eIF2
eIF2 – eukariotyczny czynnik inicjacji translacji 2 (ang. eukaryotic initiation factor 2) – białko, które bierze udział w inicjacji translacji (biosyntezy białek w komórkach). Wchodzi w skład tzw. kompleksu inicjacyjnego. Uczestniczy w wiązaniu tRNAiMet (inicjatorowego tRNA transportującego metioninę) z małą podjednostką rybosomu 40S przy udziale GTP. Składa się z trzech podjednostek: α, β i γ.
Kiedy proces inicjacji translacji jest zakończony, eIF2 odłącza się od rybosomu w formie nieaktywnego kompleksu związanego z GDP. Żeby był znów aktywny, GDP musi zostać zamienione na GTP – w takiej formie eIF2 jest gotowe do udziału w kolejnym procesie inicjacji translacji.
Funkcja
[edytuj | edytuj kod]
eIF2 jest czynnikiem niezbędnym do syntezy białek. Tworzy kompleks inicjacyjny (TC) z GTP i tRNAiMet, który następnie wiąże się z podjednostką rybosomu 40S, tworząc tzw. kompleks preinicjacyjny 43S (PIC). Jak wynika z eksperymentów in vitro, czynniki inicjacyjne eIF1, eIF2A i eIF3 stymulują powstanie kompleksu inicjacyjnego. Kompleks PIC wiąże mRNA, do którego związał się czynnik eIF4E, w wyniku czego powstaje kompleks 48S, który rozpoczyna poszukiwanie w mRNA kodonu startu translacji AUG (adenina – uracyl – guanina). Po związaniu kodonu AUG z antykodonem inicjatorowego tRNAiMet czynnik eIF5 (białko aktywujące GTPazę) przyłącza się do kompleksu inicjacyjnego i indukuje hydrolizę GTP przez eIF2. Wówczas czynniki inicjacyjne, a wśród nich eIF2, teraz związane z GDP (guanozynodwufosforanem), są uwalniane z kompleksu 48S, a następnie przyłącza się do niego podjednostka rybosomu 60S i powstaje kompleks inicjacyjny 80S. Wtedy rozpoczyna się etap elongacji[1].
Uwolnione eIF2 związane z GDP jest nieaktywne. GDP jest wymieniane na GTP z udziałem białka eIF2B (czynnika wymiany nukleotydów guaninowych, ang. Guanine nucleotide exchange factor). Po przyłączeniu tRNAiMet eIF2 w kompleksie potrójnym jest znów gotowe do udziału w kolejnej inicjacji translacji[2][3][4].
Struktura
[edytuj | edytuj kod]eIF2 jest heterotrimerem o masie 126 kDa. Składa się z trzech podjednostek – α, Β i γ – o silnie konserwowanych sekwencjach (sekwencja aminokwasów identyczna w kilkudziesięciu procentach (od 47% do 72%) dla każdej podjednostki eIF2 z Homo sapiens i Saccharomyces cerevisiae).
| Podjednostka | α | β | γ |
|---|---|---|---|
| Masa cząsteczkowa [kDa] | 36 | 38 | 52 |
| Podobieństwo | Rodzina białek eIF2 | Rodzina czynników elongacji wiążących GTP | Rodzina białek eIF2β / eIF5 |
| Interakcje | Wiąże się z eIF5, eIF2B i RNA | Wiąże się z GTP i z RNA |
Podjednostka α zawiera serynę w pozycji 51, która jest fosforylowana, oraz domenę S1, która prawdopodobnie wiąże RNA. Podjednostka α jest uważana za podjednostkę regulatorową trimeru.
Podjednostka β może być fosforylowana w kilku różnych miejscach (aminokwasy 2, 13, 67 i 218). W jej domenie N-końcowej znajdują się trzy fragmenty polilizyny ważne dla oddziaływań eIF2 z eIF2B. Sekwencja podjednostki β zawiera też motyw palca cynkowego, który uczestniczy zarówno w tworzeniu kompleksu potrójnego, jak i w formowaniu kompleksu preinicjacyjnego 43S. Wykryto też dwie sekwencje wiążące guaninę. Ich związek z regulacją aktywności eIF2 nie jest udowodniony. Podjednostka β prawdopodobnie oddziałuje zarówno z tRNA, jak i z mRNA.
W podjednostce γ znajdują się trzy miejsca wiążące guaninę. To głównie ona jest odpowiedzialna za wiązanie z GTP / GDP. Zawiera też szczelinę wiążącą tRNA, co wynika z danych krystalograficznych. Motyw kłykcia cynkowego (ang. zinc knuckle) może związać jeden jon Zn2+[4][6][7].
Regulacja
[edytuj | edytuj kod]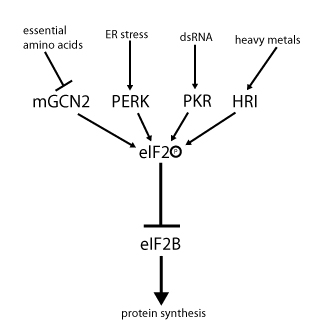
Aktywność eIF2 jest regulowana przez fosforylację oraz przez wymianę nukleotydu guaninowego GDP na GTP.
Podjednostka α jest fosforylowana w pozycji seryny 51 przez różne kinazy serynowe, które uaktywniają się w sytuacjach stresu komórkowego spowodowanego obecnością niezwiniętych lub źle zwiniętych białek, czyli na przykład w przypadku niedostatku aminokwasów (kinaza GCN2), stresu siateczki śródplazmatycznej (PERK), obecności dsRNA (PKR) lub braku hemu (HR). Ufosforylowane eIF2 wykazuje zwiększone powinowactwo do czynnika eIF2B, który wymienia GDP na GTP. Ale eIF2B wymienia GDP na GTP tylko w nieufosforylowanym czynniku eIF2. Wobec tego ufosforylowany eIF2 działa jak inhibitor własnego czynnika wymiany nukleotydów eIF2B. Ponieważ stężenie eIF2B w komórce jest dużo niższe niż stężenie eIF2, to nawet niewielka ilość ufosforylowanego eIF2 może dezaktywować wszystkie cząsteczki eIF2B. Wskutek tego przy braku eIF2B cząsteczki eIF2 pozostają nieaktywne, bo wciąż są związane z GDP, które nie może zostać wymienione na GTP. W rezultacie translacja zostaje zatrzymana z powodu braku dostępnych kompleksów eIF2-GTP-tRNAiMet[2][3][4][8].
Choroby związanie z niewłaściwym funkcjonowaniem eIF2
[edytuj | edytuj kod]Białko eIF2 jest niezbędne w procesie inicjacji translacji zależnej od kapu, defekty eIF2 są zatem letalne. eIF2 jest białkiem wysoce konserwowanym ewolucyjnie; jego cząsteczki są podobne, nawet jeśli pochodzą z odległych ewolucyjnie gatunków. Z tego wynika, że mutacje eIF2 mają duży wpływ na przeżywalność komórek.
Nie znamy więc chorób wywołanych bezpośrednio przez mutacje eIF2. Istnieje jednak wiele schorzeń związanych z niewłaściwą regulacją aktywności eIF2 przez kinazy, np.:
- Zwiększoną koncentrację aktywnej kinazy PKR i nieaktywnej (ufosforylowanej) formy eIF2 wykryto u pacjentów cierpiących na choroby neurodegeneracyjne, np. chorobę Alzheimera, Parkinsona i Huntingtona.
- Mutacje wszystkich pięciu podjednostek eIF2B (EIF2B1, EIF2B2, EIF2B3, EIF2B4 i EIF2B5) mogą być związane z leukoencefalopatią – chorobą, która wywołuje zanik substancji białej w mózgu. Przyczyny, dla których tylko komórki mózgu wydają się być dotknięte chorobą, nie są jeszcze w pełni zrozumiane.
Niestabilny poziom białek regulatorowych funkcji eIF2 może mieć znaczenie dla rozwoju wymienionych chorób[4][9].
Zobacz też
[edytuj | edytuj kod]Przypisy
[edytuj | edytuj kod]- ↑ Bruce Alberts i inni, Podstawy biologii komórki, PWN.
- ↑ a b S.R. Kimball, Eukaryotic initiation factor eIF2, „The International Journal of Biochemistry & Cell Biology”, 31 (1), 1999, s. 25–29, DOI: 10.1016/s1357-2725(98)00128-9, PMID: 10216940 (ang.).
- ↑ a b c J.W. Hershey, Protein phosphorylation controls translation rates, „Journal of Biological Chemistry”, 264 (35), 1989, s. 20823–20826, PMID: 2687263 (ang.).
- ↑ a b c d Alan G. Hinnebusch, Translational regulation of GCN4 and the general amino acid control of yeast, „Annual Review of Microbiology”, 59, 2005, s. 407–450, DOI: 10.1146/annurev.micro.59.031805.133833, PMID: 16153175 (ang.).
- ↑ Scot R. Kimball, Leonard S. Jefferson, Amino acids as regulators of gene expression, „Nutrition & Metabolism”, 1 (1), 2004, s. 3, DOI: 10.1186/1743-7075-1-3, PMID: 15507151, PMCID: PMC524028 (ang.).
- ↑ Antonina Roll-Mecak i inni, X-ray structure of translation initiation factor eIF2gamma: implications for tRNA and eIF2alpha binding, „Journal of Biological Chemistry”, 279 (11), 2004, s. 10634–10642, DOI: 10.1074/jbc.M310418200, PMID: 14688270 (ang.).
- ↑ Takuhiro Ito, Assen Marintchev, Gerhard Wagner, Solution structure of human initiation factor eIF2alpha reveals homology to the elongation factor eEF1B, „Structure”, 12 (9), 2004, s. 1693–1704, DOI: 10.1016/j.str.2004.07.010, PMID: 15341733 (ang.).
- ↑ a b J. Nika, S. Rippel, E.M. Hannig, Biochemical analysis of the eIF2beta gamma complex reveals a structural function for eIF2alpha in catalyzed nucleotide exchange, „Journal of Biological Chemistry”, 276 (2), 2001, s. 1051–1056, DOI: 10.1074/jbc.M007398200, PMID: 11042214 (ang.).
- ↑ Raymond Chuen-Chung Chang, Man-Shan Yu, Cora Sau-Wan Lai, Significance of molecular signaling for protein translation control in neurodegenerative diseases, „Neuro-Signals”, 15 (5), 2007, s. 249–258, DOI: 10.1159/000102599, PMID: 17496426 (ang.).