中胚葉
| 中胚葉 | |
|---|---|
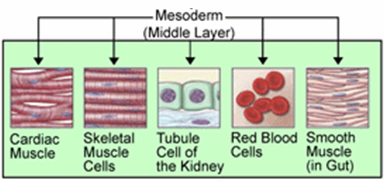 中胚葉由来の組織 | |
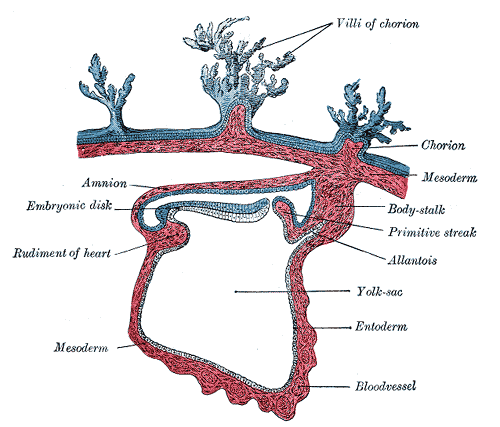 胚の区分 | |
| グレイ解剖学 | subject #6 49 |
| 日齢 | 16 |
| MeSH | Mesoderm |
中胚葉(ちゅうはいよう 英:mesoderm)とは、動物の発生初期に区別される細胞群の名称である。外胚葉と内胚葉の間を埋めるように発達し、筋肉や体腔などを作る。中胚葉を持つ動物を三胚葉性動物という。
概要
[編集]動物の発生では卵割が進むと内部に卵割腔を持つ胞胚期についで、内外の細胞層に分かれる原腸胚期に進む。そこでは細胞層が内側と外側の二層に分かれ、内部の細胞群は原腸を形成する。ここで外表の細胞層を外胚葉、その内側で原腸壁を構成する細胞群を内胚葉という。その際、多くの動物群ではこれと同時に、あるいはそれ以降の進行に従って、この二つの胚葉の間に形成される細胞群があり、これが中胚葉である。中胚葉の形成の方法は幾通りかの型があり、それは動物の系統と密接に関連すると考えられてきた。
外胚葉が表皮とその派生的な構造、および神経系に分化し、内胚葉が消化管とその派生的な構造に発達するのに対して、中胚葉はその両者の間を埋める非常に多様な構造に分化する。それはたとえば体腔や循環系、内骨格や筋肉、真皮などである。体節制のある動物では、体節もここから作られる。なお、脊索動物では脊索がここから形成されるが、これを中胚葉に含めない主張もある。
歴史
[編集]胚葉の発見はバンダーに遡る。彼は1817年にニワトリ胚の初期発生に於いて3つの層構造が存在することを示した。彼は2層がまず形成され、その後にその間にもう一層が形成されることまで観察していた。その後、彼の知人のフォン・バーがそれを脊椎動物全般に存在するものであることを示し、さらにラトケは同様の細胞層が無脊椎動物にも存在することを示した[1]。ベーアはこれらに基づいて胚葉説を提出した[2]。
形成の過程
[編集]中胚葉細胞が外胚葉と内胚葉の間を満たすものは無体腔動物、間に体腔を作って外胚葉の裏打ちのみを行うものは偽体腔動物、外胚葉・内胚葉の両側の裏打ちをするものを真体腔動物と言う。外胚葉と内胚葉の間にほとんど細胞の存在しないものを二胚葉性動物というが、実際に外皮と腸壁の間に細胞が存在しない例はヒドロ虫類しかなく、その意味では後生動物は全て三胚葉性とする主張もある[3]。
中胚葉がどこに由来するかは動物群によって様々である。大きく分けると外胚葉の細胞に由来するものと、内胚葉の細胞に由来するものがあり、前者を外中胚葉 (ectomesoderm)、後者を内中胚葉 (endomesoderm)という。真の中胚葉は後者であるとみなされ、たとえば二胚葉性動物と言われる刺胞動物と有櫛動物では外中胚葉のみを持つ。だが、これをあえて内中胚葉と区別しない立場もある[4]。
真体腔は、その発生の様式から、大きくは以下の二つに分けられてきた[5]。
- 腸体腔 (entocoel)
- 腸管の側面に腸体腔嚢と呼ばれる膨らみを生じ、これがくびれて独立したものから発達する。
- 裂体腔 (schizocoel)
- 中胚葉に当たる細胞が分かれて、その後のその内部に体腔が形成される。
- その典型的なものは環形動物に見られるもので、卵割の初期に形成される一個の端細胞(または中胚葉母細胞)が卵割腔内で分裂し、前方に向かって体節ごとの体腔を作ってゆく。
この二分法は古典的な動物系統論に基づくものである。それによると三胚葉性の動物は原口が口になる前口動物と原口が肛門になる後口動物に分かれ、前者は螺旋卵割を行い、裂体腔を持つのに対して、後口動物は放射卵割を行って腸体腔を発達させる。しかし、このような二分法は現在では疑問視されている[6]。
なお、脊椎動物も後口動物であるから腸体腔であり、その中胚葉は原腸壁から袋の形で分離するとの判断があった。それによると両生類の場合、神経胚期に脊索の両側で二層の壁を持つ袋の形で外胚葉と内胚葉の間に入り込み、その後に脊索との間でくびれるようにして分離する[7]。だが現在では脊椎動物は裂体腔であると判断されている[6]。
実際の発生では状況はさらに複雑である。たとえば発生のモデル生物として使われてきたウニの場合、胞胚腔を埋める細胞には以下のような三つの形成過程がある。どれも内胚葉起源と見なせるものの、それぞれ出現する時期も場所も異なるが、その全てが中胚葉と見なされている[8]。
- まず原腸陥入に先だって、植物極層から胞胚腔内に細胞群がこぼれ落ちる。これを第一次間充織と言い、原腸陥入が進むと原口近く、胞胚腔の植物極側の底に位置する。これは幼生の骨片を形成する。
- 原腸陥入の際に原腸の先端部から胞胚腔内にこぼれ落ちる細胞群があり、これを第二次間充織という。これは原腸を引き込む運動に与って原腸が陥入する方向の先端に位置し、後に筋肉や体腔細胞などに分化する。
- 原口の反対側に新たに口が開いた後、腸管の側面から膨らみを生じ、独立して体腔嚢となる。これは変態の際にウニ原基の形成される元になる。
分化
[編集]上記のように内胚葉が消化管、外胚葉が表皮と神経に分化する。つまり、おおよそ一番外側と一番内側の上皮をこの二つが作り上げる。中胚葉は両者を埋める構造の全てに分化し、たとえば結合組織はこれに当たる。体腔を持つ動物では体腔に面する上皮も中胚葉性である[9]。他に筋肉系、循環系、排出器、生殖巣も中胚葉から作られる。ウニの場合、中胚葉からは骨片、筋肉、色素細胞、水管系などが形成される。
脊椎動物の場合
[編集]以下、発生のモデル生物としてよく使われる両生類について説明する。他の脊椎動物では様々な違いはあるものの、基本的な特徴は共通している。
起源と発生
[編集]中胚葉に分化するのは、卵割の時期に赤道の帯域にある細胞群である[10]。
原腸胚期にはこの部分の細胞が内部の胞胚腔に侵入して原腸を形成する。その後、動物極側から入り込んだ細胞群は外層を裏打ちするように発達し、これが中胚葉となる[11]。
分化
[編集]中胚葉は原腸胚期から神経胚期にかけて、さらに分化する。一つの区分法として、これを中軸中胚葉・沿軸中胚葉・側板中胚葉と分ける方法がある。中軸中胚葉は脊索で、これは下記のように大部分の脊椎動物では発生初期のみ存在して後に消失するが、その働きと意味は重要である。この部分は両生類ではオーガナイザー領域から形成され、神経管を誘導し、それによって全身の体軸等を決めるものとなる。後二者は、一般には体節と側板と呼ばれる[12]。
普通は以下のように分ける。それらからさらに分化する器官と共に、以下に記す[13]。ただし、各部位・各器官は完全にそれぞれの部分からだけ形成されるわけではない。
- 脊索中胚葉 (chordamesoderm):背中側に位置する棒状の構造。神経管の下になる。
- すぐに退化消失し、脊椎に置き換わる。
- 体節中胚葉 (somatic mesoderm):神経管両側面に分離、次第に後方に向かって細胞塊を形成することで、体節構造を作る。神経管を包み、一部は外皮に沿って分化。
- 脊椎骨・骨格筋・真皮
- 硬節 (sclerotome):腹内側。椎骨や肋骨の軟骨、および背側大動脈の内皮細胞を形成する。
- 靭帯節 (syndetome):硬節中の最も背側。腱となる。
- 皮筋節 (dermamyotome):背外側
- 皮節 (dermatome):皮筋節の中央部。背側の真皮、筋芽細胞、褐色脂肪細胞を生じる。
- 筋節 (myotome):皮節の両側。内側は肋間筋などを含む近位筋を、外側は体壁、四肢、舌などの遠位筋を生じる。
- 腎節 / 中間中胚葉:側板の腹背外側に分化。
- 腎臓、および生殖腺に関わる輸送管
- 側板中胚葉 (lateral plate mesoderm):腸管の回りに発達。水平に2つの層(壁側中胚葉、臓側中胚葉)に分割されて内部に腔所を作り、これが体腔になる。
誘導について
[編集]中胚葉のうち、原口背唇部に由来する部分は、外胚葉に働きかけて神経を形成させる。これはシュペーマンとマンゴルドにより発見され、誘導と名付けられた。誘導現象は他にもあるため、現在ではこれを神経誘導と呼んでいる。その働きを持つ部分として、シュペーマンは原口背唇部を形成体、あるいはオーガナイザー(organizer)と呼んだが、往々にシュペーマンオーガナイザーと呼ばれる[14]。
上記のように、胞胚期以前には赤道域に位置する細胞群が中胚葉に分化するが、この区域の細胞の運命は胞胚期の直前までは決まっていないことも示されている。それが中胚葉に分化する機構として、植物極側の細胞による誘導があると考えられている。これはニューコープによる以下のような実験で示され、また分子生物学的にも裏付けされている[15]。
- 胞胚初期の胚の動物極側、植物極側の部分を切り出して単独で培養すると、それぞれ外胚葉、内胚葉と思われる細胞が分化する。ところが、両者を接触させて培養すると、その接触面の動物極側の細胞から、筋肉や血管など中胚葉細胞が分化する。
このように背側植物極領域はシュペーマンオーガナイザーを誘導するという重要なものであり、この部位はニューコープセンターと名付けられている[16]。
出典
[編集]- ^ 浅島・武田(2007),p.115
- ^ 木原・岡田編(1950)p.13
- ^ 白山(2000),p.17
- ^ 白山(2000)p.17
- ^ 白山(2000),p.20
- ^ a b 西川(2000),p.252
- ^ 古川・西沢代表(1969)p.66
- ^ 東中川他(2008),p.67-68
- ^ 白山(2000),p.16-20
- ^ 東中川他(2008),p.49
- ^ 浅島・武田(2007),p.41
- ^ 東中川他(2008),p.106-108
- ^ 浅島・武田(2007),p.43-44・東中川他(2008),p.108-115・Gilbert(2015),p.421-433
- ^ 浅島・武田(2007),p121
- ^ 東中川他(2008),p.83-84
- ^ 浅島・武田(2007),p119
参考文献
[編集]- 浅島誠・武田洋幸、『シリーズ21世紀の動物科学5 発生』、(2007)、培風館
- 東中川徹・八杉貞雄・西籠秀俊、『ベーシックマスター 発生生物学』,(2008)、オーム社
- 木原均・岡田要、『発生 現代の生物学 第2集』、(1950)、共立出版
- 古川秀男・西沢一俊代表、『原色現代科学大事典7 生命』、(1969)、学習研究社
- 白山義久、「総合的観点からみた無脊椎動物の多様性と系統」、『無脊椎動物の多様性と系統 バイオディバーシティ・シリーズ5』、2000、裳花房
- 西川輝昭、「前口動物・後口動物 二分岐節をめぐって」『無脊椎動物の多様性と系統 バイオディバーシティ・シリーズ5』、2000、裳花房
- Scott F. Gilbert 『ギルバート発生生物学』、阿形清和、高橋淑子 メディカル・サイエンス・インターナショナル、2015年。ISBN 978-4-89592-805-2。