vivier, 1975
- Classe Minisporea Cavalier-Smith 1993
- Subfilo Polaroplasta Cavalier-Smith 1993
- Classe Pleistophoridea Cavalier-Smith 1993
- Ordem Pleistophorida Stempell 1906
- Classe Disporea Cavalier-Smith 1993
- Subclasse Unikaryotia Cavalier-Smith 1993
- Subclasse Diplokaryotia Cavalier-Smith 1993
- Classe Pleistophoridea Cavalier-Smith 1993
Com o advento da filogenética molecular foi possível estabelecer sistemas de classificação independentes do habitat, do hospedeiro, da morfologia ou da ecologia das espécies integrantes. Para os Microsporidia surgiu uma primeira classificação de base essencialmente molecular em 2007, da autoria de Aleksandr Fedorovich Alimov (1933-2019), revista em 2020 por um numeroso grupo de micologistas liderado por Nalin N. Wijayawardene. As duas classificações são apresentadas no quadro seguinte.
Quadro comparativo entre as classificações de Alimov 2007 e de Wijayawardene et al. 2020
|
Ver também
[editar | editar código-fonte]- Lista de géneros de Microsporidia
- Glugea, um género de microsporídios
- Nosema apis, um microsporídio parasita de abelhas
Referências
[editar | editar código-fonte]- ↑ a b Corliss JO, Levine ND (1963). «Establishment of the Microsporidea as a new class in the protozoan subphylum Cnidospora». The Journal of Protozoology. 10 (Suppl): 26–27. doi:10.1111/jeu.1963.10.issue-s3
- ↑ Wijayawardene, N.N.; Hyde, K.D.; Dai, D.Q.; Sánchez-García, M.; Goto, B.T.; Saxena, R.K.; et al. (2022). «Outline of Fungi and fungus-like taxa – 2021». Mycosphere. 13 (1): 53–453. doi:10.5943/mycosphere/13/1/2
- ↑ Balbiani, G (1882). «Sur les microsporidies ou psorospermies des Articulés». C. R. Acad. Sci. 95: 1168–71
- ↑ Delphy, J. 1936. Sous-règne des Protozoaires. In: Perrier, R. (ed.). La Faune de la France en tableaux synoptiques illustrés, vol 1A. Delagrave: Paris.
- ↑ Levine, N. D.; et al. (1980). «A Newly Revised Classification of the Protozoa». The Journal of Protozoology. 27 (1): 37–58. PMID 6989987. doi:10.1111/j.1550-7408.1980.tb04228.x
- ↑ Sprague, V. (1977). Classification and phylogeny of the Microsporidia. In: Comparative pathobiology. vol. 2, Systematics of the Microsporidia. Lee A. Bulla & Thomas C. Cheng (ed.). pp. 1-30. New York: Plenum Press, [1].
- ↑ a b c D. S. Hibbett et al.: A higher-level phylogenetic classification of the Fungi. In: Mycological research, Mai 2007; 111(5): 509-547. Epub 2007 13. März 2007. PMID 17572334
- ↑ a b Wittner M., Weiss L. M. The microsporidia and microsporidiosis. — Washington (D. C.): ASM Press, 1999.
- ↑ Fischer WM, Palmer JD (2005). «Evidence from small-subunit ribosomal RNA sequences for a fungal origin of Microsporidia». Molecular Phylogenetics and Evolution. 36 (3): 606–22. PMID 15923129. doi:10.1016/j.ympev.2005.03.031
- ↑ Balbiani, G. 1884. Les Psorospermies des Articulés ou Microsporidies, pp. 150-168, 184. In: Leçons sur les sporozoaires. Paris: Doin, [2].
- ↑ Hoffman, G. (1999). Parasites of North American Freshwater Fishes, 2nd edn, University of California Press, Berkeley, California, USA, p. 89, [3].
- ↑ Keeling P., Five Questions about Microsporidia, PLoS Pathogens. vol. 5, 2009, n.º 9, pp. 1—3. (doi:10.1371/journal.ppat.1000489).
- ↑ Winters, A. D.; Faisal, M. (2014). «Molecular and ultrastructural characterization of Dictyocoela diporeiae n. sp. (Microsporidia), a parasite of Diporeia spp. (Amphipoda, Gammaridea)». Parasite. 21: 26. PMC 4059264
. PMID 24934702. doi:10.1051/parasite/2014028
- ↑ Hibbett, D.S.; et al. (2007). «A higher level phylogenetic classification of the Fungi» (PDF). Mycological Research. 111 (5): 509–47. PMID 17572334. doi:10.1016/j.mycres.2007.03.004
- ↑ Silar, Philippe (2016). Protistes Eucaryotes : Origine, Evolution et Biologie des Microbes Eucaryotes. [S.l.]: HAL. p. 462. ISBN 978-2-9555841-0-1
- ↑ Franzen, C. (2005). How do Microsporidia invade cells?. Folia Parasitologica, 52(1-2), 36-40. doi.org/10.14411/fp.2005.005
- ↑ Hawksworth, David (2001). «The magnitude of fungal diversity: The 1.5 million spices estimate revisited». Mycological Research. 105 (12): 1422. doi:10.1017/S0953756201004725
- ↑ a b Ronny Larsson, Lund University (Department of Cell and Organism Biology) Cytology and taxonomy of the microsporidia Arquivado em 2009-09-12 no Wayback Machine 2004.
- ↑ Didier, ES. (abril de 2005). «Microsporidiosis: an emerging and opportunistic infection in humans and animals». Acta Trop. 94 (1): 61–76. PMID 15777637. doi:10.1016/j.actatropica.2005.01.010
- ↑ Franzen, C. (2005). How do Microsporidia invade cells?. Folia Parasitologica, 52(1-2), 36-40. doi.org/10.14411/fp.2005.005
- ↑ a b Corradi, N.; Selman, M. (2013). «Latest Progress in Microsporidian Genome Research». Journal of Eukaryotic Microbiology. 60 (3): 309–312. PMID 23445243. doi:10.1111/jeu.12030
- ↑ Ironside JE (2007). «Multiple losses of sex within a single genus of Microsporidia». BMC Evolutionary Biology. 7. 48 páginas. PMC 1853083
- ↑ Andreadis TG, Hall DW (agosto de 1979). «Development, ultrastructure, and mode of transmission of Amblyospora sp. (Microspora) in the mosquito». The Journal of Protozoology. 26 (3): 444–52. PMID 536933. doi:10.1111/j.1550-7408.1979.tb04651.x
- ↑ Andreadis TG, Hall DW (setembro de 1979). «Significance of transovarial infections of Amblyospora sp. (Microspora:Thelohaniidae) in relation to parasite maintenance in the mosquito Culex salinarius». Journal of Invertebrate Pathology. 34 (2): 152–7. PMID 536610. doi:10.1016/0022-2011(79)90095-8
- ↑ Jahn GC, Hall DW, Zam SG (1986). «A comparison of the life cycles of two Amblyospora (Microspora: Amblyosporidae) in the mosquitoes Culex salinarius and Culex tarsalis Coquillett». Journal of the Florida Anti-Mosquito Association. 57 (1): 24–27
- ↑ Becnel JJ, Andreadis TG (maio de 1998). «Amblyospora salinaria n. sp. (Microsporidia: amblyosporidae), parasite of Culex salinarius (Diptera: culicidae): its life cycle stages in an intermediate host». Journal of Invertebrate Pathology. 71 (3): 258–62. PMID 9538031. doi:10.1006/jipa.1998.4729
- ↑ Heinz, E; Williams, TA; Nakjang, S; et al. (outubro de 2012). «The genome of the obligate intracellular parasite Trachipleistophora hominis: New insights into microsporidian genome dynamics and reductive evolution». PLOS Pathog. 8 (10): e1002979. PMC 3486916
- ↑ Koella, Jacob C.; Lorenz, Lena; Bargielowski, Irka (2009). Chapter 12 Microsporidians as Evolution-Proof Agents of Malaria Control?. Col: Advances in Parasitology. 68. [S.l.: s.n.] pp. 315–327. ISBN 978-0-12-374787-7. PMID 19289199. doi:10.1016/S0065-308X(08)00612-X
- ↑ Bargielowski I, Koella JC (2009). Baylis M, ed. «A Possible Mechanism for the Suppression of Plasmodium berghei Development in the Mosquito Anopheles gambiae by the Microsporidian Vavraia culicis». PLOS ONE. 4 (3): e4676. Bibcode:2009PLoSO...4.4676B. PMC 2651578
- ↑ Herren, JK; Mbaisi, L; Mararo, E; et al. (2020). «A microsporidian impairs Plasmodium falciparum transmission in Anopheles arabiensis mosquitoes». Nature Communications. 11 (2187): 2187. Bibcode:2020NatCo..11.2187H. PMC 7198529
- ↑ a b Toguebaye, B. S.; Quilichini, Y.; Diagne, P. M.; Marchand, B. (2014). «Ultrastructure and development of Nosema podocotyloidis n. sp. (Microsporidia), a hyperparasite of Podocotyloides magnatestis (Trematoda), a parasite of Parapristipoma octolineatum (Teleostei)». Parasite. 21: 44. PMC 4150386
- ↑ Miquel, Jordi; Kacem, Hichem; Baz-González, Edgar; Foronda, Pilar; Marchand, Bernard (2022). «Ultrastructural and molecular study of the microsporidian Toguebayea baccigeri n. gen., n. sp., a hyperparasite of the digenean trematode Bacciger israelensis (Faustulidae), a parasite of Boops boops (Teleostei, Sparidae)». EDP Sciences. Parasite. 29: 2. ISSN 1776-1042. PMC 8805611
- ↑ Nägeli, C. von (1857). «Über die neue Krankheit der Seidenraupe und verwandte Organismen. pp. 760–61. In: Caspary, R. (ed.). Bericht über die Verhandlungen der 33. Versammlung deutscher Naturforscher und Aerzte, gehalten in Bonn von 18 bis 24 September 1857». Botanische Zeitung. 15: 749–776
- ↑ Keeling, P. J.; Fast, N. M. (2002). «Microsporidia: biology and evolution of highly reduced intracellular parasites» (PDF). Annual Review of Microbiology. 56 (1): 93–116. PMID 12142484. doi:10.1146/annurev.micro.56.012302.160854
- ↑ Cavalier-Smith, T (1993). «Kingdom protozoa and its 18 phyla». Microbiological Reviews. 57 (4): 953–994. PMC 372943
- ↑ Keeling PJ, Slamovits CH (dezembro de 2004). «Simplicity and Complexity of Microsporidian Genomes». Eukaryotic Cell. 3 (6): 1363–9. PMC 539024
- ↑ Fischer WM, Palmer JD (setembro de 2005). «Evidence from small-subunit ribosomal RNA sequences for a fungal origin of Microsporidia». Molecular Phylogenetics and Evolution. 36 (3): 606–22. PMID 15923129. doi:10.1016/j.ympev.2005.03.031
- ↑ Liu YJ, Hodson MC, Hall BD (2006). «Loss of the flagellum happened only once in the fungal lineage: phylogenetic structure of Kingdom Fungi inferred from RNA polymerase II subunit genes». BMC Evolutionary Biology. 6. 74 páginas. PMC 1599754
- ↑ Gill EE, Fast NM (junho de 2006). «Assessing the microsporidia-fungi relationship: Combined phylogenetic analysis of eight genes». Gene. 375: 103–9. PMID 16626896. doi:10.1016/j.gene.2006.02.023
- ↑ Lee SC, Corradi N, Byrnes EJ, et al. (novembro de 2008). «Microsporidia evolved from ancestral sexual fungi». Current Biology. 18 (21): 1675–9. PMC 2654606
- ↑ Vossbrinck CR, Debrunner-Vossbrinck BA (Maio de 2005). «Molecular phylogeny of the Microsporidia: ecological, ultrastructural and taxonomic considerations». Folia Parasitologica. 52 (1–2): 131–42; discussion 130. PMID 16004372. doi:10.14411/fp.2005.017
- ↑ Tedersoo, L., Sánchez-Ramírez, S., Kõljalg, U. et al. 2018, High-level classification of the Fungi and a tool for evolutionary ecological analyses Fungal Diversity 90: 135. https://doi.org/10.1007/s13225-018-0401-0
- ↑ Guifré Torruella et al. 2015, Phylogenomics Reveals Convergent Evolution of Lifestyles in Close Relatives of Animals and Fungi. Current Biology ISSN 0960-9822 Volume 25, Issue 18, p2404–2410, 21 September 2015
- ↑ Spatafora, Joseph W.; Chang, Ying; Benny, Gerald L.; Lazarus, Katy; Smith, Matthew E.; Berbee, Mary L.; Bonito, Gregory; Corradi, Nicolas; Grigoriev, Igor; Gryganskyi, Andrii; James, Timothy Y.; O’Donnell, Kerry; Roberson, Robert W.; Taylor, Thomas N.; Uehling, Jessie; Vilgalys, Rytas; White, Merlin M.; Stajich, Jason E. (2016). «A phylum-level phylogenetic classification of zygomycete fungi based on genome-scale data». Mycologia. 108 (5): 1028–1046. ISSN 0027-5514. PMC 6078412
- ↑ C. R. Acad. Sci Paris 95: S. 1168, 1882.
- ↑ Cavalier-Smith (1993). «Kingdom Protozoa and its 18 phyla». Microbiological Reviews. 57 (4): 953–94. PMC 372943
- ↑ Alimov, A. F., ed. (Maio de 2007). Protista 2: Handbook on zoology. [S.l.]: Nauka. 1141 páginas. ISBN 9785020262249
- ↑ Wijayawardene NN, Hyde KD, Al-Ani LK, Tedersoo L, Haelewaters D, Rajeshkumar KC, et al. (2020). «Outline of Fungi and fungus-like taxa» (PDF). Mycosphere. 11 (1): 1060–1456. ISSN 2077-7019. doi:10.5943/mycosphere/11/1/8
Ligações externas
[editar | editar código-fonte] Dados relacionados com Microsporidia no Wikispecies
- BioHealthBase Bioinformatics Resource Center Database of microspordia sequences and related information.
- MeSH Microsporidia