Sharovipteryx
| Sharovipteryx Status: Uitgestorven Fossiel voorkomen: Midden- tot Laat-Trias | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|

| |||||||||||||||
| Taxonomische indeling | |||||||||||||||
| |||||||||||||||
| Geslacht | |||||||||||||||
| Sharovipteryx Sharov, 1971 | |||||||||||||||
| Typesoort | |||||||||||||||
| Sharovipteryx mirabilis | |||||||||||||||
| Afbeeldingen op | |||||||||||||||
| |||||||||||||||
Sharovipteryx[1] is een geslacht van uitgestorven reptielen behorend tot de Archosauromorpha, dat tijdens het Trias leefde in het gebied van het huidige Kirgizië. Sharovipteryx was een klein dier en had een vlieghuid tussen de achterpoten, kon minstens een glijvlucht uitvoeren en is verschillende malen in verband gebracht met de afkomst van de pterosauriërs.[2]
Naamgeving
[bewerken | brontekst bewerken]De soort werd in 1971 benoemd en beschreven door de Russische paleontoloog Alexandr Sjarov als Podopteryx mirabilis. De geslachtsnaam betekende 'voetvleugel', vanuit het Klassiek Griekse pous, 'voet' en pteryx, 'vleugel', en de soortaanduiding 'wonderlijk' in het Latijn, beide verwijzingen naar de verrassende bouw van het dier. De naam Podopteryx bleek echter al in 1871 bezet te zijn door een waterjuffer. Daarom werd de soort in 1981 door Richard Cowen hernoemd tot Sharovipteryx mirabilis; de nieuwe geslachtsnaam eert de oorspronkelijke beschrijver.
Vondstomstandigheden
[bewerken | brontekst bewerken]Het fossiel, holotype PIN 2584/8, bestaat uit een plaat schalie die doormidden gespleten is; beide delen tonen resten van een redelijk compleet en in verband liggend skelet, dat echter wel erg platgedrukt is. De splijting toont het dier van de rugkant; Sjarov dacht nog van de buikkant. Naast het skelet bevinden zich afdrukken van een vlieghuid. De aardlaag waarin het aangetroffen werd, behoort tot de Madjgen Switaformatie, een dikke afzetting van riviersedimenten, die gedateerd wordt in het Ladinien-Carnien, 230-225 miljoen jaar oud. In 1971 werd nog gedacht dat die ouder was, uit het Vroeg-Trias.

De vindplaats ligt bij Dzjaljau-Tsjo in het zuiden van Kirgizië, in de provincie Osj. Het fossiel werd bij toeval gevonden binnen het kader van een project om, gezien de uitstekende conservering die de laag kenmerkt, fossiele geleedpotigen te ontdekken; daarbij werd ook het al even opzienbarende reptiel Longisquama aangetroffen. Tot nu toe zijn geen andere vondsten van Sharovipteryx gedaan.
Beschrijving
[bewerken | brontekst bewerken]
Sharovipteryx is een klein dier van ongeveer vijftien à twintig centimeter lengte; die kan niet exact vastgesteld worden doordat het uiteinde van de staart ontbreekt. De staart maakt zo'n zestig procent van de lengte uit. De romp is zo'n drie centimeter lang en eindigt in een vrij lange nek die verbonden is aan een langwerpige spitse schedel; de afstand tussen snuit en staartbasis is zestig millimeter. De tongbeenderen zijn naar achteren verlengd. De armen liggen in het fossiel naast de romp en zijn niet goed bewaard gebleven. Ze waren ook slecht geprepareerd zodat hun precieze vorm en lengte nog onduidelijk is; meestal wordt aangenomen dat ze vrij kort zijn: Sjarov schatte ze tweemaal korter dan de benen, Tatarinov zelfs drie- tot viermaal. Volgens Unwin ging het zelfs niet om de armen maar om fout geïdentificeerde schachten van de voorste ribben; nieuwe preparatie lijkt aan te tonen dat de voorste ledematen toch aanwezig zijn. Het bekken is relatief klein, maar de hoogte ervan kan niet vastgesteld worden omdat het nog in het substraat naar beneden steekt. Er zijn vier sacrale wervels. Het darmbeen is naar voren verlengd. De benen liggen in het fossiel opgetrokken; ze zijn zeer lang met zowel een verlengd dijbeen als scheenbeen en dragen een brede voet met vijf tenen. De middenvoetsbeenderen zijn vrij kort en raken elkaar niet; de vijfde falanx is de langste: het been is zo gedraaid dat we van bovenaf op de voorkant van de voet kijken, zodat deze langste teen naar de staart gedraaid is. Het scheenbeen is iets langer dan het dijbeen; tussen beiden zijn aan beide zijden van het lichaam de afdrukken zichtbaar van een opgevouwen vlieghuid die loopt van de staartbasis tot aan de tenen. Daarboven, tussen beide dijbenen en het midden van de romp zijn ook weer afdrukken van zo'n huid te zien.
Taxonomie
[bewerken | brontekst bewerken]Sharovipteryx werd in 1971 door Sjarov in een eigen familie geplaatst, de Podopterygidae genaamd omdat hij nog de naam Podopteryx gebruikte. In 1989 hernoemde paleontoloog Leonid Petrovitsj Tatarinov die tot de Sharovipterygidae. Sharovipteryx is het enige bekende lid en de familie zegt dus niets over de verwantschappen van het dier. Sjarov wees het toe aan de Pseudosuchia, wat toen een verzamelnaam was die gebruikt werd voor 'primitieve' archosauriërs van onduidelijke herkomst.
Sjarov suggereerde dat het dier nauw verwant was aan de pterosauriërs en een soort voorloperfase van deze vliegende reptielen vertegenwoordigde. De vlieghuid van Sharovipteryx zou overeenkomen met het cruropatagium van de pterosauriërs, die ook tussen de vijfde tenen van beide voeten een membraan hebben dat niet aan de staart vastzit. Midden jaren tachtig echter hadden de eerste kladistische analyses als uitkomst dat de Pterosauria nauw verwant waren aan de Dinosauria, binnen de Ornithodira. Dat zou betekenen dat er geen speciale band bestond met Sharovipteryx, die veel te basaal leek om enige directe verwantschap met de dinosauriërs te hebben.
Meer gedetailleerd onderzoek, eind jaren negentig, door David Unwin toonde inderdaad aan dat, zoals Tatarinov al gesuggereerd had, Sharovipteryx zich ten opzichte van de dinosauriërs veel dieper in de stamboom bevond. Hij behoorde wel tot de Diapsida en meer bepaaldelijk tot de Archosauromorpha maar viel buiten de Archosauria in moderne zin (laat staan de Ornithodira), wat aangetoond werd door het ontbreken van belangrijke apomorfieën, nieuwe eigenschappen, van die groep, zoals een fenestra antorbitalis (schedelopening vóór de oogkas), een opening in de onderkaak en een vierde trochanter (uitsteeksel aan de achterkant van het dijbeen).
Volgens Unwin behoorde Sharovipteryx tot een basale groep archosauromorfen, de Prolacertiformes. Het dier had twee synapomorfieën, gedeelde nieuwe eigenschappen, van die groep: het bezit van zeven of minder nekwervels en een vierde middenvoetsbeen dat driemaal korter was dan het vijfde. Nog meer apomorfieën, nieuwe eigenschappen, werden gedeeld met verschillende meer afgeleide leden van de groep: de halswervels zijn langer dan de ruggenwervels; de doornuitsteeksels op de halswervels zijn lang en laag; de halsribben zijn lang en dun; het dijbeen is recht; het scheenbeen is langer dan het dijbeen; het tweede kootje van de vijfde teen is lang. Dat Sharovipteryx niet een nog basalere positie innam dan de prolacertiformen, werd bewezen door synapomorfieën van de klade, monofyletische afstammingsgroep, die bestaat uit de afstammelingen van de gemeenschappelijke voorouder van de Prolacertiformes en de Archosauria: een lange snuit, een smalle schedel, terugbuigende tanden, lange en smal toelopende ribben en verlengde zijuitsteeksels aan de wervels van de romp. Volgens een analyse was de soort het nauwst verwant aan de prolarcertiform Macrocnemus.
De dus goed ondersteunde plaatsing van Sharovipteryx binnen de Prolacertiformes zou ieder verband met de pterosauriërs hebben uitgesloten, ware het niet dat de oorsprong van die laatste groep zelf juist in die periode weer een twistpunt begon te worden. Christopher Bennett ontkende dat ze nauwe verwanten van de dinosauriërs waren en plaatste ze basaal in de Archosauriformes. Unwin ging nog een stapje verder en poneerde een verwantschap met de Prolacertiformes, waar ze Sharovipteryx dus weer tegenkwamen. De pterosauriërs leken in verschillende kenmerken op Sharovipteryx zoals in de holle botten, de proporties van de benen, de structuur van de voeten, de verlengde staartwervels, en de lange halswervels. Eén eigenschap wees wel heel speciaal op een verband: Unwin had ontdekt dat de afdruk van de vlieghuid van Sharovipteryx lange parallelle vezels van bindweefsel liet zien, een tot dan toe als uniek beschouwde aanpassing van de vlieghuid van de pterosauriërs. Deze structuren bij Sharovipteryx waren identiek in afmeting en vorm aan die bij de Pterosauria. Het was op zich waarschijnlijker dat zo'n gedeelde specialisatie geërfd was van een gemeenschappelijke voorouder, en dus homoloog was, dan dat die zich twee keer apart had ontwikkeld. Er is echter één probleem: pterosauriërs zijn wellicht wel nauw verwant aan de prolacertiformen, maar ze behoren, anders dan Unwin in 2000 nog dacht, vermoedelijk niet tot die groep als zodanig. Als de specifieke bindweefselstructuur van de vlieghuid geen geval is van convergente evolutie maar homoloog, moet de hoofdstamlijn van de archosauromorfen ter hoogte van de aftakking van de prolacertiformen dus uit vliegende, of minstens glijdende, vormen bestaan hebben. Een gelijkaardige hypothese ten aanzien van dinosauriërs, het Birds Came First-model van George Olshevsky, heeft geen genade kunnen vinden in de ogen van de paleontologische gemeenschap en evenzeer is er geen acceptatie geweest van een 'Pterosaurs Came First'-implicatie van de homologie van de vlieghuid van Sharovipteryx. De huidige consensus gaat dus uit van een verbluffende convergentie met als tweede mogelijkheid dat betere vondsten en analyses alsnog zouden kunnen aantonen dat de pterosauriërs wél prolacertiformen zijn.
Levenswijze
[bewerken | brontekst bewerken]Gezien zijn vlieghuid had Sharovipteryx het vermogen een glijvlucht uit te voeren. Dat zou erop wijzen dat hij in de bomen woonde. Met zijn lange benen zou hij zich krachtig van een tak hebben kunnen afzetten om na die sprong vrij vlak naar beneden — of naar een andere boom — te zeilen.[2] Zijn gewicht was laag, schattingen lopen van 4,5 tot 7,5 gram, en door de lage vleugelbelasting zou zo'n vlucht langzaam verlopen zijn zodat hij veilig kon landen — het gewicht beïnvloedt namelijk niet de glijhoek maar de voorwaartse snelheid. Deze standaardverklaring voor de morfologie van Sharovipteryx is echter problematisch gebleken.

Allereerst is het onduidelijk wat precies de vorm en totale oppervlakte van de vlieghuiden was. Tussen de achterbenen zat kennelijk het grootste membraan. Volgens Sjarov werd dat gerekt door de dijbenen licht op te trekken en de onderbenen uit te klappen, waarbij de achterrand een rechte hoek maakte met de lichaamsas. Tussen de dijbenen en armen zou nog een membraan gelopen hebben, in overeenstemming met Sjarov's hypothese dat Sharovipteryx een voorloper van de pterosauriërs was.[2] Een latere studie van Carl Gans echter, uit 1987, gaf een andere interpretatie: de onderbenen zouden ver naar voren gehouden zijn, haaks op de lichaamsas zodat de vlieghuid vanaf de voeten schuin naar achteren liep, een punt vormend die aan de staart bevestigd was. Er zou geen membraan aan de armen hebben vastgezeten of de dijen aan de romp verbonden hebben. Peter Wellnhofer nam in 1991 dit model wat de benen betreft over, maar voegde er kleine voorvleugeltjes aan toe. Al deze studies baseerden zich voornamelijk op intuïtie.
De eerste exacte modellering was van Gareth Dyke in 2006. Deze stelde vast dat de meest stabiele vlucht bereikt werd door het dier het profiel van een deltavlieger te geven waarbij de dijbenen haaks op het lichaam staan, de achterrand van de vleugel recht is, de onderbenen schuin naar achteren lopen en de op het fossiel zichtbare membranen vóór de dijbenen de lijn van de onderbenen naar voren voortzetten tot aan het lichaam, zodat de hele vleugel een driehoekige vorm krijgt. Het zwaartepunt van het lichaam zou wel nog steeds flink voor het punt gelegen hebben waar de vlieghuid de liftkracht uitoefende. Daardoor zou het dier zich niet vlak voor het doel even hebben kunnen 'optrekken' om veilig op de benen te landen, iets wat toch al een precair gebeuren moet zijn geweest omdat de benen waarop geland moest worden, meteen de dragers van de vleugel zijn. Aangezien er geen horizontaal staartvlak bekend is dat het stampen door een neerwaartse kracht uit te oefenen zou kunnen stabiliseren (overigens ook geen verticaal staartvaantje als richtingsroer, hoewel dat goed zou passen bij de verlengde staartwervels), nam Dyke aan dat de voorpoten een membraan droegen dat als een voorvleugel of canard werkte. Dat zou klein in oppervlakte hebben moeten zijn, minder dan tien procent van achterste 'delta', en niet aan de achterkant van de armen hebben moeten zitten maar als een propatagium tussen de bovenarmen en de nek gespannen een kleine voorste delta hebben moeten vormen. Sharovipteryx zou daarmee een uitstekende glijder geweest zijn, beter dan de huidige Draco, en een volledig unieke bouw hebben bezeten.
Dyke ging echter nog een stap verder. De vlieghuid van Sharovipteryx bezat een vezelstructuur die het mogelijk maakte de spanning onafhankelijk van de benen te reguleren en een profiel te handhaven als de benen naar boven bewogen werden. Verwijzend naar de mogelijke nauwe verwantschap met de pterosauriërs, zag hij Sharovipteryx daarom als een vertegenwoordiger van een fase in de ontwikkeling van het vluchtmechanisme van de Pterosauria, welke gekenmerkt zou zijn door een hoofdrol van de achterste ledematen in een klappende vlucht. Hij verwees daarbij naar een gelijkaardige mogelijkheid bij de evolutie van de vogels, waarop de lange veren op de achterpoten van Microraptor zou wijzen. Of Sharovipteryx zelf in staat zou zijn geweest zich, gebruikmakend van de geavanceerde vlieghuid, met zijn benen door de lucht voort te stuwen en dus een vlieger in plaats van een glijder geweest zou zijn, is moeilijk vast te stellen omdat een goede biomechanische analyse van deze mogelijkheid nog ontbreekt en we ook geen huidige dieren kennen waaraan we de noodzakelijke aanpassingen voor zo'n specialisatie kunnen aflezen. Het zou dan ongeveer vijftien slagen per seconde hebben moeten uitvoeren. Het vermogen tot een slagvlucht werd door Gans nog nadrukkelijk ontkend. Onder gunstige omstandigheden, thermiek bij windstilte, zou Sharovipteryx echter wel een langere zweefvlucht hebben kunnen uitvoeren.
Een tweede probleem is dat de lange poten van Sharovipteryx niet typisch zijn voor een klimmer. Het dier was evident een tweevoeter, die zich met zijn enorme benen snel lopend zou moeten hebben kunnen voortbewegen, zij het dat de vlieghuid dit wellicht wat had belemmerd. Dit zou er dus eerder op wijzen dat het membraan zich ontwikkeld heeft om een sprong op de grond te verlengen, iets wat ook gesuggereerd is als een mogelijk scenario voor de ontwikkeling van het vliegvermogen bij pterosauriërs. Aan de andere kant zou het dier met enige moeite toch wel hebben kunnen klimmen, met de benen in spreidstand om een stam geslagen.
Sharovipteryx was hoogstwaarschijnlijk een roofdier. Hij toont geen aanpassingen aan het eten van plantaardig voedsel. Zijn prooien moeten heel klein geweest zijn en meestal wordt dan ook aangenomen dat hij insecten of andere geleedpotigen at. Het is nog onduidelijk of hij die tijdens de vlucht had kunnen vangen; als hij alleen kon glijden zal dat zeer moeilijk geweest zijn. Een glijvermogen heeft zich geëvolueerd als veiligheidsmaatregel en voor het besparen van energie bij het verhuizen van boom tot boom.[2] Zelf kan hij in de lucht een prooi geweest zijn van de reusachtige libellen die tijdens het Trias voorkwamen.
Een zeer lastige vraag betreft het stofwisselingsniveau bij Sharovipteryx. Als tweevoeter werd zijn longfunctie niet belemmerd door zijdelingse bewegingen van het lichaam, zoals bij meer basale reptielen wel het geval was. De zuurstofvoorziening kan dus hoger gelegen hebben en wellicht bestond er een verband met een wat hoger metabolisme, hoewel dat voor het pure glijden niet echt nodig zou zijn geweest. Sporen van een isolerende vacht, zoals bij de pterosauriërs, zijn (nog) niet aangetroffen.
Alternatieve interpretaties
[bewerken | brontekst bewerken]De ontwikkeling van het vliegvermogen bij de Archosauromorpha, zowel bij de vogels als bij de pterosauriërs, is de laatste jaren het onderwerp geweest van een intensief debat, gevoerd onder grote publieke belangstelling. Er is een groep biologen die de heersende leer dat vogels dinosauriërs zijn, betwist. Binnen de opvattingen van deze BAND (Birds Are Not Dinosaurs) moet de oorsprong van de vogels ergens bij de 'primitieve archosauriërs' gelegen hebben. De vaagheid van dit begrip, het gebrek aan goede kandidaten, en het vermogen tot glijden van Sharovipteryx heeft ertoe geleid dat ook deze soort in de discussies opduikt, hoewel het verband hoogstens heel zijdelings kan zijn.
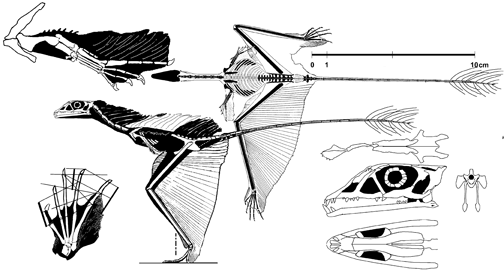
Een andere eigenzinnige invalshoek is gekozen door de amateurpaleontoloog David Peters. Gefrustreerd door het gebrek aan toegang tot de werkelijke fossielen begon deze foto's ervan te vergroten en bewerken, net zo lang totdat allerlei interessante structuren zichtbaar werden. Zo ontdekte hij bij Sharovipteryx alsnog de details van de armen en zelfs een extra membraan naast de keel. De rest van de paleontologische gemeenschap meent echter dat Peters niet anders heeft waargenomen dan het raster van de foto en de textuur van de omringende steen van het fossiel, die altijd wel de illusie van echte organen zullen opwekken als men maar voldoende bewerkingsstappen uitvoert. Peters traceert ook vaak de vorm van botten waar bij nader onderzoek helemaal geen bot aanwezig is, zoals in dit geval bij de armen. Ondanks deze totale afwijzing door de wetenschap is het Peters gelukt veel van zijn spectaculaire 'vondsten' in populair-wetenschappelijke bladen te laten afdrukken.
Literatuur
- Sharov, A. G., 1971, "Novjije ljetajoesjtsjije reptili iz mjezozoja Kazachstana i Kirgizii", Trudj Paleontologitsjeskogo Instituta, AN SSSR, 130, 104-113
- Cowen, R., 1981, "Homonyms of Podopteryx", Journal of Paleontology 55: 483
- Gans, C., Darevski, I. and Tatarinov, L.P., 1987, "Sharovipteryx, a reptilian glider?", Paleobiology 13: 415-426
- Tatarinov, L.P. 1989. [The systematic position and way of life of the problematic Upper Triassic reptile Sharovipteryx mirabilis.] Palaeontologicheskii Zhurnal 1989 (2): 110-112
- Tatarinov, L.P., 1994, "Terrestrial vertebrates from the Triassic of the USSR with comments on the morphology of some reptiles" Pp. 165–170 in J.-M. Mazin and G. Pinna (eds), Evolution, Ecology and Biogeography of the Triassic Reptiles, Paleontologia Lombarda, New Series, 2
- Peters, D., 1997, "A new phylogeny for the Pterosauria", J. Vertebr. Paleontol. 17, 69A
- Unwin, D. M., 2000, "Sharovipteryx: what can it tell us about the origin of pterosaurs?" 48th Symposium of Vertebrate Palaeontology and Comparative Anatomy, Portsmouth, England
- Unwin, D. M., 2000, "Sharovipteryx and its significance for the origin of the pterosaur flight apparatus", 5th European Workshop on Vertebrate Palaeontology
- Peters, D., 2000, "A Redescription of Four Prolacertiform Genera and Implications for Pterosaur Phylogenesis", Rivista Italiana di Paleontologia e Stratigrafia 106(3): 293-336
- Unwin, D. M., Alifanov, V.A. & Benton, M.J., 2003, "Enigmatic small reptiles from the Late Triassic of Kirghizia", In: Benton, M. J., Kurochkin, E., Shiskin, M. and Unwin, D. M., (eds), The Age of Dinosaurs in Russia and Mongolia, Cambridge University Press
- Peters, D., 2006, "The Front Half of Sharovipteryx", Prehistoric Times 76: 10-11
- Dyke, G.J., Nudds, R.L. and Rayner, J.M.V., 2006, "Flight of Sharovipteryx mirabilis: the world's first delta-winged glider." Journal of Evolutionary Biology, (2006): 1040-1043
Noten
- ↑ Fossilworks: Sharovipteryx mirabilis. www.fossilworks.org. Geraadpleegd op 11-01-2023.
- ↑ a b c d Dixon, Dougal (2001). Dinosauriërs In De Lucht. Corona Gidsen, p. 4. ISBN 9054955945.