Protein membran

Protein membran ialah protein biasa yang merupakan sebahagian daripada, atau berinteraksi dengan, membran biologi. Protein membran dikelaskan dalam beberapa kategori luas bergantung pada lokasinya. Protein membran integral bersifat kekal di membran sel, dan boleh sama ada menembusi membran (transmembran) atau terpasang di satu lapisan membran (monotopik integral). Protein membran periferi terpasang secara sementara dengan membran sel.
Protein membran adalah lazim serta penting dari segi perubatan; kira-kira sepertiga daripada semua protein manusia adalah protein membran, dan ini adalah sasaran bagi lebih separuh daripada semua ubat.[1] Walau bagaimanapun, berbanding dengan kelas protein lain, penentuan struktur protein membran kekal sebagai cabaran, sebahagian besarnya disebabkan oleh kesukaran dalam mewujudkan keadaan eksperimen yang dapat mengekalkan konformasi protein yang betul secara berasingan daripada persekitaran asalnya.
Fungsi
[sunting | sunting sumber]Protein membran melakukan pelbagai fungsi penting untuk kemandirian organisma:[2]
- Protein reseptor membran menyampaikan isyarat antara persekitaran dalaman dan luaran sel.
- Protein pengangkutan menggerakkan molekul dan ion merentasi membran. Ia boleh dikategorikan mengikut pangkalan data Pengelasan Pengangkut.
- Enzim membran mungkin mempunyai banyak aktiviti, seperti oksidoreduktase, transferase atau hidrolase.[3]
- Molekul lekatan sel membolehkan sel mengenal pasti satu sama lain dan berinteraksi protein yang terlibat dalam tindak balas imun.
Penyetempatan protein dalam membran boleh diramalkan dengan pasti menggunakan analisis hidrofobik jujukan protein, iaitu penyetempatan jujukan asid amino hidrofobik.
Protein membran integral
[sunting | sunting sumber]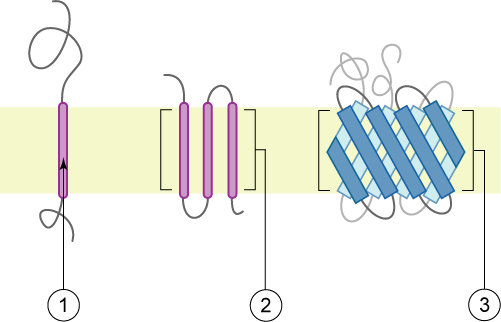
Membran diwakili dalam warna kuning pucat.
Protein membran integral melekat secara kekal pada membran. Protein sedemikian boleh diasingkan daripada membran biologi hanya menggunakan detergen, pelarut tak berkutub, atau kadangkala agen penyahaslian.[perlu rujukan] Mereka boleh dikelaskan mengikut hubungannya dengan dwilapisan:
- Protein politopik integral ialah protein transmembran yang merentangi membran lebih daripada sekali. Protein ini mungkin mempunyai topologi transmembran yang berbeza.[4][5] Protein ini mempunyai satu daripada dua seni bina struktur:
- Protein berkas heliks, yang terdapat dalam semua jenis membran biologi;
- Protein balang beta yang hanya terdapat dalam membran luar bakteria negatif Gram, dan membran luar mitokondrion dan kloroplas.[6]
- Protein bitopik ialah protein transmembran yang merentangi membran sekali sahaja. Heliks transmembran daripada protein ini mempunyai taburan asid amino yang berbeza dengan ketara kepada heliks transmembran berbanding protein politopik.[7]
- Protein monotopik integral ialah protein membran integral yang melekat di satu sisi membran sahaja dan bukan kedua-duanya.
Protein membran periferi
[sunting | sunting sumber]
Protein membran periferi terlekat secara sementara sama ada didwilapisan lipid atau pada protein integral melalui gabungan interaksi hidrofobik, elektrostatik dan ikatan bukan kovalen lain. Protein periferi tercerai selepas rawatan dengan reagen berkutub, seperti larutan dengan pH tinggi atau kepekatan garam yang tinggi.
Protein integral dan periferi boleh diubah suai selepas terjemahan protein dengan tambahan asid lemak, diasilgliserol[8], rantai prenil, atau GPI (glikosilfosfatidilinositol) yang mungkin dipaut di dwilapisan lipid.
Toksin polipeptida
[sunting | sunting sumber]Toksin polipeptida dan pelbagai peptida antibakteria seperti kolisin danhemolisin, dan protein-protein apoptosis tertentu kadangkala dikelaskan dengan kategorinya yang tersendiri. Protein-protein ini adalah larut air, tetapi boleh berkumpul dan bersekutu dengan membran, sama ada secara berbalik atau tidak.[perlu rujukan]
Dalam genom
[sunting | sunting sumber]Protein membran, seperti protein globul larut, protein gentian dan protein tidak teratur, ialah komponen kelaziman.[9] Dianggarkan bahawa 20-30% daripada semua gen dalam kebanyakan genom mengekod bagi protein membran.[10][11] Sebagai contoh, kira-kira 1000 daripada ~4200 protein E. coli dianggap sebagai protein membran, dan 600 daripadanya telah disahkan secara eksperimen sebagai pemastautin membran.[12] Dalam manusia, pemikiran semasa berpendapat bahawa sepenuhnya 30% genom mengekod protein membran.[13]
Dalam penyakit
[sunting | sunting sumber]Protein membran menjadi sasaran lebih 50% daripada semua ubat perubatan moden.[1] Antara penyakit manusia yang melibatkan protein membran adalah penyakit jantung, Alzheimer dan fibrosis sista.[13]
Penulenan
[sunting | sunting sumber]Walaupun protein membran memainkan peranan penting dalam semua organisma, penulenannya secara sejarah, dan terus menjadi, cabaran besar bagi saintis protein. Pada tahun 2008, 150 struktur unik protein membran telah tersedia,[14] dan menjelang 2019, hanya 50 protein membran manusia telah dijelaskan strukturnya.[13] Sebaliknya, kira-kira 25% daripada semua protein ialah protein membran.[15] Permukaan hidrofobik mereka menyukarkan pencirian struktur dan fungsinya secara lanjutan.[13][16] Detergen boleh digunakan untuk menjadikan protein membran larut dalam air, tetapi ini juga boleh mengubah struktur dan fungsi protein.[13] Penghasilan protein membran yang larut air juga boleh dicapai melalui kejuruteraan jujukan protein dengan menggantikan asid amino hidrofobik terpilih dengan asid hidrofilik sambil berhati-hati untuk mengekalkan struktur sekunder sambil menilai semula keseluruhan cas.[13]
Kromatografi pertalian adalah salah satu penyelesaian terbaik dalam penulenan protein membran. Aktiviti protein membran berkurangan dengan sangat cepat berbeza dengan protein lain.[perlu rujukan] Dengan itu, kromatografi afiniti menyediakan penulenan protein membran yang cepat dan spesifik. Teg polihistidina ialah teg yang biasa digunakan dalam penulenan protein membran,[17] dan teg rho1D4 alternatif juga telah berjaya digunakan.[18][19]
Rujukan
[sunting | sunting sumber]- ^ a b "How many drug targets are there?". Nature Reviews. Drug Discovery (Opinion). 5 (12): 993–6. December 2006. doi:10.1038/nrd2199. PMID 17139284.
- ^ "Mapping the human membrane proteome: a majority of the human membrane proteins can be classified according to function and evolutionary origin". BMC Biology. 7: 50. August 2009. doi:10.1186/1741-7007-7-50. PMC 2739160. PMID 19678920.
- ^ "The substitution of Arg149 with Cys fixes the melibiose transporter in an inward-open conformation". Biochimica et Biophysica Acta (BBA) - Biomembranes. 1828 (8): 1690–9. August 2013. doi:10.1016/j.bbamem.2013.03.003. PMID 23500619 – melalui Elsevier Science Direct.
- ^ "Membrane-protein topology". Nature Reviews. Molecular Cell Biology. 7 (12): 909–18. December 2006. doi:10.1038/nrm2063. PMID 17139331.
- ^ Gerald Karp (2009). Cell and Molecular Biology: Concepts and Experiments. John Wiley and Sons. m/s. 128–. ISBN 978-0-470-48337-4. Dicapai pada 13 November 2010 – melalui Google Books.
- ^ "Assembly of β-barrel proteins into bacterial outer membranes". Biochimica et Biophysica Acta (BBA) - Molecular Cell Research. 1843 (8): 1542–50. August 2014. doi:10.1016/j.bbamcr.2013.10.009. PMID 24135059 – melalui Elsevier Science Direct.
- ^ "Charged residues next to transmembrane regions revisited: "Positive-inside rule" is complemented by the "negative inside depletion/outside enrichment rule"". BMC Biology. 15 (1): 66. July 2017. doi:10.1186/s12915-017-0404-4. PMC 5525207. PMID 28738801.
- ^ "Structure of the alternative complex III in a supercomplex with cytochrome oxidase". Nature. 557 (7703): 123–126. May 2018. Bibcode:2018Natur.557..123S. doi:10.1038/s41586-018-0061-y. PMC 6004266. PMID 29695868.
- ^ "SCOP2 prototype: a new approach to protein structure mining". Nucleic Acids Research. 42 (Database issue): D310-4. January 2014. doi:10.1093/nar/gkt1242. PMC 3964979. PMID 24293656.
- ^ Liszewski, Kathy (1 October 2015). "Dissecting the Structure of Membrane Proteins". Genetic Engineering & Biotechnology News (paper). 35 (17): 1, 14, 16–17. doi:10.1089/gen.35.17.02.
- ^ "Predicting transmembrane protein topology with a hidden Markov model: application to complete genomes" (PDF). Journal of Molecular Biology. 305 (3): 567–80. January 2001. doi:10.1006/jmbi.2000.4315. PMID 11152613. Diarkibkan daripada yang asal (PDF) pada 2020-08-04 – melalui Semantic Scholar.
- ^ "Global topology analysis of the Escherichia coli inner membrane proteome". Science (Report). 308 (5726): 1321–3. May 2005. Bibcode:2005Sci...308.1321D. doi:10.1126/science.1109730. PMID 15919996.
- ^ a b c d e f Martin, Joseph; Sawyer, Abigail (2019). "Elucidating the Structure of Membrane Proteins". Tech News. BioTechniques (Print issue). Future Science. 66 (4): 167–170. doi:10.2144/btn-2019-0030. PMID 30987442.
- ^ "Overcoming the challenges of membrane protein crystallography". Current Opinion in Structural Biology. 18 (5): 581–6. October 2008. doi:10.1016/j.sbi.2008.07.001. PMC 2580798. PMID 18674618.
- ^ "Predicting transmembrane protein topology with a hidden Markov model: application to complete genomes" (PDF). Journal of Molecular Biology. 305 (3): 567–80. January 2001. doi:10.1006/jmbi.2000.4315. PMID 11152613. Diarkibkan daripada yang asal (PDF) pada 2020-08-04 – melalui Semantic Scholar.
- ^ "Membrane proteins: always an insoluble problem?". Biochemical Society Transactions. 44 (3): 790–5. June 2016. doi:10.1042/BST20160025. PMC 4900757. PMID 27284043.
- ^ "Genetic Approach to Facilitate Purification of Recombinant Proteins with a Novel Metal Chelate Adsorbent". Nature Biotechnology. 6 (11): 1321–1325. November 1988. doi:10.1038/nbt1188-1321.
- ^ "Expression, surface immobilization, and characterization of functional recombinant cannabinoid receptor CB2". Biochimica et Biophysica Acta (BBA) - Proteins and Proteomics. 1834 (10): 2045–56. October 2013. doi:10.1016/j.bbapap.2013.06.003. PMC 3779079. PMID 23777860.
- ^ "Large-scale production and study of a synthetic G protein-coupled receptor: human olfactory receptor 17-4". Proceedings of the National Academy of Sciences of the United States of America. 106 (29): 11925–30. July 2009. Bibcode:2009PNAS..10611925C. doi:10.1073/pnas.0811089106. PMC 2715541. PMID 19581598.