Taxonomia humana
| Homo (humanos) | |
|---|---|
| |
| Classificação científica | |
| Domínio: | Eukaryota |
| Reino: | Animalia |
| Filo: | Chordata |
| Classe: | Mammalia |
| Ordem: | Primates |
| Subordem: | Haplorhini |
| Infraordem: | Simiiformes |
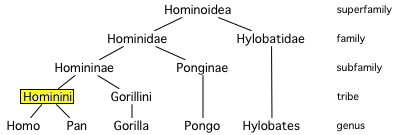
| Família: | Hominidae |
| Subfamília: | Homininae |
| Tribo: | Hominini |
| Gênero: | Homo Linnaeus, 1758 |
| Espécie-tipo | |
| Homo sapiens Linnaeus, 1758
| |
| Espécie | |
|
Outras espécies e subespécies já foram sugeridas | |
| Sinónimos | |
|
Sinônimos
| |

A taxonomia humana é a classificação da espécie humana (Homo sapiens, do latim "homem sábio") dentro da taxonomia zoológica. O gênero Homo foi projetado para incluir tanto os humanos anatomicamente modernos quanto as variedades extintas de humanos arcaicos. Os seres humanos atuais foram designados como subespécie Homo sapiens sapiens, diferenciada, de acordo com alguns, do ancestral direto, Homo sapiens idaltu (com algumas outras pesquisas classificando os idaltu e os seres humanos atuais como pertencentes à mesma subespécie).[1][2][3]
Desde a introdução de nomes sistemáticos no século XVIII, o conhecimento da evolução humana aumentou drasticamente, e vários táxons intermediários foram propostos no século XX e no início do século XXI. O agrupamento taxonômico mais amplamente aceito considera o gênero Homo como originário de dois a três milhões de anos atrás, dividido em pelo menos duas espécies, o Homo erectus arcaico e o Homo sapiens moderno, com cerca de uma dúzia de outras sugestões de espécies sem reconhecimento universal.
O gênero Homo é colocado na tribo Hominini ao lado de Pan (chimpanzés). Estima-se que os dois gêneros tenham divergido durante um longo período de hibridização, abrangendo cerca de 10 a 6 milhões de anos atrás, com possível miscigenação até 4 milhões de anos atrás. Uma subtribo de validade incerta, que agrupa espécies arcaicas "pré-humanas" ou "para-humanas" mais jovens do que a separação Homo-Pan, é a Australopithecina (proposta em 1939).
Uma proposta de Wood e Richmond (2000) introduziria Hominina como uma subtribo ao lado de Australopithecina, sendo Homo o único gênero conhecido dentro de Hominina. Como alternativa, seguindo Cela-Conde e Ayala (2003), os gêneros "pré-humanos" ou "proto-humanos" Australopithecus, Ardipithecus, Praeanthropus e, possivelmente, Sahelanthropus podem ser colocados em pé de igualdade com o gênero Homo. Uma visão ainda mais extrema rejeita a divisão de Pan e Homo como gêneros separados, o que, com base no Princípio da Prioridade, implicaria a reclassificação dos chimpanzés como Homo paniscus (ou similar).[4]
A categorização dos seres humanos com base em fenótipos é um assunto socialmente controverso. Originalmente, os biólogos classificavam as raças como subespécies, mas os antropólogos contemporâneos rejeitam o conceito de raça como uma ferramenta útil para entender a humanidade e, em vez disso, veem a humanidade como um continuum genético complexo e inter-relacionado. A taxonomia dos hominídeos continua a evoluir.[5][6]
História
[editar | editar código-fonte]A taxonomia humana, por um lado, envolve a colocação dos seres humanos na taxonomia dos hominídeos (grandes símios) e, por outro, a divisão dos seres humanos arcaicos e modernos em espécies e, se aplicável, subespécies. A taxonomia zoológica moderna foi desenvolvida por Carl Linnaeus entre as décadas de 1730 e 1750. Ele foi o primeiro a desenvolver a ideia de que, assim como outras entidades biológicas, grupos de pessoas também poderiam compartilhar classificações taxonômicas.[7] Ele nomeou a espécie humana como Homo sapiens em 1758, como a única espécie membro do gênero Homo, dividida em várias subespécies correspondentes às grandes raças. O substantivo latino homō (genitivo hominis) significa "ser humano". O nome sistemático Hominidae para a família dos grandes símios foi introduzido por John Edward Gray (1825).[8] Gray também apresentou Hominini como o nome da tribo que inclui os chimpanzés (gênero Pan) e os humanos (gênero Homo).
A descoberta da primeira espécie humana arcaica extinta a partir de registros fósseis data de meados do século XIX: Homo neanderthalensis, classificado em 1864. Desde então, várias outras espécies arcaicas foram nomeadas, mas não há consenso universal quanto ao seu número exato. Após a descoberta do H. neanderthalensis, que, mesmo sendo "arcaico", é reconhecível como claramente humano, a antropologia do final do século XIX e início do século XX ocupou-se por algum tempo em encontrar o suposto "elo perdido" entre o Homo e o Pan. A farsa do "Homem de Piltdown" de 1912 foi a apresentação fraudulenta dessa espécie de transição. Desde meados do século XX, o conhecimento sobre o desenvolvimento dos hominídeos tornou-se muito mais detalhado, e a terminologia taxonômica foi alterada várias vezes para refletir isso.
A introdução do Australopithecus como um terceiro gênero, ao lado do Homo e do Pan, na tribo Hominini deve-se a Raymond Dart (1925). Australopithecina como uma subtribo contendo Australopithecus e Paranthropus (Broom 1938) é uma proposta de Gregory & Hellman (1939). Mais recentemente, foram propostas adições à subtribo Australopithecina, incluindo Ardipithecus (1995) e Kenyanthropus (2001). A posição do Sahelanthropus (2002) em relação ao Australopithecina dentro dos Hominini não é clara. Cela-Conde e Ayala (2003) propõem o reconhecimento de Australopithecus, Ardipithecus, Praeanthropus e Sahelanthropus (este último incertae sedis) como gêneros separados.[9]
Outros gêneros propostos, agora considerados em sua maioria como parte do Homo, incluem: Pithecanthropus (Dubois, 1894), Protanthropus (Haeckel, 1895), Sinanthropus (Black, 1927), Cyphanthropus (Pycraft, 1928), Africanthropus (Dreyer, 1935),[10] Telanthropus (Broom & Anderson 1949), Atlanthropus (Arambourg, 1954), Tchadanthropus (Coppens, 1965).
O gênero Homo foi considerado originário de cerca de dois milhões de anos atrás, desde a descoberta de ferramentas de pedra em Garganta de Olduvai, na Tanzânia, na década de 1960. O Homo habilis (Leakey et al., 1964) seria a primeira espécie "humana" (membro do gênero Homo) por definição, sendo seu espécime tipo o fóssil OH 7. Entretanto, a descoberta de mais fósseis desse tipo abriu o debate sobre a delimitação do H. habilis do Australopithecus. Em especial, o fóssil de mandíbula LD 350-1 descoberto em 2013, datado de 2,8 milhões de anos, foi considerado como sendo de transição entre os dois.[11] Também é debatido se o H. habilis foi o primeiro hominídeo a usar ferramentas de pedra, já que o Australopithecus garhi, datado de cerca de 2,5 milhões de anos, foi encontrado em conjunto com artefatos de ferramentas de pedra.[12] O fóssil KNM-ER 1470 (descoberto em 1972, designado Pithecanthropus rudolfensis por Alekseyev 1978) é agora visto como uma terceira espécie primitiva de Homo (ao lado de H. habilis e H. erectus) há cerca de 2 milhões de anos ou, alternativamente, como uma transição entre o Australopithecus e o Homo.[13]
Wood e Richmond (2000) propuseram que a tribo Hominini (hominídeos) de Gray abrangesse todas as espécies após o último ancestral comum chimpanzé-humano por definição, incluindo os Australopitecíneos e outras possíveis espécies pré-humanas ou para-humanas (como Ardipithecus e Sahelanthropus) não conhecidas na época de Gray.[14] Nessa sugestão, a nova subtribo de Hominina deveria ser designada como incluindo exclusivamente o gênero Homo, de modo que Hominini teria duas subtribos, Australopithecina e Hominina, com o único gênero conhecido em Hominina sendo Homo. Orrorin (2001) foi proposto como um possível ancestral de Hominina, mas não de Australopithecina.[15]
Foram propostas designações alternativas a Hominina: Australopithecinae (Gregory & Hellman 1939) e Preanthropinae (Cela-Conde & Altaba 2002).[16]
Espécie
[editar | editar código-fonte]Ao menos uma dúzia de espécies de Homo diferentes do Homo sapiens foi proposta, com diferentes graus de consenso. O Homo erectus é amplamente reconhecido como a espécie diretamente ancestral do Homo sapiens. A maioria das outras espécies propostas são apresentadas como pertencente alternativamente ao Homo erectus ou ao Homo sapiens como uma subespécie. Isso se refere particularmente ao caso do Homo ergaster.[17][18] Uma proposta divide o Homo erectus em uma variedade africana e uma asiática; a africana é o Homo ergaster, e a asiática é o Homo erectus sensu stricto. (A inclusão do Homo ergaster com o Homo erectus asiático é o Homo erectus sensu lato).[19] Parece haver uma tendência recente, com a disponibilidade de fósseis cada vez mais difíceis de classificar, como os crânios de Dmanisi (2013) ou os fósseis de Homo naledi (2015), de incluir todas as variedades arcaicas no Homo erectus.[20][21][22]
| Espécie | Período de existência (Ma) | Onde viveu | Altura (adulto) | Massa (adulto) | Capacidade encefálica (cm³) | Registro fóssil | Descoberta / publicação do nome |
|---|---|---|---|---|---|---|---|
| Hominídeo de Denisova | 0.04 | Krai de Altai (Rússia) | 1 sitio | 2010 | |||
| H. antecessor | 0.85 – 0.75 | Espanha | 1,75 m | 90 kg | 1.000 | 2 sitios | 1997 |
| H. cepranensis | 0.8 – 0.35 | Itália | 1.000 | 1 calota craniana | 1994/2003 | ||
| H. erectus | 1.8 – 0.03 | África, Eurásia (Java, China, Índia, Cáucaso) | 1.8 m | 60 kg | 850 (primeiro) – 1.100 (último) | Muitos | 1891/1892 |
| H. ergaster | 1.8 – 1.3 | África Oriental e Austral | 1,9 m | 700–850 | Muitos | 1975 | |
| H. floresiensis | 0.095 – 0.012 | Indonésia | 1,0 m | 25 kg | 400 | 7 indivíduos | 2003/2004 |
| H. gautengensis | 1.9 – 0.6 | África do Sul | 1,0 m | 1 indivíduo | 2010/2010 | ||
| H. georgicus | 1.8 | Geórgia | 600 | 4 indivíduos | 1999/2002 | ||
| H. habilis | 2.1 – 1.5 | África | 1,0 – 1,5 m | 33 – 55 kg | 510–660 | Muitos | 1960/1964 |
| H. heidelbergensis | 0.6 – 0.2 | Europa, África, China | 1,8 m | 90 kg | 1.100–1.400 | Muitos | 1908 |
| H. neanderthalensis | 0.23 – 0.028 | Europa, Sudoeste Asiático | 1,6 m | 55 – 70 kg | 1.200–1.900 | Muitos | (1829)/1864 |
| H. naledi | 0.2 - 0.3 (2017 estimativas) | África do Sul | 1,5 m | 45 kg | 560 | 1 sitio | (2013)/2015 |
| H. rhodesiensis | 0.3 – 0.125 | Zâmbia | 1.300 | Muito poucos | 1921 | ||
| H. rudolfensis | 2.5 - 1.8 | Quênia | 1 crânio | 1972/1986 | |||
| Homem da caverna do Veado Vermelho | 0.0145 - 0.0115 | China | Muito poucos | 1979/2012 | |||
| H. sapiens idaltu | 0.16 – 0.15 | Etiópia | 1.450 | 3 crânios | 1997/2003 | ||
| H. sapiens sapiens (humanos) | 0.195 – presente | Mundo | 1,5 – 1,8 m | 50 – 80 kg | 1.000–1.850 | Ainda vivo | —/1758 |
Subespécie
[editar | editar código-fonte]
Subespécies de Homo sapiens
[editar | editar código-fonte]O reconhecimento ou não reconhecimento de subespécies de Homo sapiens tem uma história complicada. A classificação de subespécie em zoologia é introduzida por conveniência, e não por critérios objetivos, com base na consideração pragmática de fatores como isolamento geográfico e seleção sexual. A classificação taxonômica informal de raça é considerada equivalente ou subordinada à classificação de subespécie, e a divisão dos humanos anatomicamente modernos (H. sapiens) em subespécies está intimamente ligada ao reconhecimento dos principais grupos raciais com base na variação genética humana.
Uma subespécie não pode ser reconhecida independentemente: uma espécie será reconhecida como não tendo nenhuma subespécie ou como tendo pelo menos duas (incluindo qualquer uma que esteja extinta). Portanto, a designação de uma subespécie existente Homo sapiens sapiens só faz sentido se pelo menos uma outra subespécie for reconhecida. O H. s. sapiens é atribuído a Linnaeus (1758) pelo princípio taxonômico da coordenação.[24] Durante o século XIX até meados do século XX, era prática comum classificar as principais divisões do H. sapiens existentes como subespécies, seguindo Linnaeus (1758), que havia reconhecido H. s. americanus, H. s. europaeus, H. s. asiaticus e H. s. afer como agrupando as populações nativas das Américas, Eurásia Ocidental, Ásia Oriental e África Subsaariana, respectivamente. Linnaeus também incluiu o H. s. ferus, para a forma "selvagem" que ele identificou com crianças selvagens, e duas outras formas "selvagens" para espécimes relatados agora considerados muito duvidosos (consulte criptozoologia), H. s. monstrosus e H. s. troglodytes.[25]
Houve variações e acréscimos às categorias de Linnaeus, como H. s. tasmanianus para a população nativa da Austrália.[26] Bory de St. Vincent, em seu Essai sur l'Homme (1825), ampliou as categorias "raciais" de Linnaeus para até quinze: Leiotrichi ("de pelo liso"): japeticus (com subraças), arabicus, iranicus, indicus, sinicus, hyperboreus, neptunianus, australasicus, columbicus, americanus, patagonicus; Oulotrichi ("de pelo crespo"): aethiopicus, cafer, hottentotus, melaninus.[27] Da mesma forma, Georges Vacher de Lapouge (1899) também tinha categorias baseadas na raça, como priscus, spelaeus, entre outras.
O Homo sapiens neanderthalensis foi proposto por King (1864) como uma alternativa ao Homo neanderthalensis.[28] Houve "guerras taxonômicas" sobre se os neandertais eram uma espécie separada desde sua descoberta na década de 1860. Pääbo (2014) enquadra isso como um debate que não pode ser resolvido em princípio, "uma vez que não há definição de espécie que descreva perfeitamente o caso".[29] Louis Lartet (1869) propôs o Homo sapiens fossilis com base nos fósseis de Cro-Magnon.
Várias propostas de variedades extintas de Homo sapiens foram feitas no século XX. Muitas das propostas originais não usavam nomenclatura trinomial explícita, embora ainda sejam citadas como sinônimos válidos de H. sapiens por Wilson e Reeder (2005):[30] Homo grimaldii (Lapouge, 1906), Homo aurignacensis hauseri (Klaatsch & Hauser, 1910), Notanthropus eurafricanus (Sergi, 1911), Homo fossilis infrasp. proto-aethiopicus (Giuffrida-Ruggeri, 1915), Telanthropus capensis (Broom, 1917),[31] Homo wadjakensis (Dubois, 1921), Homo sapiens cro-magnonensis, Homo sapiens grimaldiensis (Gregory, 1921), Homo drennani (Kleinschmidt, 1931),[32] Homo galilensis (Joleaud, 1931) = Paleanthropus palestinus (McCown & Keith, 1932).[33] Rightmire (1983) propôs o Homo sapiens rhodesiensis.[34]
Após a Segunda Guerra Mundial, a prática de dividir as populações existentes de Homo sapiens em subespécies diminuiu. Uma das primeiras autoridades que evitou explicitamente a divisão do H. sapiens em subespécies foi Grzimeks Tierleben, publicado em 1967-1972.[35] Um exemplo tardio de uma autoridade acadêmica que propôs que os grupos raciais humanos deveriam ser considerados subespécies taxonômicas foi John Baker (1974).[36] A nomenclatura trinomial Homo sapiens sapiens tornou-se popular para "humanos modernos" no contexto dos neandertais, que eram considerados uma subespécie do H. sapiens na segunda metade do século XX. Derivada da convenção, difundida na década de 1980, de considerar duas subespécies, H. s. neanderthalensis e H. s. sapiens, a alegação explícita de que "H. s. sapiens é a única subespécie humana existente" aparece no início da década de 1990.[37]
Desde os anos 2000, o extinto Homo sapiens idaltu (White et al, 2003) ganhou amplo reconhecimento como uma subespécie do Homo sapiens, mas mesmo nesse caso há uma opinião divergente que argumenta que "os crânios podem não ser suficientemente distintos para justificar um novo nome de subespécie".[38] O H. s. neanderthalensis e o H. s. rhodesiensis continuam a ser considerados espécies separadas por algumas autoridades, mas a descoberta, em 2010, de evidências genéticas de mistura de humanos arcaicos com humanos modernos reabriu os detalhes da taxonomia dos humanos arcaicos.[39]
Subespécies de Homo erectus
[editar | editar código-fonte]Desde sua introdução em 1892, o Homo erectus foi dividido em várias subespécies, muitas delas anteriormente consideradas espécies individuais de Homo. Nenhuma dessas subespécies tem consenso universal entre os paleontólogos.
- Homo erectus erectus (Homem de Java) (anos 1970)[40]
- Homo erectus yuanmouensis (Homem de Yuanmou) (Li et al., 1977)
- Homo erectus lantianensis (Homem de Lantian) (Woo Ju-Kang, 1964)
- Homo erectus nankinensis (Homem de Nanjing) (1993)
- Homo erectus pekinensis (Homem de Pequim) (1970s)[40]
- Homo erectus palaeojavanicus (Megantropo) (Tyler, 2001)
- Homo erectus soloensis (Oppenoorth, 1932)
- Homo erectus tautavelensis (Homem de Tautavel) (de Lumley e de Lumley, 1971)
- Homo erectus georgicus (1991)
- Homo erectus bilzingslebenensis (Vlček, 2002)[41]
Ver também
[editar | editar código-fonte]- Cronologia da evolução humana
- Evolução humana
- Rotas de dispersão do Homo sapiens
- Cronologia da pré-história humana
- Lista de fósseis da evolução humana
- ↑ Stringer, Chris (12 de junho de 2003). «Human evolution: Out of Ethiopia». Nature (em inglês). 423 (6941): 693–695. Bibcode:2003Natur.423..692S. PMID 12802315. doi:10.1038/423692a
- ↑ «Herto skulls (Homo sapiens idaltu)» (em inglês). talkorigins org. Consultado em 7 de junho de 2016
- ↑ Stringer, C. (2016). «The origin and evolution of Homo sapiens». Philosophical Transactions of the Royal Society B: Biological Sciences (em inglês). 371 (1698). 20150237 páginas. PMC 4920294
. PMID 27298468. doi:10.1098/rstb.2015.0237
- ↑ Jared Diamond in The Third Chimpanzee (1991), and Morris Goodman (2003) Hecht, Jeff (19 de maio de 2003). «Chimps are human, gene study implies». New Scientist (em inglês). Consultado em 8 de dezembro de 2011
- ↑ K. Wagner, Jennifer (2016). «Anthropologists' views on race, ancestry, and genetics». American Journal of Physical Anthropology (em inglês). 162 (2): 318–327. PMC 5299519
- ↑ «AAA Statement on Race». American Anthropological Association (em inglês)
- ↑ Marks, Jonathan (Maio de 2007). «Long shadow of Linnaeus's human taxonomy». Nature (em inglês). 447 (7140). 28 páginas. Bibcode:2007Natur.447Q..28M. ISSN 1476-4687. PMID 17476243. doi:10.1038/447028a
- ↑ J. E. Gray, "An outline of an attempt at the disposition of Mammalia into Tribes and Families, with a list of genera apparently appertaining to each Tribe", Annals of Philosophy, new series (1825), pp. 337–344.
- ↑ Cela-Conde, C. J.; Ayala, F. J. (2003). «Genera of the human lineage». Proceedings of the National Academy of Sciences (em inglês). 100 (13): 7684–7689. Bibcode:2003PNAS..100.7684C. PMC 164648
- ↑ Coryndon Memorial Museum.; Museum, Coryndon Memorial; Society, East Africa Natural History; Kenya, Museums Trustees of (1944). Journal of the East Africa Natural History Society (em inglês). 18. Nairobi, Kenya: The Museum
- ↑ Villmoare, B. (2015). «Early Homo at 2.8 Ma from Ledi-Geraru, Afar, Ethiopia». Science (em inglês). 347 (6228): 1352–1355. Bibcode:2015Sci...347.1352V. PMID 25739410. doi:10.1126/science.aaa1343
- ↑ De Heinzelin, J; Clark, JD; White, T; Hart, W; Renne, P; Woldegabriel, G; Beyene, Y; Vrba, E (1999). «Environment and behavior of 2.5-million-year-old Bouri hominids». Science (em inglês). 284 (5414): 625–9. Bibcode:1999Sci...284..625D. PMID 10213682. doi:10.1126/science.284.5414.625
- ↑ Kaplan, Matt (8 de agosto de 2012). «Fossils point to a big family for human ancestors». Nature (em inglês). doi:10.1038/nature.2012.11144. Consultado em 8 de agosto de 2012
- ↑ Wood, Bernard; Richmond, Brian G. (2000). «Human evolution: taxonomy and paleobiology». Journal of Anatomy (em inglês). 197 (Pt 1): 19–60. PMC 1468107
- ↑ Reynolds, Sally C; Gallagher, Andrew (29 de março de 2012). African Genesis: Perspectives on Hominin Evolution (em inglês). [S.l.: s.n.] ISBN 9781107019959
- ↑ Brunet, M.; et al. (2002). «A new hominid from the upper Miocene of Chad, central Africa» (PDF). Nature. 418 (6894): 145–151. Bibcode:2002Natur.418..145B. PMID 12110880. doi:10.1038/nature00879 Cela-Conde, C.J.; Ayala, F.J. (2003). «Genera of the human lineage». PNAS. 100 (13): 7684–7689. Bibcode:2003PNAS..100.7684C. PMC 164648
- ↑ Hazarika, Manji (Junho de 2007). «Homo erectus / ergaster and Out of Africa: Recent Developments in Paleoanthropology and Prehistoric Archaeology» (PDF) (em inglês)
- ↑ Klein, R. (1999). The Human Career: Human biological and cultural origins (em inglês). Chicago, IL: University of Chicago Press. ISBN 0226439631
- ↑ Antón, S.C. (2003). «Natural history of Homo erectus». American Journal of Physical Anthropology (em inglês). 122: 126–170. PMID 14666536. doi:10.1002/ajpa.10399.
Na década de 1980, o número crescente de espécimes de "H. erectus", especialmente na África, levou à constatação de que o "H. erectus" asiático ("H. erectus sensu stricto"), antes considerado tão primitivo, era, na verdade, mais derivado do que seus equivalentes africanos. Essas diferenças morfológicas foram interpretadas por alguns como evidência de que mais de uma espécie poderia ser incluída no H. erectus sensu lato (por exemplo, Stringer, 1984; Andrews, 1984; Tattersall, 1986; Wood, 1984, 1991a, b; Schwartz e Tattersall, 2000) ... Ao contrário da linhagem europeia, na minha opinião, as questões taxonômicas que envolvem o H. erectus asiático versus o africano são mais intratáveis. A questão foi abordada de forma mais incisiva com a denominação de H. ergaster com base na mandíbula tipo KNM-ER 992, mas também incluindo o esqueleto parcial e os dentes isolados do KNM-ER 803 entre outros restos mortais de Koobi Fora (Groves e Mazak, 1975). Recentemente, esse nome específico foi aplicado à maioria dos primeiros H. erectus da África e da Geórgia, em reconhecimento à natureza menos derivada desses restos mortais em relação às condições do H. erectus asiático (ver Wood, 1991a, p. 268; Gabunia et al., 2000a). Pelo menos partes do parátipo do H. ergaster (por exemplo, KNM-ER 1805) não estão incluídas na maioria das concepções atuais desse táxon. A questão do "H. ergaster" permanece notoriamente não resolvida (por exemplo, Stringer, 1984; Tattersall, 1986; Wood, 1991a, 1994; Rightmire, 1998b; Gabunia et al., 2000a; Schwartz e Tattersall, 2000), em grande parte porque o diagnóstico original não forneceu nenhuma comparação com o registro fóssil asiático.
- ↑ «Skull suggests three early human species were one». News & Comment (em inglês). Nature
- ↑ Lordkipanidze, David; Ponce de Leòn, Marcia S.; Margvelashvili, Ann; Rak, Yoel; Rightmire, G. Philip; Vekua, Abesalom; Zollikofer, Christoph P. E. (18 de outubro de 2013). «A Complete Skull from Dmanisi, Georgia, and the Evolutionary Biology of Early Homo». Science (em inglês). 342 (6156): 326–331. Bibcode:2013Sci...342..326L. PMID 24136960. doi:10.1126/science.1238484
- ↑ Switek, Brian (17 de outubro de 2013). «Beautiful skull spurs debate on human history» (em inglês). National Geographic. Consultado em 22 de setembro de 2014
- ↑ Ralph, Bob (19 de fevereiro de 1987). «Conforming to type». New Scientist (em inglês) (1548). p. 59.
as far as I know, there is no type material for Homo sapiens. To be fair to Linnaeus, the practice of setting type specimens aside doesn't seem to have developed until a century or so later.
- ↑ «article 46.1». ICZN glossary (em inglês) 4th ed. [S.l.]: International Code of Zoological Nomenclature.
Declaração do Princípio de Coordenação aplicado a nomes de grupos de espécies. Considera-se que um nome estabelecido para um táxon em qualquer uma das classificações do grupo de espécies foi estabelecido simultaneamente pelo mesmo autor para um táxon na outra classificação do grupo; ambos os táxons nominais têm o mesmo tipo de nome, quer esse tipo tenha sido fixado originalmente ou posteriormente.
O Homo sapiens sapiens é raramente usado antes da década de 1940. Em 1946, John Wendell Bailey atribui o nome a Linnaeus (1758) de forma explícita: "Linnaeus. Syst. Nat. ed. 10, Vol. 1. pp. 20, 21, 22, lista cinco raças de homem, viz: ''Homo sapiens sapiens'' (branco - caucasiano) [...]". Essa é uma atribuição errônea, mas o ''H. s. sapiens'' tem sido frequentemente atribuído a Linnaeus desde então. De fato, Linnaeus, Syst. Nat. ed. 10 Vol. 1. p. 21 não tem ''Homo sapiens sapiens'', a raça "branca" ou "caucasiana" sendo chamada de ''Homo sapiens Europaeus''. Isso é explicitamente apontado no ''Bulletin der Schweizerische Gesellschaft für Anthropologie und Ethnologie'' Volume 21 (1944), p. 18 (argumentando não contra o ''H. s. sapiens'', mas contra o ''H. s. albus L'' proposto por von Eickstedt e Peters): "die europide Rassengruppe, als Subspecies aufgefasst, [würde] Homo sapiens eurpoaeus L. heissen" ("o grupo racial europeu, considerado como uma subespécie, seria chamado de ''H. s. europeaeus L."). Veja também: John R. Baker, Race, Oxford University Press (1974), 205. - ↑ Linné, Carl von; Gesellschaft, Deutsche Zoologische. Caroli Linnæi Systema naturæ (em sueco) Editio decima, 1758, / cura Societatis Zoologicae Germanicae iterum edita ed. Lipsiæ: Sumptibus Guilielmi Engelmann
- ↑ Fage, John Donnelly; Clark, John Desmond (1975). The Cambridge History of Africa (em inglês). [S.l.]: Cambridge University Press
- ↑ Annals of Philosophy (em inglês). [S.l.]: Baldwin, Cradock, and Joy. 1826
- ↑ Szalay, Frederick S.; Delson, Eric (22 de outubro de 2013). Evolutionary History of the Primates (em inglês). [S.l.]: Academic Press
- ↑ Pääbo, Svante (2014). Neanderthal Man: In Search of Lost Genomes (em inglês). New York: Basic Books. p. 237
- ↑ Groves, C.P. (2005). Wilson, D. E.; Reeder, D. M, eds. Mammal Species of the World 3.ª ed. Baltimore: Johns Hopkins University Press. ISBN 978-0-8018-8221-0. OCLC 62265494
- ↑ Harrison, T. (2013). Kimbel, William H.; Martin, Lawrence B., eds. Species, Species Concepts and Primate Evolution (em inglês). [S.l.: s.n.] p. 361. ISBN 9781489937452
- ↑ M. R. Drennan, "An Australoid Skull from the Cape Flats", The Journal of the Royal Anthropological Institute of Great Britain and Ireland Vol. 59 (Jul. - Dec., 1929), 417-427.
- ↑ among other names suggested for fossils later subsumed under neanderthalensis, see: Delson, Eric; Tattersall, Ian; Couvering, John Van; Brooks, Alison S. (2004). Encyclopedia of Human Evolution and Prehistory Second ed. [S.l.]: Routledge. ISBN 9781135582272
- ↑ Rightmire, GP (3 de junho de 1983). «The Lake Ndutu cranium and early Homo sapiens in Africa». American Journal of Physical Anthropology (em inglês). 61 (2): 245–54. PMID 6410925. doi:10.1002/ajpa.1330610214
- ↑ English translation (1972–1975): Grzimek's Animal Life Encyclopedia, Volume 11, p. 55.
- ↑ John R. Baker, Race, Oxford University Press (1974).
- ↑ Kitahara, Michio (1991). The tragedy of evolution: the human animal confronts modern society (em inglês). [S.l.: s.n.] p. xi.
We are the only surviving subspecies of Homo sapiens
- ↑ Stringer, Chris (12 de junho de 2003). «Human evolution: Out of Ethiopia». Nature (em inglês). 423 (6941): 692–3, 695. Bibcode:2003Natur.423..692S. PMID 12802315. doi:10.1038/423692a
- ↑ Hublin, J. J. (2009). «The origin of Neandertals». Proceedings of the National Academy of Sciences (em inglês). 106 (38): 16022–7. Bibcode:2009PNAS..10616022H. JSTOR 40485013. PMC 2752594
- ↑ a b In the 1970s a tendency developed to regard the Javanese variety of H. erectus as a subspecies, Homo erectus erectus, with the Chinese variety being referred to as Homo erectus pekinensis. See: Sartono, S. (12 de maio de 2011). «Implications arising from Pithecanthropus VIII». In: Tuttle, Russell H. Paleoanthropology: Morphology and Paleoecology (em inglês). [S.l.: s.n.] p. 328. ISBN 9783110810691
- ↑ Emanuel Vlček: Der fossile Mensch von Bilzingsleben (= Bilzingsleben. Bd. 6 = Beiträge zur Ur- und Frühgeschichte Mitteleuropas 35). Beier & Beran, Langenweißbach 2002.