Dobre hélice

En bioloxía molecular, o termo dobre hélice[1] refírese á estrutura formada por moléculas de ácidos nuicleicos bicatenarias como o ADN. A estrutura en dobre hélice dun complexo de ácido nucleico orixínase como consecuencia da súa estrutura secundaria, e é un compoñente fundamental para determinar a súa estrutura terciaria. James Watson publicou en 1968 o libro The Double Helix: A Personal Account of the Discovery of the Structure of DNA explicando ao gran público o descubrimento da estrutura da dobre hélice do ADN.
O polímero da dobre hélice do ADN mantense unido por medio de enlaces entre as bases nitroxenadas dos nucleótidos complementarias.[2] No ADN B, que é a estrutura máis común da dobre hélice, descrita por Watson e Crick, esta é dextroxira e ten uns 10–10,5 nucleótidos por volta.[3] A estrutura de dobre hélice do ADN forma un suco maior (máis profundo ou amplo) e un suco menor.[2] Dada a diferenza de largura entre os dous tipos sucos, moitas proteínas que se unen ao ADN fano no suco maior.[4]
Historia
[editar | editar a fonte]O modelo de dobre hélice da estrutura do ADN publicárono por primeira vez na revista Nature James D. Watson e Francis Crick en 1953,[5] (e as coordenadas X, Y, Z en 1954[6]) baseándose nunha imaxe de difracción de raios X crucial do ADN etiquetada como "Foto 51", obtida polo equipo de Rosalind Franklin en 1952,[7] seguida dunha imaxe do ADN máis clara obtida xunto con Raymond Gosling,[8][9] Maurice Wilkins, Alexander Stokes, e Herbert Wilson,[10] xunto con información bioquímica e de apareamento de bases de Erwin Chargaff.[11][12][13][14][15][16] O modelo previo era un ADN de tripla febra.[17]
A determinación de que a estrutura do ADN era unha dobre hélice serviu para dilucidar o mecanismo de apareamento de bases polo cal a información xenética se almacena e copia nos organismos vivos e é considerado como un dos descubrimentos científicos máis importantes do século XX. Crick, Wilkins, e Watson compartiron o premio Nobel de Medicina de 1962 polas súas contribucións a este descubrimento.[18] (Franklin, cuxos datos de difracción de raios X foran fundamentais, morrera en 1958, polo que non se lle puido dar o premio Nobel, que só se adoitaba dar a científicos vivos.)
Hibridación de ácidos nucleicos
[editar | editar a fonte]- Artigo principal: Termodinámica dos ácidos nucleicos.
A hibridación é o proceso de unión por apareamento de bases complementario de dúas moléculas de ácido nucleico para formar a dobre hélice. Denomínase "fusión" ao proceso polo cal a dobre hélice se separa en dúas febras independentes ao romperen as interaccións entre as febras (fundamentalmente enlaces de hidróxeno). Estes enlaces son febles e as febras poden separarse facilmente aplicando calor suave, encimas, ou forza física. A fusión ocorre preferentemente en certos puntos do ácido nucleico.[19] As secuencias ricas en T e A funden máis facilmente que as rexións ricas en C e G.[20] Estas características mecánicas aprovéitaas a célula para usar secuencias como TATA ao inicio de moitos xenes para axudar á ARN polimerase a separar as febras do ADN para facer a transcrición.
A separación das febras por quentamento suave, como o utilizado na PCR, é simple con tal que as moléculas teñan menos de 10.000 pares de bases (10 pares de quilobases ou 10 kbp). O entrelazado das febras de ADN fai que os segmentos longos sexan difíciles de separar. A célula evita este problema permitindo que os seus encimas separen (fundan) o ADN (helicases) traballen en conxunción coas topoisomerases, que poden cortar quimicamente a cadea de enlaces fosfodiéster dunha das febras para que poida rotar arredor da outra. As helicases desenrolan as febras para facilitar o avance dos encimas que len o ADN como as ADN polimerases.
Xeometría dos pares de bases
[editar | editar a fonte]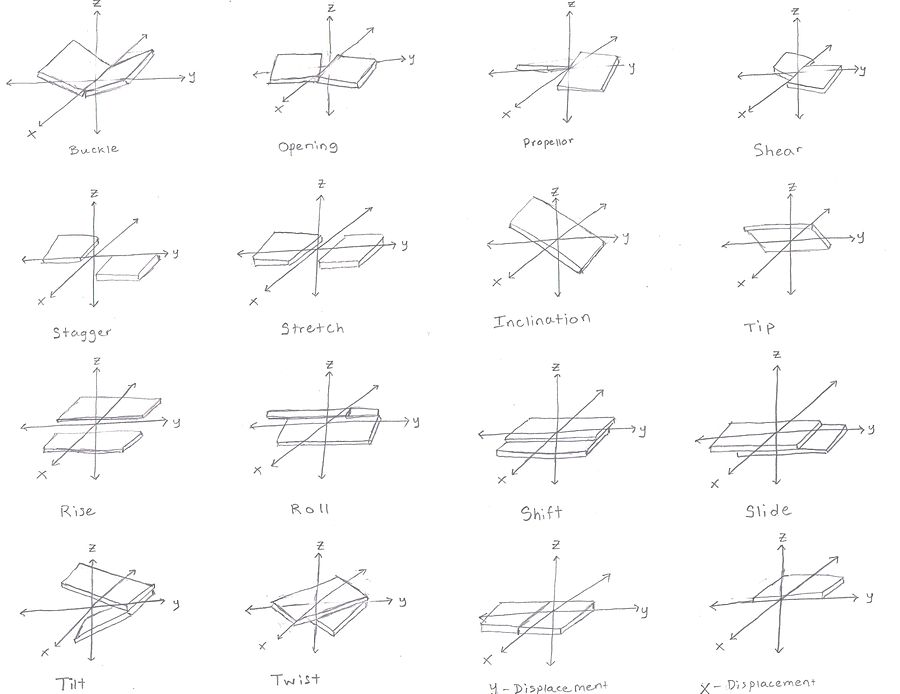
Para cada par de bases, considerado en relación co seu predecesor, hai que considerar as xeometrías dos pares de bases indicadas na figura, que se caracterizan por 6 coordenadas.[21][22][23]
Estes valores definen de forma precisa a localización e orientación no espazo de cada base ou par de bases nunha molécula de ácido nucleico en relación coa súa predecesora ao longo do eixe da hélice. Xuntos, caracterizan a estrutura helicoidal da molécula. En rexións do ADN ou ARN onde a estrutura "normal" está distorsionada, o cambio nestes valores pode utilizarse para describir dita distorsión.
Xeometrías da hélice
[editar | editar a fonte]Crese que na natureza existen polo menos tres conformacións do ADN, chamadas ADN A, ADN B, e ADN Z. A forma "B" é a descrita por James D. Watson e Francis Crick e é a que predomina nas células.[24] A súa hélice ten 23,7 Å de diámetro e unha lonxitude de 34 Å por cada secuencia de 10 pares de bases. A dobre hélice dá un xiro completo sobre o seu eixe cada 10,4-10,5 pares de bases en solución. Esta frecuencia de xiro (paso de rosca) depende en gran medida das forzas que cada base exerce sobre as súas veciñas da cadea.
O ADN A e o Z teñen diferenzas significativas na súa xeometría e dimensións con respecto ao ADN B, pero sempre forman estruturas helicoidais. A forma A probablemente aparece só en mostras deshidratadas de ADN, como as utilizadas en experimentos de cristalografía, e posiblemente en moléculas híbridas formadas por apareamento de febras de ADN e ARN. Os segmentos de ADN que as células metilaron con propósitos reguladores poden adoptar a xeometría Z, na cal as febras xiran arredor do eixe da hélice de forma oposta a como o fan no ADN A e ADN B, é dicir, o xiro é levoxiro. Hai tamén probas de que complexos proteína-ADN forman estruturas de tipo ADN Z.
Ademais dos ADN A, B e Z son posibles outras conformacións; entre as descritas ata agora están tamén as do ADN C, ADN E,[25] L-ADN (forma enantiomérica do D-ADN),[26] ADN P,[27] ADN S etc.[28] De feito, só as letras F, Q, U, V, e Y quedaban dispoñibles en 2011 para describir as novas estruturas do ADN que puidesen descubrirse no futuro.[29][30] Algunhas destas formas foron creadas sinteticamente. Hai tamén formas de ADN de tripla hélice e formas cuádruplex como o cuádruplex G.


| Atributos de xeometría | ADN A | ADN B | ADN Z |
|---|---|---|---|
| Sentido da hélice | dextroxira | dextroxira | levoxira |
| Unidade repetida | 1 par de bases | 1 par de bases | 2 pares de bases |
| Rotación por par de bases | 32,7° | 34,3° | 60°/2 |
| Pares de bases por xiro | 11 | 10,5 | 12 |
| Inclinación dos pares de bases con respecto ao eixe | +19° | −1,2° | −9° |
| Elevación (rise) por par de bases ao longo do eixe | 2,3 Å (0.23 nm) | 3,32 Å (0,332 nm) | 3,8 Å (0,38 nm) |
| Xiro da hélice | 28,2 Å (2,82 nm) | 33,2 Å (3,32 nm) | 45,6 Å (4,56 nm) |
| Torsión media da hélice (mean propeller twist) | +18° | +16° | 0° |
| Ángulo glicosilo (orientación dos substituíntes das bases con respecto dos residuos de azucres) | anti | anti | C: anti, G: sin |
| Pregamento do azucre (ring pucker) | C3'-endo | C2'-endo | C: C2'-endo, G: C2'-exo |
| Diámetro | 23 Å (2,3 nm) | 20 Å (2,0 nm) | 18 Å (1,8 nm) |
| Fontes:[31][32][33] | |||
Sucos do ADN
[editar | editar a fonte]
As dúas febras helicoidais do ADN forman unha serie de espazos entrantes. Pode trazarse outra dobre hélice seguindo estes espazos, ou sucos, que quedan entre as febras. Estes espazos baleiros son adxacentes aos pares de bases e poden proporcionar un sitio de unión para moléculas. Como as febras non están directamente en fronte unha da outra, os sucos son de tamaño desigual. Un dos sucos, chamado suco maior, ten unha largura de 22 Å e o outro, chamado suco menor, ten unha largura de 12 Å.[34] A estreitura que presenta o suco menor significa que as beiras das bases son máis accesibles no suco maior. Como resultado, proteínas como os factores de transcrición que poden unirse a secuencias específicas no ADN de dobre febra, xeralmente fan contacto cos lados das bases expostas no suco maior.[4] Esta situación varía en conformacións infrecuentes do ADN que se atopan na célula, pero os sucos maior e menor sempre se denominan así facendo referencia ás diferenzas de tamaño que se verían se o ADN adoptase a conformación B ordinaria.
Formas non helicoidais
[editar | editar a fonte]Na década de 1970 foron considerados durante breve tempo modelos non helicoidais alternativos do ADN como unha solución potencial a certos problemas observados na replicación do ADN en plásmidos e na cromatina. Porén, estes modelos foron abandonados en favor do modelo en dobre hélice debido aos avances experimentais que houbo como a cristalografía de raios X de dúplex de ADN e posteriormente da partícula central do nucleosoma, e o descubrimento das topoisomerases, de modo que estes modelos non helicoidais non son aceptados actualmente na corrente científica predominante.[35][36]
Os ácidos nucleicos monocatenarios non adoptan unha forma helicoidal.
Notas
[editar | editar a fonte]- ↑ Kabai, Sándor (2007). "Double Helix". The Wolfram Demonstrations Project.
- ↑ 2,0 2,1 Alberts; et al. (1994). The Molecular Biology of the Cell. New York: Garland Science. ISBN 978-0-8153-4105-5.
- ↑ Wang JC (1979). "Helical repeat of DNA in solution". PNAS 76 (1): 200–203. Bibcode:1979PNAS...76..200W. PMC 382905. PMID 284332. doi:10.1073/pnas.76.1.200. Arquivado dende o orixinal o 24 de marzo de 2019. Consultado o 20 de marzo de 2015.
- ↑ 4,0 4,1 Pabo C, Sauer R (1984). "Protein-DNA recognition". Annu Rev Biochem 53: 293–321. PMID 6236744. doi:10.1146/annurev.bi.53.070184.001453.
- ↑ James D. Watson and Francis Crick (1953). "A structure for deoxyribose nucleic acid" (PDF). Nature 171 (4356): 737–738. Bibcode:1953Natur.171..737W. PMID 13054692. doi:10.1038/171737a0.
- ↑ Crick F, Watson JD (1954). "The Complementary Structure of Deoxyribonucleic Acid" (PDF). Proceedings of the Royal Society of London. 223, Series A: 80–96.
- ↑ "Secret of Photo 51". NOVA. PBS.
- ↑ http://www.nature.com/nature/dna50/franklingosling.pdf
- ↑ "The Structure of the DNA Molecule". Arquivado dende o orixinal o 21 de xuño de 2012. Consultado o 20 de marzo de 2015.
- ↑ Wilkins MHF, Stokes AR, Wilson HR (1953). "Molecular Structure of Deoxypentose Nucleic Acids" (PDF). Nature 171 (4356): 738–740. Bibcode:1953Natur.171..738W. PMID 13054693. doi:10.1038/171738a0.
- ↑ Elson D, Chargaff E (1952). "On the deoxyribonucleic acid content of sea urchin gametes". Experientia 8 (4): 143–145. PMID 14945441. doi:10.1007/BF02170221.
- ↑ Chargaff E, Lipshitz R, Green C (1952). "Composition of the deoxypentose nucleic acids of four genera of sea-urchin". J Biol Chem 195 (1): 155–160. PMID 14938364.
- ↑ Chargaff E, Lipshitz R, Green C, Hodes ME (1951). "The composition of the deoxyribonucleic acid of salmon sperm". J Biol Chem 192 (1): 223–230. PMID 14917668.
- ↑ Chargaff E (1951). "Some recent studies on the composition and structure of nucleic acids". J Cell Physiol Suppl 38 (Suppl).
- ↑ Magasanik B, Vischer E, Doniger R, Elson D, Chargaff E (1950). "The separation and estimation of ribonucleotides in minute quantities". J Biol Chem 186 (1): 37–50. PMID 14778802.
- ↑ Chargaff E (1950). "Chemical specificity of nucleic acids and mechanism of their enzymatic degradation". Experientia 6 (6): 201–209. PMID 15421335. doi:10.1007/BF02173653.
- ↑ Pauling L, Corey RB (Feb 1953). "A proposed structure for the nucleic acids". Proc Natl Acad Sci U S A 39 (2): 84–97. Bibcode:1953PNAS...39...84P. PMC 1063734. PMID 16578429. doi:10.1073/pnas.39.2.84.
- ↑ "Nobel Prize - List of All Nobel Laureates".
- ↑ Breslauer KJ, Frank R, Blöcker H, Marky LA (1986). "Predicting DNA duplex stability from the base sequence". PNAS 83 (11): 3746–3750. Bibcode:1986PNAS...83.3746B. PMC 323600. PMID 3459152. doi:10.1073/pnas.83.11.3746.
- ↑ Owczarzy, Richard (2008-08-28). "DNA melting temperature - How to calculate it?". High-throughput DNA biophysics. owczarzy.net. Arquivado dende o orixinal o 30 de abril de 2015. Consultado o 2008-10-02.
- ↑ Dickerson RE (1989). "Definitions and nomenclature of nucleic acid structure components". Nucleic Acids Res 17 (5): 1797–1803. PMC 317523. PMID 2928107. doi:10.1093/nar/17.5.1797.
- ↑ Lu XJ, Olson WK (1999). "Resolving the discrepancies among nucleic acid conformational analyses". J Mol Biol 285 (4): 1563–1575. PMID 9917397. doi:10.1006/jmbi.1998.2390.
- ↑ Olson WK, Bansal M, Burley SK, Dickerson RE, Gerstein M, Harvey SC, Heinemann U, Lu XJ, Neidle S, Shakked Z, Sklenar H, Suzuki M, Tung CS, Westhof E, Wolberger C, Berman HM (2001). "A standard reference frame for the description of nucleic acid base-pair geometry". J Mol Biol 313 (1): 229–237. PMID 11601858. doi:10.1006/jmbi.2001.4987.
- ↑ Richmond; Davey, CA; et al. (2003). "The structure of DNA in the nucleosome core". Nature 423 (6936): 145–150. Bibcode:2003Natur.423..145R. PMID 12736678. doi:10.1038/nature01595.
- ↑ Vargason JM, Eichman BF, Ho PS (2000). "The extended and eccentric E-DNA structure induced by cytosine methylation or bromination". Nature Structural Biology 7 (9): 758–761. PMID 10966645. doi:10.1038/78985.
- ↑ Hayashi G, Hagihara M, Nakatani K (2005). "Application of L-DNA as a molecular tag". Nucleic Acids Symp Ser (Oxf) 49 (49): 261–262. PMID 17150733. doi:10.1093/nass/49.1.261.
- ↑ Allemand JF, Bensimon D, Lavery R, Croquette V (1998). "Stretched and overwound DNA forms a Pauling-like structure with exposed bases". PNAS 95 (24): 14152–14157. Bibcode:1998PNAS...9514152A. PMC 24342. PMID 9826669. doi:10.1073/pnas.95.24.14152.
- ↑ "List of 55 fiber structures". Arquivado dende o orixinal o 26 de maio de 2007. Consultado o 20 de marzo de 2015.
- ↑ Bansal M (2003). "DNA structure: Revisiting the Watson-Crick double helix". Current Science 85 (11): 1556–1563.
- ↑ Ghosh A, Bansal M (2003). "A glossary of DNA structures from A to Z". Acta Cryst D59 (Pt 4): 620–626. PMID 12657780. doi:10.1107/S0907444903003251.
- ↑ Sinden, Richard R (1994-01-15). DNA structure and function (1st ed.). Academic Press. p. 398. ISBN 0-12-645750-6.
- ↑ Rich A, Norheim A, Wang AHJ (1984). "The chemistry and biology of left-handed Z-DNA". Annual Review of Biochemistry 53: 791–846. PMID 6383204. doi:10.1146/annurev.bi.53.070184.004043.
- ↑ Ho PS (1994-09-27). "The non-B-DNA structure of d(CA/TG)n does not differ from that of Z-DNA". Proc Natl Acad Sci USA 91 (20): 9549–9553. Bibcode:1994PNAS...91.9549H. PMC 44850. PMID 7937803. doi:10.1073/pnas.91.20.9549.
- ↑ Wing R, Drew H, Takano T, Broka C, Tanaka S, Itakura K, Dickerson R (1980). "Crystal structure analysis of a complete turn of B-DNA". Nature 287 (5784): 755–8. Bibcode:1980Natur.287..755W. PMID 7432492. doi:10.1038/287755a0.
- ↑ Stokes, T. D. (1982). "The double helix and the warped zipper—an exemplary tale". Social Studies of Science 12 (2): 207–240. doi:10.1177/030631282012002002.
- ↑ Gautham, N. (25 May 2004). "Response to 'Variety in DNA secondary structure'". Current Science 86 (10): 1352–1353. Arquivado dende o orixinal o 13 de abril de 2020. Consultado o 25 May 2012.
Porén, o descubrimento das topoisomerases quitou a "pulla" das obxeccións topolóxicas á dobre hélice plectonémica. A solución máis recente da estrutura de raios X de cristal único da partícula central do nucleosoma mostrou case 150 pares de bases de ADN (é dicir, uns 15 xiros completos), cunha estrutura que é en todos os aspectos esenciais a mesma do modelo de Watson e Crick. Isto acabou coa idea de que as outras formas de ADN, especialmente no ADN en dobre hélice, existían simplemente como estruturas transitorias locais.
Véxase tamén
[editar | editar a fonte]Outros artigos
[editar | editar a fonte]| Commons ten máis contidos multimedia sobre: Dobre hélice |
- ADN tricatenario, ADN con 3 febras
- G-cuádruplex
- Nanotecnoloxía do ADN
- Modelos moleculares do ADN