Ipotesi del mondo a RNA
L'ipotesi del mondo a RNA è una teoria che propone una prima fase con la presenza di forme di vita basate esclusivamente sull'RNA (acido ribonucleico) che avrebbe preceduto la formazione degli attuali organismi viventi basati soprattutto sul DNA (acido desossiribonucleico). L'evoluzione all'attuale sistema basato sul DNA e proteine avrebbe portato notevoli vantaggi in termini di stabilità e flessibilità.
Storia
[modifica | modifica wikitesto]La dicitura mondo a RNA (RNA world nell'originale inglese) fu utilizzata per la prima volta dal premio Nobel Walter Gilbert nel 1986 in un articolo di commento sulle funzioni catalitiche di numerose forme di RNA che, proprio in quel periodo, iniziavano a essere messe in evidenza all'interno della comunità scientifica.[1] In ogni caso, l'idea di una vita a RNA indipendente da DNA e proteine era stata già formulata due decenni prima nel libro del 1968 The Genetic Code[2] di Carl Woese. Un'idea del genere era stata comunque già lanciata nel 1963 dal biologo molecolare Alexander Rich, del Massachusetts Institute of Technology, che ne parlò in un articolo inserito in un volume pubblicato in onore del premio Nobel Albert Szent-Györgyi[3].
L'ipotesi e le proprietà dell'RNA
[modifica | modifica wikitesto]Secondo l'ipotesi del mondo a RNA, tale macromolecola potrebbe esser stata originariamente l'unica responsabile della vita cellulare o pre-cellulare. Alcune teorie relative all'origine della vita presentano l'informazione e la catalisi mediata da RNA come primo passaggio nell'evoluzione della vita cellulare. L'RNA è infatti in grado di immagazzinare informazione ma, rispetto al DNA, è in grado anche di catalizzare reazioni come gli enzimi proteici.
L'ipotesi presuppone che tale sistema basato sull'RNA si sarebbe evoluto nel corrente sistema comprendente anche DNA e proteine grazie alla grande stabilità chimica del DNA (necessario per la conservazione della preziosissima informazione genica) e alla maggiore flessibilità catalitica che gli amminoacidi garantiscono. Secondo l'ipotesi del mondo a RNA, dunque, l'RNA ancora presente nelle cellule (nei ribosomi e nei ribozimi) è solo un residuo del mondo a RNA originale.
RNA come enzima
[modifica | modifica wikitesto]Gli RNA con funzione di enzima, o ribozima, sono possibili sebbene non comuni nell'odierna vita basata sul DNA. Tuttavia i ribozimi svolgono un ruolo importante; i ribozimi sono componenti essenziali del ribosoma, quest'ultimo è vitale per la sintesi proteica. Molte sono le funzioni possibili del ribozima: la natura utilizza ampiamente l'RNA self-splicing e l'evoluzione diretta ha creato ribozimi con una varietà di attività.
Tra le proprietà catalitiche più rilevanti relativamente all'origine della vita figurano:
- L'abilità di auto-duplicarsi, o di duplicare altre molecole di RNA. Molecole relativamente corte di RNA in grado di duplicarne altre sono state prodotte in laboratorio. La più corta a essere individuata è di 165 basi, sebbene si creda che ne possano bastare anche meno. La più fedele, di 189 basi, ha mostrato un'accuratezza del 98,9%,[4] che significa in parole povere che, replicando sé stessa, essa sarebbe in grado di realizzare una esatta copia ogni otto tentativi.
- L'abilità di catalizzare semplici reazioni chimiche, che rende possibile la creazione di nuove molecole. Filamenti relativamente corti con tali capacità sono stati realizzati in laboratorio.[5][6]
- L'abilità di formare legami peptidici e, quindi, brevi peptidi. Questa operazione è correntemente svolta dai ribosomi, complessi costituiti da proteine e due lunghe molecole di RNA (note come rRNA) ritenute le principali responsabili dell'attività di sintesi proteica. In laboratorio è stata sintetizzata una molecola in grado di realizzare brevi peptidi. Si può ipotizzare che gli attuali ribosomi possano essersi evoluti da molecole del genere.[7]. È stato anche suggerito che gli amminoacidi possano esser stati complessati all'inizio con molecole di RNA in qualità di cofattori in grado di amplificare e diversificare le capacità enzimatiche; l'mRNA potrebbe essersi evoluto da simili molecole e il tRNA da filamenti in grado di catalizzare il trasferimento degli stessi amminoacidi verso i brevi peptidi.[8].
RNA nella conservazione dell'informazione
[modifica | modifica wikitesto]L'RNA è una molecola molto simile al DNA, con due sole differenze chimiche. Tale somiglianza, ad esempio, permette di realizzare doppie eliche miste di DNA e RNA. Per tale motivo, è possibile ipotizzare un ruolo nella conservazione dell'informazione, tipica dei DNA, anche per gli RNA.
Confronto tra la struttura del DNA e dell'RNA
[modifica | modifica wikitesto]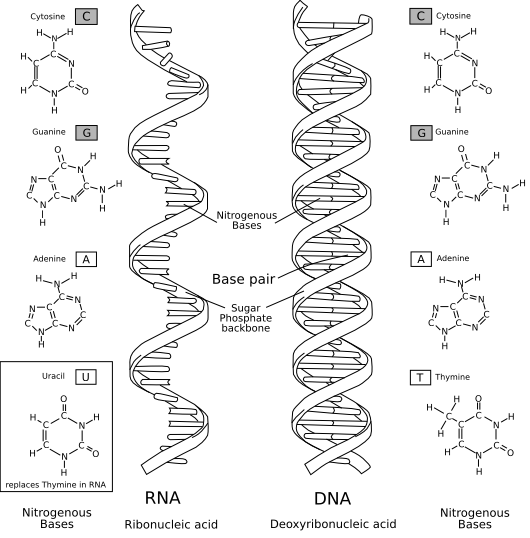
La principale differenza tra le molecole è la presenza di un gruppo ossidrile in posizione 2' del ribosio presente nella molecola di RNA. Tale gruppo forza il ribosio nella conformazione C3'-endo, a differenza della C2'-endo tipica del deossiribosio, generando così due molecole comunque differenti tra loro. Soprattutto, questo gruppo rende la molecola meno stabile, poiché può attaccare il vicino legame fosfodiesterico e romperlo.
L'altra differenza rilevante è il set di basi utilizzate dell'RNA, che comprende uracile al posto della timina usata dal DNA. Si tratta di molecole simili, sebbene l'uracile richieda meno energia per essere prodotto. Dal punto di vista dell'appaiamento non vi sono conseguenze rilevanti: l'adenina è in grado di legare entrambe le basi indifferentemente. Il vero limite dell'utilizzo dell'uracile consiste nel fatto che esso può derivare dalla deamminazione della citosina, rendendo le molecole di RNA particolarmente suscettibili a mutazioni che sostituiscono paia di basi come GC con GU.
Limiti nella conservazione dell'informazione nell'RNA
[modifica | modifica wikitesto]La conservazione di grandi quantità di informazione nell'RNA non è semplice. La struttura dell'RNA ne rende intrinsecamente fragili i lunghi filamenti, che possono andare incontro a degradazione tramite idrolisi. Le basi aromatiche, che assorbono efficientemente le radiazioni UV, sono inoltre molto prone a modificazioni strutturali, che rendono decisamente bassa l'accuratezza di tale conservazione.[9][10] Queste limitazioni non rendono impossibile la conservazione di informazione da parte dell'RNA, infatti esso è usato come materiale genetico da virus come i retrovirus e i viroidi. La presenza di una molecola ottimizzata come il DNA spiega come mai oggi l'RNA non venga utilizzato per tale scopo, ma non esclude che questo possa essere avvenuto nelle fasi primordiali della vita sulla Terra.
Valore dell'ipotesi
[modifica | modifica wikitesto]Le proprietà dell'RNA rendono concettualmente possibile la presenza di un mondo a RNA, sebbene la sua plausibilità come spiegazione dell'origine della vita sia ancora dibattuta.
Una versione lievemente diversa dell'ipotesi è che un tipo differente di acido nucleico, denominato pre-RNA, sia stato il primo ad apparire come molecola in grado di auto-replicarsi, per poi essere successivamente sostituito dall'RNA. Questi tipi di acido nucleico sono talvolta più facilmente prodotti e/o polimerizzati in condizioni prebiotiche. Suggerimenti per questi tipi di acido nucleico includono il PNA, il TNA o il GNA.[11][12]
Argomenti a favore
[modifica | modifica wikitesto]L'ipotesi è ritenuta estremamente verosimile a causa dell'enorme versatilità della molecola di RNA, in grado di conservare, trasmettere e duplicare l'informazione genetica in modo analogo al DNA ma anche, in aggiunta, di agire come ribozima (grazie alla sua capacità di assumere complesse strutture terziarie) e quindi di catalizzare reazioni come fanno gli enzimi proteici.
La prova più convincente è che nel ribosoma, che è composto di proteine e di RNA ribosomiale, la sintesi proteica è operata non da enzimi (che sono proteine) ma dall'RNA ribosomiale, che in questo caso si comporta come un ribozima. Una seconda prova importante è il fenomeno dello splicing, dove una molecola di pre-RNA è capace di tagliarsi da sola senza l'intervento di enzimi, sebbene l'intervento delle proteine sia, come per il ribosoma, quello di stabilizzare e da agire come "impalcatura" per il complesso di reazione. Ancora, i viroidi, le più semplici entità autoreplicantesi, sono costituiti da RNA che agisce da ribozima. L'RNA inoltre costituisce l'unico materiale genetico di alcuni virus come i retrovirus, prova che l'RNA da solo possa costituire un genoma.
Queste e altre prove presenti negli organismi attualmente viventi supportano fortemente l'idea che l'RNA fosse stata l'ultima molecola autoreplicante prima dell'avvento del DNA[13].
Sebbene i nucleotidi non siano stati individuati nel classico esperimento di Miller-Urey, esistono altri esperimenti, come quello di Joan Oró, che evidenziano la loro possibile sintesi autonoma nelle condizioni ambientali che hanno caratterizzato l'origine della vita. Un successivo esperimento in atmosfera meno riducente di quella di Urey ha prodotto nucleotidi[14], rafforzando ulteriormente l'ipotesi del mondo a RNA.
L'ipotesi è sostenuta anche da studi su ribozimi molto semplici, come gli RNA Q-beta virali, che hanno mostrato capacità autoreplicative anche sotto pressioni selettive molto importanti.[15]
Inoltre le condizioni ambientali della Terra primordiale potrebbero essere state ideali per una molecola labile come l'RNA. I raggi ultravioletti, infatti, inducono contemporaneamente la polimerizzazione dell'RNA e la rottura di altri tipi di molecole organiche potenzialmente in grado di catalizzare la degradazione dell'RNA (come le ribonucleasi). Si tratta in ogni caso di un aspetto ancora non corroborato da osservazioni sperimentali.
Argomenti contrari
[modifica | modifica wikitesto]Le argomentazioni contrarie all'ipotesi si basano sull'improbabilità della formazione spontanea di molecole di RNA, avvalorata anche dal fatto che la base citosina non sia stata sufficientemente testata in metodi di sperimentazione prebiotica, dal momento che essa va facilmente incontro a idrolisi.
Le condizioni prebiotiche necessarie alla formazione spontanea dei tre elementi che costituiscono un nucleotide sono diverse tra loro. Le basi azotate si formano in ambienti differenti rispetto a quelli necessari alla formazione degli zuccheri presenti nello scheletro dell'acido nucleico. Per tale motivo, sarebbe dunque necessario ipotizzare una sintesi spontanea delle due classi di molecole in ambienti separati, seguiti da una successiva unione. Va però detto che, in ambiente acquoso, tale unione è poco probabile, poiché basi azotate e zuccheri non sono comunque in grado di reagire. In ambiente anidro le purine sono in grado di legare gli zuccheri (ma solo l'8% presso il corretto carbonio), mentre tra pirimidine e ribosio non vi è possibilità di legame spontaneo nemmeno in un ambiente non acquoso.
Il terzo elemento, il fosfato, è di per sé estremamente raro nelle soluzioni naturali, poiché precipita velocemente. E anche una volta presente, esso dovrebbe combinarsi con il nucleoside presso il corretto ossidrile. Per potersi inserire in una molecola di RNA, poi, il nucleotide dovrebbe attivarsi attraverso il legame di altri due gruppi fosfato (a formare ad esempio l'adenosintrifosfato). Oltre a tutto ciò, il ribosio deve avere la corretta stereoisomeria, poiché nucleotidi aventi chiralità errata agiscono come terminatori di trascrizione.[16]
Anche sulla base di considerazioni di questo tipo, Cairns-Smith criticò nel 1982 gli esponenti della comunità scientifica per avere esagerato nel trarre conseguenze dall'esperimento di Miller-Urey. Egli sostenne infatti che tale esperimento non avesse dimostrato che gli acidi nucleici fossero alla base dell'origine della vita, ma semplicemente che questa ipotesi non fosse implausibile. Cairns-Smith argomentò che, per raggiungere quantità di molecole necessarie per dare origine alla vita, il processo di costruzione degli acidi nucleici avrebbe dovuto rispettare 18 condizioni autonome tra loro per diversi milioni di anni.
Il mondo a RNA nel dettaglio
[modifica | modifica wikitesto]Meccanismi di sintesi prebiotica dell'RNA
[modifica | modifica wikitesto]L'ipotesi presuppone la presenza nel brodo primordiale di nucleotidi in grado di formare facilmente legami chimici tra loro e di rompere tali legami con la stessa probabilità, grazie alla bassa energia richiesta per tali eventi. In questo ambiente, alcune sequenze di basi aventi proprietà catalitiche sarebbero state in grado di amplificare la formazione di sequenze con identiche caratteristiche, grazie proprio all'attività catalitica in grado di ridurre l'energia necessaria alla formazione di tali sequenze. La formazione di tali sequenze avrebbe avuto come principale conseguenza il fatto che la produzione di filamenti di RNA divenisse decisamente più vantaggiosa della rottura degli stessi.
Queste sequenze sono ritenute essere le prime, primitive forme di vita. In un mondo a RNA, la selezione naturale avrebbe avuto come obiettivo proprio le sequenze di RNA in competizione tra loro. Solo le più efficienti in termini di catalisi e auto-riproduzione sarebbero sopravvissute fino a evolversi e formare il moderno RNA.
La competizione tra RNA potrebbe aver favorito l'emergere di cooperazione tra diverse catene, spianando così la strada alle prime proto-cellule. All'interno di questo set di RNA, alcuni potrebbero avere sviluppato la capacità di catalizzare la formazione di un legame peptidico con la conseguenza, evolutivamente vantaggiosa, di poter generare peptidi accessori per le attività catalitiche dei ribozimi. Allo stesso modo potrebbero esser stati reclutati nel processo di formazione della vita anche tutte le altre molecole chimiche che oggi la caratterizzano, come il DNA, i lipidi o i carboidrati.
Altre ipotesi
[modifica | modifica wikitesto]Esiste una differente versione dell'ipotesi, denominata ipotesi del mondo a pre-RNA. Secondo tale teoria, sarebbe esistito un altro acido nucleico prima dell'RNA. Tra quelli proposti, figura soprattutto il PNA, più stabile dell'RNA e di più facile sintesi nelle condizioni prebiotiche (nelle quali la formazione di ribosio e l'aggiunta dei gruppi fosfato, entrambi assenti nel PNA, è decisamente problematica). Anche il TNA e il GNA sono stati proposti come acidi nucleici pre-RNA.
Un'ulteriore teoria, in parte complementare, è quella dell'ipotesi del mondo a IPA (idrocarburi policiclici aromatici).
Patrick Forterre ha ipotizzato che i virus potrebbero essere stati degli strumenti necessari per la transizione da RNA a DNA degli Eubacteria, Archaea ed Eukaryota. Egli ha proposto che l'ultimo antenato comune tra i tre domini possa essere stato un virus a RNA. Alcuni virus avrebbero in seguito adottato il DNA, molto meno soggetto a danni esterni, iniziando a infettare gli organismi appartenenti ai tre domini con tale acido nucleico, permettendo così anche la loro evoluzione.[17]
Implicazioni correlate al mondo a RNA
[modifica | modifica wikitesto]L'ipotesi del mondo a RNA, se vera, ha importanti conseguenze correlate alla stessa definizione di vita. Per la maggior parte del ventesimo secolo, la comunità scientifica ha considerato la vita alla stregua di una combinazione di DNA e proteine, considerate le due macromolecole dominanti, relegando l'RNA allo status di semplice molecola accessoria.
Questa ipotesi pone invece l'RNA al centro dell'origine della vita. Ciò è suggerito da numerosi studi che, negli ultimi dieci anni, hanno rivalutato il ruolo dell'RNA, scoprendone funzioni precedentemente non note ed evidenziandone il ruolo critico nel funzionamento della vita. Nel 2001 sono state risolte le strutture tridimensionali dei ribosomi, mettendo in evidenza che, come già detto, il sito catalitico è composto da ribozimi (RNA) e non da enzimi (proteine) come precedentemente ipotizzato.
Ulteriori scoperte in questo senso, che valorizzano il ruolo fisiologico fondamentale dell'RNA sono state quelle relative al ruolo delle small nuclear ribonucleoproteins (snRNPs) nel processamento del pre-mRNA, nell'editing dell'RNA, nella trascrizione inversa e nel mantenimento dei telomeri.
Note
[modifica | modifica wikitesto]- ^ Walter Gilbert, The RNA World, in Nature, vol. 319, febbraio 1986, p. 618, DOI:10.1038/319618a0.
- ^ Carl Woese, The Genetic Code, Harper & Row, gennaio 1968, ISBN 978-0-06-047176-7.
- ^ Alexander Rich, On the problems of evolution and biochemical information transfer., in Horizons In Biochemistry, 1962, pp. 103–126.
- ^ W. K. Johnston, P. J. Unrau, M. S. Lawrence, M. E. Glasner and D. P. Bartel RNA-Catalyzed RNA Polymerization: Accurate and General RNA-Templated Primer Extension. Science 292, 1319 (2001)
- ^ Huang, Yang, and Yarus, RNA enzymes with two small-molecule substrates Archiviato il 3 luglio 2012 in Archive.is.. Chemistry & Biology, Vol 5, 669-678, November 1998
- ^ Unrau, P.J. and Bartel, D.P. (1998) RNA-catalysed nucleotide synthesis. Nature 395, 260-263
- ^ Zhang and Cech, Peptide bond formation by in vitro selected ribozymes. Nature 390, 96-100
- ^ Szathmary E., The origin of the genetic code: amino acids as cofactors in an RNA world Archiviato il 2 ottobre 2007 in Internet Archive.. Trends in Genetics, Volume 15, Number 6, 1 June 1999 , pp. 223-229(7)
- ^ T Lindahl, Instability and decay of the primary structure of DNA, in Nature, vol. 362, n. 6422, aprile 1993, pp. 709-15, PMID 8469282.
- ^ S Pääbo, Ancient DNA, in Scientific American, vol. 269, n. 5, novembre 1993, pp. 60-66.
- ^ Leslie Orgel, A Simpler Nucleic Acid, in Science, vol. 290, n. 5495, novembre 2000, pp. 1306-7, DOI:10.1126/science.290.5495.1306.
- ^ Nelson, K.E., Levy, M.; Miller, S.L., Peptide nucleic acids rather than RNA may have been the first genetic molecule, in Proc. Natl. Acad. Sci. USA, vol. 97, n. 8, aprile 2000, pp. 3868–71, PMID 10760258.
- ^ Life from an RNA World. The Ancestor Within, Harvard University press.
- ^ Formation of nucleobases in a Miller–Urey reducing atmosphere, su ncbi.nlm.nih.gov.
- ^ The Basics of Selection (London: Springer, 1997)
- ^ Joyce GF, Visser GM, van Boeckel CA, van Boom JH, Orgel LE, van Westrenen J., Chiral selection in poly(C)-directed synthesis of oligo(G), in Nature, vol. 310, n. 5978, agosto 1984, pp. 602-4, PMID 6462250.
- ^ Zimmer C., Did DNA come from viruses?, in Science, vol. 312, n. 5775, 2006, pp. 870-2, PMID 16690855.
Bibliografia
[modifica | modifica wikitesto]- A. G. Cairns-Smith, Genetic Takeover: And the Mineral Origins of Life, Cambridge University Press, 1993, ISBN 0-521-23312-7.
- L. E. Orgel, The origin of life on the Earth, in Scientific American, vol. 271, Oct 1994, pp. 76-83.
- Adrian Woolfson, Life Without Genes, London, Flamingo, Sep 2000, ISBN 978-0-00-654874-4.
- Alexander V. Vlassov, The RNA World on Ice: A New Scenario for the Emergence of RNA Information, in Journal of Molecular Evolution, vol. 61, Jul 2005, pp. 264-273.
- Matthew W. Powner, Béatrice Gerland, John D. Sutherland, Synthesis of activated pyrimidine ribonucleotides in prebiotically plausible conditions, Nature 459, 239-242 (14 May 2009) doi:10.1038.
- Jack W. Szosta, Origins of life: Systems chemistry on early Earth, Nature 459, 171-172 (14 May 2009) doi:10.1038.
Voci correlate
[modifica | modifica wikitesto]Collegamenti esterni
[modifica | modifica wikitesto]- (EN) "The RNA world", articolo del premio Nobel Sidney Altman (2001).
- (EN) "Exploring the new RNA world", articolo del premio Nobel Thomas Cech (2004).
- Origine della vita, c’è un nuovo ingrediente per il brodo primordiale, su galileonet.it, 6 dicembre 2018.

