Trilobieten
| Trilobieten Status: Uitgestorven, als fossiel bekend Fossiel voorkomen: Cambrium - Perm | |||||||
|---|---|---|---|---|---|---|---|

| |||||||
| Paradoxides, 163 mm, Midden-Cambrium, Marokko | |||||||
| Taxonomische indeling | |||||||
| |||||||
| Klasse | |||||||
| Trilobita Walch, 1771 | |||||||

| |||||||
| Koneprusia brutoni | |||||||
| Afbeeldingen op | |||||||
| Trilobieten op | |||||||
| |||||||
De Trilobieten (Trilobita, vertaald drielobbigen) vormen een bekende klasse uitgestorven geleedpotigen, die in zee leefden. Trilobieten behoren met ammonieten en dinosauriërs ongetwijfeld tot de bekendste fossielen. Trilobieten zijn bekend van 521 tot 250, ammonieten van 400 tot 66 en dinosauriërs van 231 tot 66 miljoen jaar geleden.
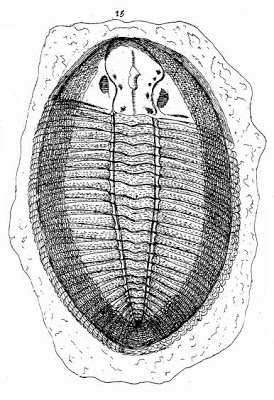
Trilobieten zijn te herkennen aan een verkalkt exoskelet aan de rugkant van het lichaam, dat aan de rand van de buikzijde is teruggeslagen en verder aan de buikzijde is er een eveneens verkalkt gehemelte. Het lichaam is van voor naar achter opgebouwd uit een aantal segmenten. De voorste daarvan zijn samengegroeid tot het kopschild (of cephalon) en de achterste tot het staartschild (of pygidium). De segmenten daar tussenin vormen samen het lijf (of thorax) en scharnieren afzonderlijk ten opzichte van elkaar, en het kop- en staartschild. Trilobieten konden zich in de lengterichting ter verdediging oprollen zoals een pissebed of een gordeldier. Elk van die drie lichaamsdelen heeft in de lengterichting een as, die vaak met een groef is gescheiden van de wangen en ribben aan weerszijden, en naar deze drie lobben verwijst de naam trilobiet. Op de wangen staat meestal een paar ogen die zijn samengesteld uit facetlenzen gemaakt van calciet.[1]
Zelden worden ook niet-verkalkende lichaamsdelen van trilobieten gevonden, die duidelijk maken dat vermoedelijk het tweede segment van voren aan de buikkant een paar onvertakte antennes draagt die bestaan uit tientallen als een kralensnoer geregen elementen. Alle volgende segmenten dragen aan de buikkant gelijkvormige paren twee-armige poten, waarvan de binnenste looppoot of endopodiet bestaat uit 6 of 7 elementen en de buitenste kieuwpoot of exopodiet een grote hoeveelheid parallelle lamellen draagt. Verder worden resten gevonden waarvan kan worden afgeleid dat de darm vanaf het gehemelte middendoor loopt tot het puntje van het staartschild. Bij één soort zijn twee onvertakte antenneachtige uitsteeksels (of cerci) achter aan het lichaam gevonden.[1]
De eerste keer dat trilobieten in het sediment verschijnen definieert de basis van het Atdabanien, ongeveer halverwege het Onder-Cambrium (521 miljoen jaar geleden). Ze waren algemeen en kregen een steeds grotere verscheidenheid tot het eind van het Ordovicium, toen een aantal orden en levenswijzen verdween. Die types zijn daarna niet opnieuw ontstaan. Gedurende het laatste deel van het Devoon zijn alle nog overgebleven ordes behalve de Proetida uitgestorven. Deze laatste groep trilobieten verdween tijdens de massa-extinctie aan het einde van het Perm, ongeveer 250 miljoen jaar geleden. De trilobieten waren een van de meest succesvolle vroege groepen dieren, en bewoonden de oceanen meer dan 270 miljoen jaar lang.[1]
Toen de fossielen van trilobieten verschenen, hadden ze al verschillende vormen en kwamen ze al snel voor langs de randen van alle continenten. Omdat trilobieten aan de rugkant een exoskelet hebben dat bestaat uit kalk, zijn er veel bewaard gebleven. In totaal zijn ongeveer 17.000 soorten in 9 ordes beschreven, verspreid over het Paleozoïcum. De studie van deze fossielen heeft belangrijke bijdrages geleverd aan de biostratigrafie, paleontologie, evolutiebiologie en het begrip van de platentektoniek. Over de plaatsing van de trilobieten binnen de geleedpotigen wordt door wetenschappers nog verschillend gedacht, maar recent onderzoek suggereert verwantschap met de Chelicerata.[2]
Trilobieten specialiseerden zich in verschillende levenswijzen. Veel soorten bewogen over de zeebodem als roofdier, aaseter of voedselfilteraar en sommige zwommen in het water en leefden van plankton. De meeste levensstijlen die je tegenkomt bij moderne mariene geleedpotigen kwamen ook voor bij trilobieten, mogelijke met uitzondering van parasitisme (waar nog wetenschappelijk debat over is).[3] Van enkele trilobieten, met name uit de familie Olenidae, denken sommige wetenschappers dat ze een symbiotische relatie onderhielden met anaerobe bacteriën die organisch materiaal oxideerden met zuurstof uit sulfaat,[4] maar deze interpretatie is omstreden.
Oorsprong en evolutie
[bewerken | brontekst bewerken]Ook de vroegste trilobieten hebben bijna alle gemeenschappelijke kenmerken van hun afstammelingen. Alleen van de onderorde Olenellina zijn de vroegste larvale stadia (phaselus en protaspide) nooit gevonden, naar wordt aangenomen omdat in deze stadia bij deze onderorde het exoskelet niet verkalkt was. Ook ontbreken de gezichtsscheurnaden aan de rugkant van het kopschild bij deze groep. Er wordt van uitgegaan dat deze unieke eigenschappen de oorspronkelijke toestand vertegenwoordigen, en dat alle andere trilobieten dus van de Olenellina afstammen. Ongeveer 521 miljoen jaar geleden (aan het begin van de 3de etage van het Cambrium) verschijnen er bijna gelijktijdig op verschillende plaatsen op de wereld plotseling een aantal soorten trilobiet die tot verschillende families en zelfs ordes worden gerekend, zoals Profallotaspis jakutensis uit Siberië en Fritzaspis sp. uit het westen van de VS, beide uit de familie Fallotaspidae, onderorde Olenellina; Lemdadella linaresae, familie Redlichiidae, onderorde Redlichiina; en Hupetina antiqua (Marokko) en Serrania gordaensis (Spanje), beide familie Bigotinidae, orde Ptychopariida.[5]
Trilobieten zijn waarschijnlijk ontstaan in het huidige Siberië, om zich van daaruit te verspreiden over de randzeeën van alle toenmalige continenten.[6] Het is dus wel zeker dat de recentste gemeenschappelijke voorouder van alle trilobieten meer dan 521 miljoen jaar geleden leefde, maar de fossielen daarvan zijn nog niet gevonden. Er zijn tot dusver geen fossiele overgangsvormen gevonden die een combinatie hebben van de eigenschappen van trilobieten met eigenschappen van andere groepen vroege geleedpotigen.[7] De morfologische overeenkomsten tussen trilobieten en andere vroege en mogelijk geleedpotige wezens die leefden tijdens het Ediacarium, zoals Spriggina en Parvancorina, zijn te dubbelzinnig om gefundeerde uitspraken te doen over de voorouders van trilobieten.
Evolutionaire trends
[bewerken | brontekst bewerken]In veel grote families uit het Cambrium varieert het aantal scharnierende segmenten van het middenlijf (of thorax) sterk (Redlichiidae 11-17, Paradoxididae 13-22, Oryctocephalidae 5-18, Ptychopariidae 12-17, Conocoryphidae 7-25, Solenopleuridae 11-17, Olenidae 9-19). Vanaf het Boven-Cambrium wordt de spreiding daarin echter veel kleiner in vrijwel alle grote groepen trilobieten (Cyclopygidae 5-6, Calymenidae 12-13, Illaenidae, Proetidae en Odontopleuridae 8-10) of vast (Scutellidae 10, Asaphidae 8, Trinucleidae 6, Phacopina 11). Deze evolutionaire tendens naar minder variabiliteit in het aantal thoraxsegmenten is een illustratie van de Wet van Rosa (in het Engels Rosa's rule, 1899).[8] Overigens hebben de soorten van de orde Agnostida die in het Onder-Cambrium ontstond een heel geringe variatie met slechts 2 of 3 thoraxsegmenten, en 2 segmenten bij de Agnostina, de onderorde die het Midden-Cambrium overleefde.
Het verkalkte rugschild van trilobieten is, in combinatie met het oprollen van het lichaam, waarschijnlijk een bescherming tegen roofdieren en verklaart mogelijk deels het langdurige succes van deze diergroep. We kunnen veronderstellen dat roofdieren zich aanpasten om de verdediging van de trilobieten te omzeilen en in reactie daarop is in de loop van de tijd het oprollen van trilobieten steeds perfecter geworden. Bij de vroegste trilobieten is het staartschild meestal klein, maar bij latere trilobieten wordt het staartschild in omtrek steeds meer een kopie van het kopschild.[9]
Gedurende het Cambrium en Ordovicium zijn er veel voorbeelden te vinden van miniaturisatie en reductie van het aantal thorax segmenten en vaak verlies van de ogen. Voorbeelden daarvan zijn alle Agnostida, maar ook Thoracocare, Shumardia, Schmalenseeia en Acanthopleurella.[10] Ctenopyge soorten zijn zeer klein en hebben lange stekels in het platte vlak, een aanpassing aan het leven als plankton. Ze hebben ook heel kleine maar functionele ogen, wat suggereert dat de soort leefde in de bovenste waterlaag.[11] Na het Ordovicium komen deze miniatuursoorten niet meer voor.
Uitsterven
[bewerken | brontekst bewerken]
Verspreid over het Paleozoïcum zijn er een aantal kleinere en grotere massa-extincties geweest waarbij ook veel trilobieten zijn uitgestorven. Al aan het eind van het Onder-Cambrium sterven belangrijke groepen trilobieten uit, zoals alle leden van de onderorde Olenellina, maar na deze vroege gebeurtenissen neemt de diversiteit weer snel toe. Deze massa-extincties hebben diverse oorzaken, zoals grootschalig vulkanisme, het neerstorten van een grote meteoriet en het samenkomen van alle continenten in een supercontinent. Die gebeurtenissen hebben bijvoorbeeld geleid tot plotselinge klimaatverandering en zuurstofarmoede in het water. Mogelijk heeft ook het op gang komen van de plantengroei op het land en het daardoor in zee spoelen van grote hoeveelheden humuszuur een rol gespeeld. De diversiteit onder de trilobieten bereikte een hoogtepunt tijdens de overgang van het Cambrium naar het Ordovicium. Aan het eind van het Ordovicium, en gedurende het Devoon nam de variatie in soorten steeds verder af. De enige orde waarvan vertegenwoordigers het eind van het Devoon overleefden waren de Proetida. Tijdens de Perm-Trias-massa-extinctie stierf ook deze laatste groep trilobieten uit.[10]
Lichaamsbouw
[bewerken | brontekst bewerken]


Van trilobieten wordt over het algemeen alleen (meestal een deel van) het verkalkte exoskelet gevonden. Alleen op een handvol locaties (Konservat-Lagerstätten) zijn herkenbare niet-verkalkte lichaamsdelen gevonden zoals antennes, poten, kieuwen, spieren en het spijsverteringskanaal. In de wetenschappelijke literatuur worden zulke delen vaak aangeduid als een week lichaam of soft tissue, hoewel de uitwendige delen een buitenkant van chitine hadden.
Trilobieten variëren in lengte van minuscuul (minder dan 3 mm) tot zeer groot (meer dan 30 cm), met een gemiddelde grootte van 3-10 cm. Een van de kleinste soorten is Acanthopleurella met een maximum van 1½ mm.[14] 's Werelds grootst bekende trilobiet is een exemplaar van Isotelus rex van 72 cm, dat in 1998 werd gevonden door Canadese wetenschappers in gesteente uit het Ordovicium aan de oevers van de Hudsonbaai.[15]
Het exoskelet is samengesteld uit calciet en calciumfosfaat ingebed in een eiwitraster van chitine dat het oppervlak aan de bovenkant/rugkant (of dorsaal) van de trilobiet bedekt en omslaat naar de onderkant/buikkant (of ventraal) en daar een zoom vormt, de zogenoemde doublure. Er zijn drie hoofdonderdelen (of tagmata): het kopschild (of cephalon), het lijf (of thorax) en het staartschild (of pygidium). Tijdens het vervellen, scheurde het exoskelet over het algemeen tussen de kop en het lijf, en dat is de oorzaak dat bij zo veel trilobietfossielen de een of de ander ontbreekt. Ook de scheurnaden op het kopschild hebben geholpen de vervelling te vergemakkelijken (behalve bij de Olenellina, de Agnostina, de Phacopidae en een aantal oogloze soorten waar deze scheurnaden ontbreken of niet functioneel zijn). Net als bij kreeften pompten trilobieten waarschijnlijk hun lijf op als het nieuwe exoskelet nog zacht was, zodat ze tussen de vervellingen ruimte hadden voor groei van hun interne massa.[1]
Kopschild
[bewerken | brontekst bewerken]

Het kopschild van een trilobiet (of cephalon) is zeer variabel en complex. De centrale as wordt in de kop glabella genoemd. Daarnaast en daarvoor bevinden zich de zogenoemde wangen (of met een wetenschappelijke term genae). De glabella is van de wangen afgescheiden door een groef, of in elk geval een plotselinge verandering van de helling. De glabella heeft altijd een koepelvorm, waarschijnlijk om daaronder ruimte te hebben voor de mondholte of maag. In de glabella zijn vaak nog maximaal 5 segmenten te herkennen, van elkaar afgegrensd door dwarsgroeven. Aan de binnenkant/onderkant van de glabella zitten vaak deuken die worden gezien als de plaatsen waar spieren waren vastgehecht. De rest van het kopschild wordt aangeduid als wangen, en daarin zijn geen overblijfselen van segmenten, met uitzondering van het achterste dat vaak een voortzetting is van de occipitale ring in de glabella in de vorm van een zoom, die overigens een geheel vormt met een zoom langs de zij- en voorrand van het kopschild. De wangen bestaan uit de vaste wangen (of fixigenae) tegen de glabella aan, en vrije wangen (of librigenae), die bij het vervellen van de kop afbreken op een zwakke scheurnaad. De rand van het kopschild heeft meestal een zoom (in het Engels border), die van de rest van de wangen is afgescheiden door een groef, of in elk geval een plotselinge verandering van de helling. Deze loopt vaak over in het achterste segment in de wang en in de glabella, waar dit segment occipitale ring heet. Aan de voor- en zijrand van het kopschild is deze zoom omgeslagen naar de buikzijde (de zogenoemde doublure). Eveneens aan de buikzijde en ongeveer tegenover de voorste segmenten van de glabella bevindt zich het gehemelte (of hypostoom), een verkalkte plaat die vaak aan de voorkant met een scheurnaad verbonden is met de zoom, maar soms ook geen contact had met de rest van het verkalkte kopschild. De mondopening was waarschijnlijk naar achter gericht en zat op de achterste rand van de hypostoom. De segmenten in het kopschild, de afgrenzing tussen glabella en wangen en van de zoom zijn soms uitgevlakt, zodat ze niet goed meer kunnen worden onderscheiden. Dit verschijnsel (effacement) heeft zich vele malen onafhankelijk voorgedaan bij trilobieten uit allerlei tijdvakken en taxonomische groepen.[1]
Scheurnaden
[bewerken | brontekst bewerken]Op het kopschild van trilobieten komen verschillende lijnvormige zwaktes voor die openbreken of scheuren als het dier vervelde. In dit artikel worden die aangeduid met de term scheurnaden (in het Engels sutures). De aanwezigheid en het verloop van deze scheurnaden kan sterk verschillen tussen de verschillende groepen trilobieten en zijn behulpzaam bij de identificatie ervan. De volgende scheurnaden komen voor in het kopschild van trilobieten:
- lipscheurnaden (rostral sutures) scheiden aan de buikkant de doublure aan de zijkant van het kopschild van de voorkant ervan (de lip, in het Engels rostral plate).
- gehemeltescheurnaad (hypostomal suture) scheidt aan de buikkant het gehemelte van de lip.
- randscheurnaden (marginal sutures) lopen over de buitenrand van het kopschild tussen de zoom (border) aan de rugkant en die aan de buikkant (doublure).
- oogscheurnaden (ocular sutures) lopen aan de rugkant rondom elk van beide ogen, en zorgen er mogelijk voor dat de ogen niet gelijktijdig met de rest van het exoskelet vervangen hoeven te worden. Oogscheurnaden komen veel voor bij trilobieten uit het Cambrium.
- gezichtsscheurnaden (facial sutures) scheiden aan de rugkant de vaste wangen van de vrije wangen.[1]
Gezichtsscheurnaden
[bewerken | brontekst bewerken]
Gezichtsscheurnaden kunnen worden onderscheiden in vijf types, afhankelijk van waar de achterkant ervan de rand van het kopschild snijdt, en of ze oorspronkelijk afwezig zijn of naderhand verloren zijn gegaan.[16]
- afwezig: gezichtsscheurnaden zijn afwezig bij de voorouderlijke onderorde van alle overige trilobieten, de Olenellina.
- zijrandig (proparian): de gezichtsscheurnaden lopen vanaf de achterpunt van het oog naar de zoom, dus voor de achterhoek van het kopschild.
- kophoekig (gonatoparian): de gezichtsscheurnaden lopen vanaf de achterpunt van het oog naar de achterhoek van het kopschild.
- achterrandig (opistoparian): de gezichtsscheurnaden lopen vanaf de achterpunt van het oog naar de achterrand van het kopschild.
- randvolgend (hypoparian of marginal): de gezichtsscheurnaden vallen samen met de randscheurnaad. In dit geval is de afwezigheid een nieuwe ontwikkeling die voorkomt bij veel (maar niet alle) oogloze trilobieten.

zeegroen = vrije wang, lichtgrijs = vaste wang, middelgrijs = glabella
De primitieve staat van de gezichtsscheurnaden is zijrandig. Achterrandige gezichtsscheurnaden zijn meermaals onafhankelijk ontstaan. Er zijn geen voorbeelden van zijrandige gezichtsscheurnaden in soorten met voorouders met achterrandige gezichtsscheurnaden. Trilobieten met achterrandige gezichtsscheurnaden als volwassenen, hebben gewoonlijk larvale stadia met zijrandige gezichtsscheurnaden (bekende uitzonderingen zijn Yunnanocephalus en Duyunaspis).[17] Ook randvolgende gezichtsscheurnaden zijn onafhankelijk in verschillende groepen trilobieten ontstaan.
Overigens varieert het verloop van de gezichtsscheurnaad vanaf de voorpunt van het oog minstens even sterk als vanaf de achterpunt, maar door het ontbreken van een duidelijk referentiepunt vergelijkbaar met de kophoek is een scherpe categorisering daarvoor niet mogelijk. Een van de meer uitgesproken situaties is dat de voorkant van de gezichtsscheurnaad de voor- of zijrand niet elk voor zich doorsnijden, maar het allervoorste stuk van beide op de middenas samenvalt, zoals onder andere het geval is bij de Asaphida. Nog extremer is het als de voorste delen van de gezichtsscheurnaden in elkaar overlopen en de voorrand niet bereiken, zodat de vrije wangen als een juk met elkaar zijn vergroeid. Dit is bijvoorbeeld de situatie bij het geslacht Triarthrus, en bij de Phacopidae waar de gezichtsscheurnaden niet meer functioneel zijn.
Gehemelte
[bewerken | brontekst bewerken]
Het gehemelte (of hypostoom) is een verkalkte plaat aan de buikzijde van kopschild, meestal onder de voorkant van de glabella. De hypostoom kan worden ingedeeld in drie typen afhankelijk van of deze zijn bevestigd aan het middelste/voorste deel van de doublure of niet, en of de voorrand ervan samenvalt met de voorste rand van de glabella.
- zwevend (of natant): het hypostoom is niet aan de doublure bevestigd, de voorkant correspondeert met de voorkant van de glabella.
- gekoppeld (of conterminant): het hypostoom is aan de voorkant bevestigd aan de doublure, en de voorkant correspondeert met de voorkant van de glabella.
- onafhankelijk (of impendent): het hypostoom is aan de voorkant bevestigd aan de doublure, maar de voorkant correspondeert niet met de voorkant van de glabella.

Ogen
[bewerken | brontekst bewerken]



Trilobieten zijn uniek omdat ze de enige groep dieren zijn waarvan bekend is dat de lenzen van hun samengestelde ogen bestaan uit calciet. De facetten van de vroegste trilobieten vormen samen een halve cirkel in het horizontale vlak, die zo een tweedimensionaal beeld geven van de zeebodem waarop ze leefden. Trilobieten hebben nooit meer dan twee ogen, in tegenstelling tot andere geleedpotigen die soms in aanvulling op hun facetogen ook eenlenzige oogjes (ocelli) hebben, zoals insecten, duizendpoten, spinnen, schorpioenen en kreeftachtigen. Er zijn ook complexe samengestelde ogen bekend uit het Onder-Cambrium die geen lenzen van calciet hebben en daarom niet van een trilobiet afkomstig kunnen zijn, maar die ogen zijn ook complexer dan trilobietenogen uit dezelfde tijd. Deze ogen hadden de vorm van halve bollen en vormden dus een driedimensonaal beeld, en zijn misschien afkomstig van een dier dat in de waterkolom zwom.[18] Zulke ogen zijn pas later, tijdens het Ordovicium, bekend van trilobieten die in de waterkolom zwommen, zoals de Cyclopygidae en de Telephinidae. Bij de fossielen van vroege trilobieten zijn de ogen meestal afwezig. Dat komt omdat er een scheurnaad direct om het oog heen loopt en het oppervlak van het oog afzonderlijk werd vernieuwd van de rest van het exoskelet. Bij latere trilobieten zijn die oogscheurnaden verdwenen.
Er worden drie oogtypen onderscheiden:
- Het holochroale oogtype bestaat uit een aaneengesloten hexagonaal plaveisel van lenzen van zuiver kristallijn calciet met een gemeenschappelijk hoornvlies. Dit is het oorspronkelijke oogtype waarvan beide andere types zijn afgeleid. Het komt voor bij alle trilobieten die ogen hebben met uitzondering van de Phacopina, en (een deel van?) de Eodiscina. Dit holochroale oog is een zogenoemd appositieoog. Elke lens verzamelt licht van een heel smalle kegel recht voor en projecteert dat direct tot een beeldpunt op een groepje lichtgevoelige cellen. In de hersenen worden de signalen uit elk facet gecombineerd tot een beeld. Dit type oog is heel geschikt voor het waarnemen van beweging. De lenzen van trilobieten zijn doorgemeten en ze hebben in water inderdaad dezelfde werking als de lenzen van schaaldieren en insecten. Het aantal lenzen per oog kan van soort tot soort sterk verschillen van vele tienduizenden tot slechts een, en ook de doorsnede van de lenzen varieert, meestal tussen 30 en 100 µm. Voor welke omstandigheden de samengestelde ogen van trilobieten geschikt zijn kan worden gereconstrueerd omdat de ruimtelijke oriëntatie van de facetlenzen in het oog vaak behouden is gebleven. De combinatie van de diameter van elke facetlens (D) met de hoek tussen de assen van naburige facetten (Δφ) geeft een goede voorspelling van de omstandigheden waarin het oog wordt gebruikt. Bij moderne geleedpotigen varieert DΔφ van 0,3 voor dieren actief in fel zonlicht tot 20 of meer voor dieren die in de diepzee leven of 's nachts actief zijn. Bij enkele soorten met het holochroale oogtype (Asaphus cornutus, Isotelus gigas en Homotelus sp.) valt de waarde voor DΔφ in dezelfde categorie als bij moderne geleedpotigen die ‘s nachts actief zijn.[19]
- Het schizochroale oogtype bestaat uit maximaal 700 afzonderlijk geplaatste, relatief grote lenzen (tot 0,7 mm) die bestaan uit een buitenlens van heel zuiver kristallijn calciet en een binnenlens van minder zuiver kristallijn calciet die een verschillende brekingsindex hebben en in elkaar overgaan op een gebogen vlak. Op deze manier wordt sferische aberratie sterk verminderd en wordt een scherper beeld geprojecteerd, maar kunnen ook reflecties worden vermeden waardoor de lichtopbrengst van het lenssysteem groter wordt.[20] De huid (of sclera) rond de lenzen is dikker dan de lenzen, die elk een afzonderlijk hoornvlies hebben dat dieper dan de huid in het onderliggende weefsel steekt. Dit oogtype komt uitsluitend voor bij de soorten die horen bij de Phacopina, mits ze ogen hebben. Bij twee ondersoorten met schizochroale ogen (Phacops rana crassituberculata en P. rana milleri) varieert DΔφ van 10 tot meer dan 150, veel groter dan in enige levende geleedpotige. Dit impliceert dat die relatie tussen DΔφ en lichtomstandigheden niet geldt voor het schizochroale oogtype. Mogelijk vormt iedere lens in een schizochroaal oog een ruimtelijk beeld. Daarmee zouden de gezichtsvelden van de afzonderlijke lenzen in schizochroale ogen met die van diverse andere overlappen.[19] Dit zou consistent zijn met de zeer grote cilindervormige ogen van Erbenochile erbeni waar de wenkbrauw een stukje overheen steekt, misschien als een zonnescherm. Recent zijn echter analyses gedaan op basis van opnames met μct-scanning, synchrotron straling en zeer dunne plakjes in een lichtmicroscoop, die lijken te suggereren dat er geen volwaardig netvlies achter elke lens aanwezig was maar slechts 6 of 7 heel grote zintuigcellen (ongeveer 80 μm in doorsnede), een situatie die vergeleken zou kunnen worden met de zijdelingse ogen van degenkrabben.[21][22]
- Het abatochroale oogtype bestaat uit tot zo'n 70 wat kleinere lenzen die elk hun eigen hoornvlies hebben dat ophoudt aan de rand van de lens en niet in het onderliggende weefsel steekt, maar waar de tussenliggende huid minder dik is dan de lenzen. Dit type komt voor bij soorten van de Eodiscina. Er lijken bij soorten van de Eodiscina overgangen voor te komen tussen holochroale en abatochroale ogen. Heel veel Eodiscina hebben overigens geen ogen.
Er wordt van uitgegaan dat zowel het schizochroale als het abatochroale oog uit het holochroale oog zijn ontstaan als gevolg van pedomofose, het behoud van de larvale toestand in het volwassen individu, omdat de lenzen van holochroale ogen in eerste instantie ook niet aan elkaar raakten als een plaveisel, maar gescheiden zijn.
Lijf
[bewerken | brontekst bewerken]
Tussen het kopschild en het staartschild ligt een reeks scharnierende segmenten die samen het lijf of thorax worden genoemd. Het aantal segmenten bij de meeste soorten varieert van 2 tot 16, maar een aantal vroege soorten had een veel groter aantal thoraxsegmenten, tot een exemplaar van Balcoracania dailyi met 103.
Elk segment bestaat uit een bolle ring in het midden en ribben (of pleurae) aan de buitenkant, die de ledematen en kieuwen daaronder beschermen. De ribben zijn aan hun uiteinde soms afgerond, spits of eindigen in een kortere of langere stekel. Op het binnen/onderoppervlak (ventrale oppervlak) van het exoskelet staan uitsteeksels (of apodemes) waar spieren aangehecht zijn waarmee waarschijnlijk de poten bewogen worden, hoewel sommige beenspieren direct vastgemaakt zouden zijn aan het doorlopende binnenoppervlak van het exoskelet. Doordat het achterste segment van het kopschild soms op thoraxsegmenten lijkt - en dat geldt ook voor de segmenten in het staartschild - kan het lastig zijn te bepalen hoeveel segmenten het lijf nu heeft.
Bij vroege trilobieten (de onderordes Olenellina en Redlichiina) is het lijf regelmatig gedifferentieerd in een voorlijf (of prothorax) en een achterlijf (of opisthothorax), waarvan de segmenten smaller en minder lang zijn en de ribben kleiner ten opzichte van de as. Eveneens bij vroege trilobieten zijn de ribben van het derde segment vaak veel forser dan die van de andere segmenten en dragen soms veel stevigere en langere naar achter gebogen stekels aan hun einde. Bij één kleine familie zijn het vijfde en zesde segment vergroeid en dragen samen een grote stekel.
Staartschild
[bewerken | brontekst bewerken]
Het staartschild (of pygidium) is gevormd uit een aantal segmenten en het telson die met elkaar zijn vergroeid. De segmenten in de pygidium lijken op die in de thorax en dragen ook de karakteristieke twee-armige poten, maar kunnen niet scharnieren. Trilobieten worden in de wetenschappelijke literatuur ingedeeld naar de relatieve grootte van het staartschild ten opzichte van het kopschild met de Engelse termen "micropygous" (staartschild kleiner dan kopschild), "isopygous" (staartschild en kopschild even groot), en "macropygous" (staartschild groter dan kopschild). De grens tussen de segmenten is soms niet meer zichtbaar, het vaakst in de zijlobben, maar ook wel in de as en uiteindelijk kan ook onduidelijk worden waar de as in de zijlobben overgaat, het resultaat van een proces dat uitvlakking ("effacement") wordt genoemd.
Zachte lichaamsdelen
[bewerken | brontekst bewerken]


Van slechts omstreeks twintig soorten zijn niet verkalkte lichaamsdelen bewaard gebleven, zodat soms moeilijk bepaald kan worden wat hun variabiliteit is geweest. Zo zijn de antenne-achtige cerci aan het achtereind van het lichaam alleen bekend van de soort Olenoides serratus.[1][23][24][25]
Ledematen
[bewerken | brontekst bewerken]Trilobieten hadden voor de mond een enkel paar onvertakte antennes, bestaande uit tientallen korte segmenten, en voor de overige, weinig van elkaar verschillende tweearmige ledematen, die vermoedelijk homoloog zijn aan die in andere vroege geleedpotigen. Er zijn 2 tot 4 paar onder het kopschild, gevolgd door een paar per thoraxsegment en nog een aantal onder het staartschild.[24] Elke ledemaat bestaat uit een stevige schouder of coxa aan de basis (of proximaal), die zich onder de as van het rugschild bevindt. De binnenkant van de coxa (of gnathobasis) draagt doorns, waarschijnlijk om voedsel te vermalen en naar voren naar de mond te vervoeren. Aan de top van de coxa zitten twee armen. De binnenste arm (endopodiet of looppoot) heeft 6 of 7 segmenten, waarvan het verst van het lichaam verwijderde segment (of distaal) gewoonlijk klauwen of stekels had die vermoedelijk voor grip zorgen. De buitenste arm (exopodiet of kieuwpoot) is veerachtig vertakt en werd waarschijnlijk gebruikt voor de ademhaling en in sommige soorten of larvale stadia gebruikt voor het zwemmen. De vele haren op de poten suggereren dat ze een functie kunnen vervullen bij het lopen, het vasthouden of vervoer van voedsel of bij het waarnemen van de omgeving.[14] De best gefossiliseerde ledematen zijn die van de larven van Agnostus pisiformis (Agnostida). Die wijken echter sterk af van de ledematen van volwassen exemplaren uit de andere trilobietenordes, bijvoorbeeld omdat de exopodiet niet veerachtig vertakt is, maar een soort hark draagt, zodat deze waarschijnlijk een rol speelt bij het verzamelen van zwevende voedseldeeltjes en niet bij de ademhaling. Ledematen van volwassen Agnostida en van larven van andere ordes zijn echter niet bekend zodat onduidelijk is of de ledematen van Agnostida sterk verschilden van die van andere trilobieten of dat de ledematen tijdens de larvale ontwikkeling sterk veranderden.[26]
Organen
[bewerken | brontekst bewerken]De mond van trilobieten is naar achteren gericht en ligt aan de achterrand van het gehemelte, maar voor de poten onder het kopschild. De mond geeft vermoedelijk toegang tot een krop die verder naar voren lag dan de mond, onder de glabella. Daarvandaan leidt de darm achterwaarts naar de anus aan de achterpunt van het staartschild. Er wordt verondersteld dat de ledematen het voedsel naar voren vervoeren en zo nodig hebben vermalen doordat de doorns op de schouder het uit elkaar trokken en de binnenkant van de schouders wordt daarom ook kaakbasis of gnathobases genoemd. Detrituseters hebben misschien het sediment met de poten of het achterlijf losgeschopt om de voedseldeeltjes eruit op te woelen. Vleeseters, die in een keer een grote prooi moesten opeten, hadden waarschijnlijk een grote glabella die ruimte gaf aan een grote krop, in combinatie met een onafhankelijk gehemelte (of impendent hypostoom). Voor de aanwezigheid van andere inwendige organen zoals bloedvatenstelsel, zenuwknopen en lever zijn slechts onduidelijke aanwijzingen gevonden.[14]
Spieren
[bewerken | brontekst bewerken]Zeldzame sporen suggereren lange spieren in de lengterichting van het lichaam die zorgden voor het oprollen en andere spieren verantwoordelijk voor het bewegen van afzonderlijke poten.[25]
Ontwikkeling gedurende het leven
[bewerken | brontekst bewerken]
A. protaspide, B.-E. meraspides
Embryo's van trilobieten zijn nog nooit gevonden, maar het lijkt alsof het rugschild al snel na het uitkomen met calciet werd versterkt. Net als bij andere geleedpotigen verloopt de ontwikkeling van trilobieten via een aantal larvale stadia, die in elkaar overgingen door middel van een vervelling, waarbij de veranderingen in uiterlijk geleidelijk tot stand komen. In het allereerste stadium, de zogenoemde phaselus, is er een schijfje waarin geen segmenten herkenbaar zijn. Het volgende stadium heet protaspide waarin er zes segmenten kunnen worden onderscheiden, waarvan de voorste vijf samen de kop vormen en uit het achterste zowel het pygidium als de thorax ontstaan. Dan verandert de naam in meraspide, het stadium waarin zich telkens een segment aan de voorkant van het achterste segment afsplitst. Dat blijkt bijvoorbeeld uit het feit dat het derde thoraxsegment geteld vanaf de kop bij sommige groepen extra fors is en dit segment nadat het als derde in de tijd is ontstaan steeds op diezelfde positie blijft terwijl er achter telkens meer segmenten worden toegevoegd. Uiteindelijk ontstaan de segmenten die samengroeien tot het pygidium. Als ook het laatste segment is ontstaan spreken we van het volwassen stadium of holaspide. Ook na het bereiken van het volwassen uiterlijk groeit een trilobiet door vervellingen. Dan is er wel sprake van groei, maar zonder dat het aantal segmenten verandert.[9] Doordat een trilobiet vele malen in z'n leven vervelde zijn veruit de meeste van hun fossielen niet van complete dieren, maar (delen van) afgestoten pantsers.
Vindplaatsen van fossielen
[bewerken | brontekst bewerken]
Trilobieten zijn uitsluitend te vinden in afzettingen met fossielen van andere zoutwaterorganismen zoals brachiopoden, zeelelies en koralen, en we mogen dus aannemen dat het exclusief zeeorganismen zijn. Maar afgezien van de beperking tot mariene sedimenten worden trilobieten gevonden in uiteenlopende zeemilieus, variërend van lagunes tot aan de diepzee. Trilobieten zijn te vinden op alle continenten en in sedimenten uit elke oceaan uit het Paleozoicum. Naast resten van het exoskelet van de trilobieten zelf, worden ook kruip- en rustsporen gevonden, die worden toegeschreven aan trilobieten. Veel soorten trilobiet hadden een groot verspreidingsgebied en bestonden slechts een relatief korte periode. Hierdoor kunnen die soorten worden gebruikt als gidsfossiel.[27]
Omdat in Nederland aan de oppervlakte van nature geen gesteenten uit het Paleozoïcum voorkomen, worden daar alleen trilobieten aangetroffen die van elders afkomstig zijn, bijvoorbeeld in zwerfstenen uit Scandinavië uit het Ordovicium, grind in banken in de Maas en in natuursteen die door de mens wordt aangevoerd uit vooral de Ardennen, bijvoorbeeld als dijkbekleding.[28] In België zijn trilobieten vooral te vinden in een strook rond de Maas en de Samber met gesteentes uit het Devoon en rond Bergen en Charleroi uit het Carboon.[29]
Taxonomie
[bewerken | brontekst bewerken]De taxonomische rang van de vele groepen trilobieten blijkt uit de uitgang waarmee hun naam eindigt. Hieronder staat een overzicht van de verschillende rangen, van het ruimst verwant tot het meest verwant, voorzien van een voorbeeld.
| Uitgang van namen van trilobietengroepen van verschillende taxonomische rang | ||
| rang | uitgang | voorbeeld |
| klasse | -a | Trilobita |
| orde | -ida | Ptychopariida |
| onderorde | -ina | Eodiscina |
| superfamilie | -oidea | Acastoidea |
| familie | -idae | Cyclopygidae |
| onderfamilie | -inae | Cybelinae |
De 9 ordes van de trilobieten zijn: Agnostida, Redlichiida, Corynexochida, Lichida, Phacopida, Proetida, Asaphida, Harpetida en Ptychopariida. Soms worden ook de Nektaspida meegerekend, maar die hebben geen verkalkt exoskelet. Andere onderzoekers beschouwen de Agnostina niet als trilobieten en daarmee ook niet verwant aan de Eodiscina. Deze laatste onderorde wordt dan verheven tot orde en moet dan Eodiscida heten.
Geschiedenis van onderzoek en gebruik
[bewerken | brontekst bewerken]Dominee Edward Lhwyd publiceerde in 1698 in The Philosophical Transactions, het oudste wetenschappelijke tijdschrift in het Engels, een deel van zijn brief met betrekking tot verschillende door hem gevonden, regelmatig getekende stenen, voorzien van een pagina met etsen van fossielen.[30] Een van zijn etsen is van een trilobiet die hij vond bij Llandeilo, waarschijnlijk op het terrein bij het kasteel van Heer Dynefor, die hij beschreef als ... het skelet van een of andere platvis ....[31]
De ontdekking van Calymene blumenbachi, oorspronkelijk genoemd Dudley locust (Dudley-sprinkhaan) in 1749 door Charles Lyttleton, kan worden beschouwd als het begin van het wetenschappelijk onderzoek van trilobieten. Lyttleton schreef een brief aan de Royal Society of London in 1750 over een versteend insect dat hij had gevonden in de kalksteengroeve bij Dudley. In 1754 schreef Manuel Mendez da Costa dat de Dudley-sprinkhaan geen insect is, maar in plaats daarvan behoorde tot de stam der schaaldieren. Hij stelde voor om de vondsten van Dudley Pediculus marinus major trilobos (grote drielobbige zeeluis) te noemen, een naam die tot ver in de negentiende eeuw gebruikt werd. De Duitse theoloog en natuuronderzoeker Johann Walch, die als eerste een omvattende studie van deze groep uitvoerde, bedacht de naam trilobiet. Hij vond het passend om de naam ontlenen aan de unieke drielobbig karakter van de centrale as met een zijlob aan elke kant.[32]
De eerste beschrijving van een trilobiet dateert mogelijk uit de derde eeuw voor Christus, maar in ieder geval uit de vierde eeuw na Christus. De Spaanse geologen Eladio Liñán en Rodolfo Gozalo beweren dat sommige door Griekse en het Romeinse steenbewerkers als schorpioensteen, keversteen en miersteen aangeduide objecten in werkelijkheid fossielen van trilobieten zijn. Chinese bronnen geven minder dubbelzinnige verwijzingen naar trilobieten. Fossielen uit de Kushan-formatie in Noordoost-China werden gewaardeerd als inktsteen en voor de sier.[30]
Rond 1865 vonden Amerikaanse fossielenjagers gesteentelagen met grote hoeveelheden van Elrathia kingi in het westen van Utah. De Ute indianen, die deze trilobieten pachavee (klein waterbeestje) noemden, droegen ze tot in het begin van de twintigste eeuw als amulet. Door de kop werd een gat geboord en het fossiel werd gedragen aan een snoer om de nek.[33] Volgens de Ute zelf, beschermen deze amuletten tegen ziekten en kogels. In 1931 fotografeerde Frank Beckwith, reizend door de badlands twee rotstekeningen die hoogstwaarschijnlijk gemaakt zijn door de Ute en trilobieten voorstellen. Op dezelfde reis onderzocht hij een graf van onbekende ouderdom waar een doorboorde trilobiet in de borstholte van de overledene lag. Sindsdien zijn trilobiet-amuletten elders in het Grote Bekken gevonden, evenals in British Columbia en Australië.[30] In de jaren 1880 ontdekten archeologen in de Grotte du Trilobite, in de buurt van de Yonne, Frankrijk, een afgesleten fossiel van een trilobiet dat was doorboord alsof hij was bedoeld om te worden gedragen als een hanger. De laag waarin de trilobiet werd gevonden is gedateerd vijftienduizend jaar oud. Omdat de hanger zo was versleten, is niet duidelijk tot welke soort deze trilobiet hoort. Dit type trilobiet wordt echter niet gevonden in de buurt van de Yonne, dus is het denkbaar dat deze toentertijd hogelijk werden gewaardeerd, van elders aangevoerd en verhandeld.[30]
Externe link
[bewerken | brontekst bewerken]- ↑ a b c d e f g h Treatise on Invertebrate Paleontology, Part O, Arthropoda 1, Trilobita. Geological Society of America /University of Kansas Press, Boulder, CO/Lawrence, KA (1959), O38. ISBN 0-8137-3015-5.
- ↑ Cotton, T.J., Braddy, S.J. (2004). The phylogeny of arachnomorph arthropods and the origins of the Chelicerata. Transactions of the Royal Society of Edinburgh: Earth Sciences 94 (3): 169–193. ISSN: 0263-5933. DOI: 10.1017/S0263593303000105.
- ↑ Fortey, Richard. The Lifestyles of the Trilobites (PDF). American Scientist 92: 446–453. DOI: 10.1511/2004.49.944.
- ↑ Fortey, Richard (juni 2000). Olenid trilobites: The oldest known chemoautotrophic symbionts?. Proceedings of the National Academy of Sciences 97 (12): 6574–6548. DOI: 10.1073/pnas.97.12.6574.
- ↑ Hollingsworth, J.S. (2008). Advances in trilobite research. Instituto Geológico y Minero de España, Madrid, "The first trilobites in Laurentia and elsewhere".[dode link]
- ↑ Lieberman, B.S. (2002). Phylogenetic analysis of some basal early Cambrian trilobites, the biogeographic origins of the eutrilobita, and the timing of the Cambrian radiation. Journal of Paleontology 76 (4): 692–708. DOI: <0692:PAOSBE>2.0.CO;2 10.1666/0022-3360(2002)076<0692:PAOSBE>2.0.CO;2.
- ↑ Clarkson, E.N.K. (1998). Invertebrate Paleontology and Evolution, 4th. Wiley/Blackwell Science, Oxford, p. 452. ISBN 0-632-05238-4.
- ↑ Jaanusson, V. (1975). Evolutionary processes leading to the trilobite suborder Phacopina. Fossils and Strata 4: 209–218.
- ↑ a b Hughes (2007). The Evolution of Trilobite Body Patterning. Annual Review of Earth and Planetary Sciences. DOI: 10.1146/annurev.earth.35.031306.140258.
- ↑ a b Sam Gon III, Evolutionary Trends in Trilobites. A Guide to the Orders of Trilobites.
- ↑ B. Schoenemann, E.N.K. Clarkson, P. Ahlberg & M.E. Diez Alvarez (2010). A tiny eye indicating a planktonic trilobite. Palaeontology 53 (4): 695–701.
- ↑ Feist, R. (1991). The Late Devonian Trilobite crisis. Historical Biology 5: 197-214.
- ↑ Feist, R. (2002). Trilobites from the latest Frasnian Kellwasser Crisis in North Africa (Mrirt, central Moroccan Meseta). Acta Palaeontologica Polonica 47 (2): 203–210.
- ↑ a b c Whittington, H.B. (1997). Treatise on Invertebrate Paleontology, Part O, Arthropoda 1, Trilobita, revised. Volume 1: Introduction, Order Agnostida, Order Redlichiida. The Geological Society of America/The University of Kansas, Boulder, CO/Lawrence, KA. ISBN 0-8137-3115-1.
- ↑ The world's biggest trilobite: Isotelus rex new species from the Upper Ordovician of northern Manitoba, Canada[dode link] 99–112 (2003).
- ↑ Michael Kipping, Change of suit. www.trilobita.de. Geraadpleegd op 13 april 2011.
- ↑ Dai, T., Zhang, X. (2008). Ontogeny of the trilobite Yunnanocephalus yunnanensis from the Chengjiang lagerstätte, lower Cambrian, southwest China. Alcheringa 32: 465–468. ISSN: 0311-5518. DOI: 10.1080/03115510802418057.
- ↑ Lee, Michael S. Y., Jago, James B., García-Bellido, Diego C., Edgecombe, Gregory D., Gehling, James G. (2011). Modern optics in exceptionally preserved eyes of Early Cambrian arthropods from Australia. Nature 474: 631–634. DOI: 10.1038/nature10097.
- ↑ a b Fordyce, David, Cronin, Thomas W. (1993). Trilobite Vision: A Comparison of Schizochroal and Holochroal Eyes with the Compound Eyes of Modern Arthropods. Paleobiology 19 (3): 288-303.
- ↑ Horvat, Gabor (1996). The lower lens unit in Schizochroal trilobite eyes reduces reflectivity: on the possible optical function of the intralensar bowl. Historical Biology 12: 83-92.
- ↑ Schoenemann, Brigitte, Clarkson, Euan N.K. (2013). Discovery of some 400 million year-old sensory structures in the compound eyes of trilobites. Scientific Reports 3 (1429). DOI: 10.1038/srep01429.
- ↑ Horvat, Gabor, Clarkson, Euan N.K., Pix, Waltraud (1997). Survey of modern counterparts of schizochroal trilobite eyes: Structural and functional similarities and differences. Historical Biology 12: 229-263.
- ↑ Whittington, H.B. (1980). Exoskeleton, moult stage, appendage morphology, and habits of the Middle Cambrian trilobite Olenoides serratus. Palaeontology 23: 171–204.
- ↑ a b Trilobite tagmosis and body patterning from morphological and developmental perspectives 185–205 (2003).
- ↑ a b Bruton, D. L., Haas, W. (2003). Trilobites and Their Relatives: Contributions from the Third International Conference, Oxford 2001. Blackwell Publishing & Palaeontological Association, 331–348. ISBN 978-0-901702-81-4.
- ↑ Mueller, Klaus J., and Walossek, Dieter (1987). Morphology, Ontology and Life habit of Agnostus pisiformis from the Upper Cambrian of Sweden. Fossils and Strata 19: 1-124.
- ↑ Naturalis, Wat is een Gidsfossiel?. Natuurinformatie.
- ↑ Soortenlijst IJburg. fossilworks.
- ↑ Fossielenlocaties België. fossilworks.
- ↑ a b c d John J. McKay, The first trilobite. OhioLINK ETD Center. Geraadpleegd op 3 oktober 2012.
- ↑ Fortey, R.A. (2000). Trilobite! Eyewitness to Evolution. Knopf Doubleday Publishing Group. ISBN 9780375706219.
- ↑ Alex J. Chestnut, Using morphometrics, phylogenetic systematics and parsimony analysis to gain insight into the evolutionary affinities of the Calymenidae Trilobita. OhioLINK ETD Center. Gearchiveerd op 31 maart 2012. Geraadpleegd op August 21, 2011.
- ↑ Joleen Robinson (oktober 1970). Tracking the Trilobites.