Poro nuclear
Os poros nucleares son orificios ocupados por grandes complexos proteicos (complexo do poro nuclear) que atravesan a envoltura nuclear, comunicando o núcleo celular co citoplasma da célula eucariota. Como media na envoltura nuclear dunha célula de vertebrado hai uns 2000 complexos dos poros nucleares, pero isto varía dependendo do tipo celular e o estado do ciclo celular no que se atope. As proteínas que forman parte do complexo do poro nuclear denomínanse nucleoporinas. Aproximadamente a metade das nucleoporinas conteñen un pregamento en solenoide alfa (formado por varias hélices alfa enroladas circularmente) ou un pregamento de tipo propulsor beta (ou β-propeller, formado por varias láminas beta en disposición toroidal), ou ben, noutros casos, teñen ambos en dominios estruturais separados. A outra metade das nucleoporinas presenta características estruturais típicas de proteínas "non pregadas en estado nativo", é dicir, son proteínas moi flexibles que no seu estado normal carecen dunha estrutura secundaria ordenada.[1] Estas proteínas de estrutura desordenada son as nucleoporinas FG, así chamadas porque a súa secuencia de aminoácidos contén moitas repeticións do dipéptido fenilalanina—glicina.[2]
Os complexos dos poros nucleares permiten o transporte de moléculas solubles en auga a través da envoltura nuclear. Este transporte inclúe o paso de ARN e subunidades dos ribosomas desde o núcleo ao citoplasma, e de proteínas (como as ADN polimerases, laminas ou histonas), carbohidratos, moléculas de sinalización e lípidos desde o citoplasma cara ao núcleo. É de salientar que o complexo do poro nuclear pode canalizar activamente 1000 translocacións de substancias por complexo e por segundo. Aínda que as moléculas pequenas poden simplemente difundir a través dos poros, as moléculas grandes poden ter secuencias sinalizadoras específicas de recoñecemento e difundir cara ao interior ou exterior do núcleo coa axuda das nucleoporinas. Isto coñécese como o ciclo Ran. Cada unha das oito subunidades proteicas que rodean o poro real (o anel externo) proxecta unha proteína con forma radial na canle do poro. O centro do poro con frecuencia parece que contén unha estrutura a modo de tapón. Aínda non se sabe se esta estrutura é un verdadeiro tapón ou se se trata simplemente dunha molécula transportada que está atravesando o poro.
Tamaño e complexidade
[editar | editar a fonte]O complexo do poro nuclear completo ten un diámetro duns 120 nanómetros en vertebrados.[3] O diámetro da canle interna é menor, duns 9 nm,[4] pero varía segundo as especies, cunha profundidade, tamén variable, duns 45 nm.[5][6] Como comparación, as moléculas de ARNm, que son monocatenarias e teñen que atravesar os poros, teñen un grosor duns 0,5 a 1 nm.[7] A masa molecular dos complexos do poro de mamíferos é duns 124 megadaltons e comprende aproximadamente 30 compoñentes proteicos diferentes, cada un con múltiples copias.[8] No lévedo Saccharomyces cerevisiae é menor, e pesa só 66MDa.[9]
Transporte a través do complexo do poro nuclear
[editar | editar a fonte]
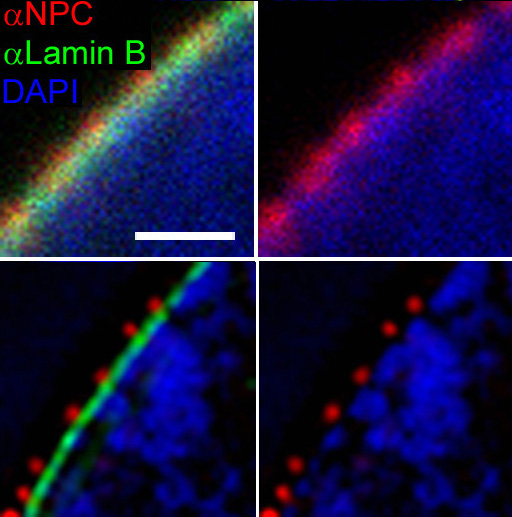
As pequenas partículas (<~40kDa) poden pasar a través do complexo do poro nuclear por difusión pasiva. Partículas meirandes tamén poden pasar a través do poro pero en cantidades case desprezables.[10][11] Para un transporte eficiente a través do poro requírense varios factores proteicos.[12] As carioferinas, as cales poden funcionar como importinas ou exportinas son parte da superfamilia de proteínas Importinas-β, na que todas comparten unha estrutura tridimensional semellante.
Importación de proteínas
[editar | editar a fonte]Calquera molécula que leve un sinal de localización nuclear exposto será transportada rápida e eficientemente a través do poro. Coñécense varias secuencias sinal de localización nuclear, e xeralmente conteñen unha secuencia polipeptídica conservada con residuos básicos como Pro-Lys-Lys-Lys-Arg-Lys-Val (ou PKKKRKV). Calquera substancia que leve un destes sinais de localización nuclear será recollida por importinas e levada ao núcleo.
O esquema clásico da importación de proteínas con sinais de localización nuclear comeza coa unión da importina-α á secuencia do sinal de localización nuclear, e actúa como unha ponte para que se una a importina-β. O complexo importina-β—importina-α—molécula transportada é entón dirixido cara ao poro e difunde a través del. Unha vez que o complexo está no núcleo, a RanGTP únese á importina-β e sepáraa do complexo. Entón a proteína de susceptibilidade da apoptose celular (CAS), que é unha exportina que está ligada no núcleo á RanGTP, separa a importina-α da molécula transportada. A proteína con sinal de localización nuclear queda así libre no nucleoplasma. A importinaβ-RanGTP e o complexo importinaα-CAS-RanGTP difunde de volta ao citoplasma, onde a hidrólise do GTP a GDP provoca a liberación das importinaβ e α, as cales quedan dispostas para a importación dunha nova proteína con sinal de localización nuclear.
Aínda que a molécula transportada pasa a través do poro coa axuda de proteínas chaperonas, a translocación a través do poro non depende do gasto de enerxía. Porén, para completar o ciclo de importación precísase a hidrólise de 2 GTPs e é, por tanto, dependente dun gasto enerxético, polo que debe considerarse como un transporte activo. O ciclo de importación está alimentado polo gradiente núcleo-citoplásmico de RanGTP. Este gradiente xordee da localización exclusiva dos RanGEFs no núcleo, que son proteínas que intercambian GDP a GTP nas moléculas Ran. Así hai unha elevada concentración de RanGTP no núcleo comparado co citoplasma.
Exportación de proteínas
[editar | editar a fonte]Algunhas proteínas nucleares deben exportarse do núcleo ao citoplasma, igual que as subunidades dos ribosomas e os ARN mensaxeiros. Para isto existe un mecanismo de exportación similar ao de importación.
No esquema clásico da exportación, as proteínas cunha secuencia nuclear de exportación poden unirse no núcleo para formar un complexo heterotrimérico cunha exportina e a RanGTP (por exemplo a exportina CRM1). O complexo pode entón difundir ao citoplasma onde se hidroliza GTP e a proteína con secuencia nuclear de exportación é lberada. O CRM1-RanGDP difunde de volta ao núcleo onde as RanGEFs fan o intercambio de GDP a GTP. Este proceso é tamén dependente do gasto de enerxía, xa que consome un GTP. A exportación coa exportina CRM1 pode ser inhibida pola leptomicina B.
Exportación de ARN
[editar | editar a fonte]Existen diferentes vías de exportación a través dos poros nucleares para cada tipo de ARN. A exportación de ARN tamén está mediada por secuencias de exportación nuclear; estas secuencias están en proteínas de unión ao ARN (agás no caso do ARNt, que non ten ningún adaptador). É salientable que, excepto o ARNm, todos os ARN virais e ARN celulares (ARNt, ARNr, snRNA, microARN) son dependentes da RanGTP. Para a exportación do núcleo do ARNm son necesarios factores de exportación. Son factores de exportación o Mex67/Tap (subunidade maior) e o Mtr2/p15 (subunidade menor).
Ensamblaxe do complexo dos poros nucleares
[editar | editar a fonte]Como os complexos dos poros controlan o acceso ao xenoma, é esencial que existan en grandes cantidades en momentos do ciclo celular nos que se necesita moita transcrición. Por exemplo, certos mamíferos e as células de lévedos dobran a cantidade de complexos dos poros no núcleo entre as fases G1 e G2 do ciclo celular. E os ovocitos acumulan un gran número de complexos dos poros para prepararse para as rápidas mitoses que se producen nas fases iniciais do desenvolvemento. As células en interfase deben tamén acrecentar o nivel de xeración de complexos dos poros para manter constantes os niveis dos mesmos na célula, xa que algúns poden estragarse. Algunhas células poden mesmo incrementar o número de complexos dos poros debido a un aumento da demanda de transcrición.[13]
Teorías de ensamblaxe
[editar | editar a fonte]Hai varias teorías que tratan de explicar como se ensamblan estes grandes complexos proteicos. Dado que a immunodepleción de certos complexos proteicos, como o complexo Nup 107–160, leva á formación de núcleos sen poros, é probable que os complexos Nup estean implicados na fusión da membrana externa da envoltura nuclear coa interna, en lugar de que sexa a fusión previa da membrana (con unión posterior dos complexos) a que comece a formación do poro. Hai varias maneiras de que isto conduza á formación dun complexo do poro completo.
- Unha posibilidade é que o Nup se una á cromatina como un complexo proteico. Despois insírese na dobre membrana da envoltura nuclear preto da cromatina. Isto, á súa vez, orixina a fusión desas membranas, creando o poro. Arredor deste complexo proteico inicial uniríanse outros formando o complexo do poro completo. Este método é posible durante cada fase da mitose. Células post mitóticas poderían formar a membrana primeiro e os poros serían inseridos despois.
- Outro modelo para explicar a formación dos complexos dos poros é a produción dun complexo preporo inicial, en lugar dun único complexo proteínico completo. Este preporo formaríase cando se unisen varios complexos Nup e se ligasen á cromatina. Isto faría que se formase unha dobre membrana arredor do complexo do preporo durante a rexeneración da envoltura nuclear ao final da mitose. Usando microscopía electrónica observáronse posibles estruturas de preporo na cromatina antes da formación da envoltura nuclear.[14] Durante a interfase do ciclo celular a formación do complexo do preporo tería lugar dentro do núcleo, e cada compoñente sería transportado ao núcleo atravesando complexos do poro xa existentes. Os Nups necesarios, unha vez formados no citoplasma, uniríanse a unha importina, o que impediría a ensamblaxe do preporo no citoplasma. Unha vez que xa foron tansportados dentro do núcleo, as RanGTP uniríanse á importina e causarían a liberación do material transportado. Agora as Nup estarían libres no núcleo para formar o preporo. Hai evidencias da unión de importinas para traer ao núcleo as nucleoporinas Nup 107 e Nup 153.[13] A ensamblaxe dos complexos dos poros é un proceso moi rápido, aínda que hai estadios intermedios, o que indica que esta ensamblaxe é un proceso que progresa paso a paso.[15]
- Un terceiro posible método de ensamblaxe dos complexos dos poros é a división doutro xa existente. Este método parece perfecto para a formación dos complexos dos poros durante a interfase. Sucede cando se engaden máis protómeros a un complexo do poro xa existente. Parece que a simetría de oito pregamentos do complexo do poro ten un grao de plasticidade que permitiría isto. Despois engadiríanse protómeros dabondo como para permitir que se escindise o complexo do poro orixinal, formando outro novo. Este método de ensamblaxe do complexo do poro pode suceder soamente durante a interfase do ciclo celular.
Desensamblaxe
[editar | editar a fonte]Durante a mitose os complexos dos poros nucleares parecen desensamblarse por fases. Algunhas nucleoporinas periféricas como a Nup 153, Nup 98 e Nup 214 disgréganse do complexo do poro. O resto, que poden ser consideradas proteínas de armazón, permanecen estables en forma de complexos anulares cilíndricos na envoltura nuclear. Esta desensamblaxe dos grupos periféricos dos complexos dos poros moi probablemente está impulsada polo fosfato, xa que varias destas nucleoporinas son fosforiladas durante as fases da mitose. Porén, non se coñecen os encimas implicados na fosforilación in vivo. En metazoos (os cales presentan mitose aberta) a envoltura nuclear degrádase rapidamente despois da perda das Nups periféricas. A razón disto pode ser o cambio na arquitectura dos complexos dos poros. Este cambio pode facer que o complexo do poro sexa máis permeable aos encimas que interveñen na degradación da envoltura nuclear, como a tubulina citoplasmática, e tamén permitir a entrada de proteínas reguladoras mitóticas chave.
Preservación da integridade do núcleo en certos fungos
[editar | editar a fonte]Observouse en fungos que experimentan mitoses pechadas (nas cales o núcleo non desaparece), que o cambio da barreira de permeabilidade da envoltura nuclear debíase a cambios nos complexos dos poros nucleares e isto era o que permitía a entrada de reguladores mitóticos. En Aspergillus nidulans o cambio na composición dos complexos dos poros parece efectuala a quinase mitótica NIMA, posiblemente por fosforilación das nucleoporinas Nup98 e Gle2/Rae1. Esta remodelación parece que permite aos complexos de proteínas cdc2/cyclinB entrar no núcleo xunto con moitas outras proteínas como as tubulinas solubles. O armazón dos complexos dos poros queda intacto durante toda a mitose pechada. Isto parece preservar a integridade da envoltura nuclear.
Notas
[editar | editar a fonte]- ↑ Denning D, Patel S, Uversky V, Fink A, Rexach M (2003). "Disorder in the nuclear pore complex: the FG repeat regions of nucleoporins are natively unfolded". Proc Natl Acad Sci USA 100 (5): 2450–5. PMC 151361. PMID 12604785. doi:10.1073/pnas.0437902100.
- ↑ Peters R (2006). "Introduction to nucleocytoplasmic transport: molecules and mechanisms". Methods Mol Biol 322: 235–58. PMID 16739728. doi:10.1007/978-1-59745-000-3_17. Arquivado dende o orixinal o 28 de setembro de 2007. Consultado o 26 de maio de 2011.
- ↑ Winey, Mark; Yarar, Defne; Giddings, Jr., Thomas H; Mastronarde, David N (1 November 1997). "Nuclear Pore Complex Number and Distribution throughout the Saccharomyces cerevisiae Cell Cycle by Three-Dimensional Reconstruction from Electron Micrographs of Nuclear Envelopes". Molecular Biology of the Cell 8 (11): 2119–2132. doi:10.1091/mbc.8.11.2119. Consultado o 12 December 2014.
- ↑ Bruce Alberts, Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, and Peter Walter. Molecular Biology of the Cell. 4th edition. (2001)(na Libraría en liña do NCBI). Capítulo: The Transport of Molecules between the Nucleus and the Cytosol. New York: Garland Science; 2002. ISBN 0-8153-3218-1ISBN 0-8153-4072-9
- ↑ Keminer, Oliver; Peters, Reiner (July 1999). "Permeability of Single Nuclear Pores". Biophysical Journal 77 (1): 217–228. doi:10.1016/S0006-3495(99)76883-9. Consultado o 12 December 2014.
- ↑ Mohr, Dagmar; Frey, Steffen; Fischer, Torsten; Güttler, Thomas; Görlich, Dirk (13 August 2009). "Characterisation of the passive permeability barrier of nuclear pore complexes". The EMBO Journal 28 (17): 2541–2553. doi:10.1038/emboj.2009.200. Consultado o 12 December 2014.
- ↑ Kuznetsov, Yurii G.; Daijogo, Sarah; Zhou, Jiashu; Semler, Bert L.; McPherson, A. (March 2005). "Atomic Force Microscopy Analysis of Icosahedral Virus RNA". Journal of Molecular Biology 347 (1): 41–52. doi:10.1016/j.jmb.2005.01.006. Consultado o 12 December 2014.
- ↑ Alber F, Dokudovskaya S, Veenhoff LM; et al. (2007). "Determining the architectures of macromolecular assemblies". Nature 450 (7170): 683–94. PMID 18046405. doi:10.1038/nature06404.
- ↑ Rout MP, Blobel G (1993). "Isolation of the yeast nuclear pore complex". J. Cell Biol. 123 (4): 771–83. PMC 2200146. PMID 8227139.
- ↑ Rodriguez M, Dargemont C, Stutz F (2004). "Nuclear export of RNA". Biol Cell 96 (8): 639–55. PMID 15519698. doi:10.1016/j.biolcel.2004.04.014.
- ↑ Marfori M, Mynott A, Ellis JJ; et al. (2010). "Molecular basis for specificity of nuclear import and prediction of nuclear localization". Biochimica Et Biophysica Acta. PMID 20977914. doi:10.1016/j.bbamcr.2010.10.013.
- ↑ Reed R, Hurt E (2002). "A conserved mRNA export machinery coupled to pre-mRNA splicing". Cell 108 (4): 523–31. PMID 11909523. doi:10.1016/S0092-8674(02)00627-X.
- ↑ 13,0 13,1 Rabut G, Lénárt P, Ellenberg J (2004). "Dynamics of nuclear pore complex organization through the cell cycle". Current opinion in cell biology 16 (3): 314–21. PMID 15145357. doi:10.1016/j.ceb.2004.04.001.
- ↑ Sheehan MA, Mills AD, Sleeman AM, Laskey RA, Blow JJ (1988). "Steps in the assembly of replication-competent nuclei in a cell-free system from Xenopus eggs". The Journal of cell biology 106 (1): 1–12. PMC 2114961. PMID 3339085. doi:10.1083/jcb.106.1.1.
- ↑ Kiseleva E, Rutherford S, Cotter LM, Allen TD, Goldberg MW (2001). "Steps of nuclear pore complex disassembly and reassembly during mitosis in early Drosophila embryos". Journal of cell science 114 (Pt 20): 3607–18. PMID 11707513. Arquivado dende o orixinal o 13 de setembro de 2019. Consultado o 26 de maio de 2011.
Véxase tamén
[editar | editar a fonte]Outros artigos
[editar | editar a fonte]Ligazóns externas
[editar | editar a fonte]- BUHistology - 20104loa [1]
- Nuclear Pore Complex animationsArquivado 07 de febreiro de 2009 en Wayback Machine.
- Nuclear Pore Complex illustrationsArquivado 07 de febreiro de 2009 en Wayback Machine.