Sacos aéreos
Os sacos aéreos son espazos dentro dun organismo que conteñen permanentemente aire. Nos animais modernos, as aves son as que posúen maior cantidade de sacos aéreos (de 9 a 11), que melloran o fluxo de aire durante a súa respiración e fan o seu corpo máis livián, pero os seus parentes os dinosauros extintos mostraban xa un grande incremento na pneumatización dos seus ósos (presenza de aire).[1] Os terópodos, como Aerosteon, tiñan moitos sacos aéreos no seu corpo e non simplemente nos ósos, e poden ser identificados como as formas máis primitivas que tiñan vías aéreas como as das modernas aves.[2] Os saurópodos tiñan unha gran cantidade de bolsas aéreas nos seus ósos (especialmente nas vértebras), aínda que un terópodo, Deinocheirus, tiña unha cantidade comparable.[3]
Na respiración das aves
[editar | editar a fonte]

As aves teñen nove sacos aéreos non vascularizados, situados entre os órganos e conectados aos pulmóns e aos ósos (os seus ósos teñen aire no sitio da medula). Estes sacos divídense nun grupo anterior, formado polos sacos interclavicular, os cervicais e os torácicos anteriores, e un grupo posterior, fomado polos torácicos posteriores e abdominais. Todos son pares menos o interclavicular, que só hai un. Os sacos anteriores, os pulmóns e os posteriores están conectados. Os sacos aéreos e as súas conexións permiten unha eficaz ventilación cun fluxo de aire case continuo que pasa polos pulmóns, o que proporciona o oxíxeno necesario para o elevado metabolismo das aves, e facilita a extracción de oxíxeno incluso se está en baixas concentracións. O intercambio de gases realízase nos pulmóns, onde hai un sistema de parabronquios e capilares aéreos (non teñen alvéolos coma os dos mamíferos) rodeados de capilares sanguíneos. A traquea da ave divídese en dous bronquios principais (mesobronquios), que se dirixen aos sacos aéreos posteriores (non aos pulmóns) e divídense sucesivamene en bronquios cada vez menores con moitas conexións entre eles ata acabar nos mencionados parabronquios dos pulmóns.[4]
Cando a ave respira o aire pasa polos sacos aéreos e os pulmóns realizándose dous ciclos completos de inhalación e exhalación, que se poden resumir así:
- 1ª inhalación. O aire entra na traquea e bronquios primarios, e parte del pasa aos pulmóns e parte aos sacos aéreos posteriores.
- 1ª exhalación. O aire dos sacos posteriores pasa aos pulmóns e despraza ao aire estancado que sae pola traquea.
- 2ª inhalación. Parte do aire que entra pasa aos sacos aéreos posteriores e parte aos pulmóns, empurrando o aire estancado dos pulmóns cara aos sacos anteriores.
- 2ª exhalación. O aire dos sacos anteriores e o dos pulmóns sae ao exterior, e é substituído polo aire que pasa aos pulmóns procedente dos sacos posteriores.[4]
Función nos dinosauros
[editar | editar a fonte]Desde aproximadamente 1870 en adiante os científicos concordan en xeral en que os esqueletos poscraniais de moitos dinosauros contiñan moitas cavidades cheas de aire (ósos pneumáticos), especialmente nas vértebras. A pneumatización do cranio (como nos seos paranasais) encóntrase tanto en sinápsidos coma en arcosauros, pero a pneumatización poscranial encóntrase só en aves, dinosuros saurisquios non aviarios e pterosauros.
Durante longo tempo estas cavidades foron consideradas simplemente como adaptacións para aforrar peso, pero Bakker propuxo que estaban conectados a sacos aéreos como os das aves, os cales fan que o sistema respiratorio das aves sexa o máis eficiente de todos os animais.[5]
John Ruben et al. (1997, 1999, 2003, 2004) discreparon disto e suxeriron que os dinosauros tiñan un sistema respiratorio "mareal" (de entrada e saída) impulsado por un mecanismo de pistón hepático coma o dos crocodilianos, no cal os músculos unidos principalmente ao pube tiran do fígado cara a atrás, o que fai que os pulmóns se expandan e inhalen (inspiración); cando estes músculos e relaxan, os pulmóns volven ao seu tamaño e forma previas, e o animal exhala (expiración). Tamén presentaron isto como unha razón para dubidar de que as aves descendesen dos dinosauros (do cal non se dubida na opinión científica xeral).[6][7][8][9][10]
Os críticos afirmaron que, sen sacos aéreos como os das aves, pequenas melloras nuns poucos aspectos nos sistemas respiratorio e circulatorio dun réptil moderno permitiríanlles aos réptiles obter do 50 ao 70% do fluxo de oxíxeno dun mamífero de similar tamaño,[11] e que a falta de sacos aéreos non impediría o desenvolvemento da endotermia.[12] Publicáronse moi poucas refutacións en revistas científicas da afirmación de Ruben et al. de que os dinosauos non tiñan sacos aéroes do estilo dos das aves; pero unha delas indica que o fósil Sinosauropteryx no cal eles basearan gran parte da súa argumentación estaba moi esmagado e, por tanto, era imposible dicir se o fígado tiña a forma axeitada para actuar como parte dun mecanismo de pistón hepético.[13] Algunhas publicacións recentes simplemente fan notar sen maior comentario que Ruben et al. argumentaron contra a presenza de sacos aéreos nos dinosauros.[14]
Probas da presenza de sacos aéreos en dinosauros
[editar | editar a fonte]Os investigadores presentaron probas e argumentacións en favor da presenza de sacos aéreos en saurópodos, "prosaurópodos", celurosauros, ceratosauros e nos terópodos Aerosteon e Coelophysis.
Nos saurópodos avanzados ("neosaurópodos") as vértebras da rexión lumbar e das cadeiras mostran signos da presenza de sacos aéreos. Nos saurópodos iniciais só as vértebras cervicais (do pescozo) mostraban esta característica. Se a secuencia de desenvolvemento observada en embrións de aves pode servir de guía, os sacos aéreos terían evolucionado en realidade antes das canles nos ósos do esqueleto que os albergan en formas posteriores.[15][16]
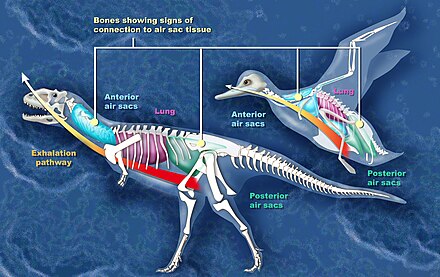
Tamén se encontraron probas de sacos aéreos en terópodos. Os estudos realizados indican que os celurosauros fósiles,[17] ceratosauros,[14] e os terópodos Coelophysis e Aerosteon mostran evidencias de sacos aéreos. Coelophysis, do Triásico tardío, é un dos primeiros dinosauros cuxos fósiles mostran probas de ter canles para sacos aéreos.[16] Aerosteon, un alosauro do Cretáceo tardío, tiña os sacos aéreos máis parecidos aos das aves atopados ata agora.[18]
Os primeiros sauropodomorfos, incluíndo o grupo tradicionalmente chamado "prosaurópodos", puido ter tamén sacos aéreos. Aínda que se atoparon posibles indentacións pneumáticas en Plateosaurus e Thecodontosaurus, estas indentacións son moi pequenas. Un estudo de 2007 concluíu que os prosaurópodos probablemente tiñan sacos abdominais e cervicais, baseándose nas probas que hai deles en taxons irmáns (terópodos e saurópodos). O estudo concluíu que era imposible determinar se os prosaurópodos tiñan un pulmón cun fluxo de aire similar ao das aves, pero eses sacos aéreos estaban case sempre presentes.[19] Outra indicación da presenza de sacos aéreos e o seu uso na vetilación pulmonar procede da reconstrución do volume de intercambio de aire (o volume de aire intercambiado en cada respiración) de Plateosaurus, que cando se expresa como a proporción de volume de aire por peso corporal en 29 ml/kg é similar aos valores observados nos gansos e outras aves, e moito maior que os valores típicos en mamíferos.[20]
Ata agora non se atopou ningunha evidencia de sacos aéreos en dinosauros ornitisquios. Pero isto non implica que os ornitisquios non puidesen ter taxas metabólicas comparables ás dos mamíferos, xa que os mamíferos tampouco teñen sacos aéreos.[21]
Desenvolvemento
[editar | editar a fonte]Propuxéronse tres explicacións para o desenvolvemento de sacos aéreos en dinosauros:[18]
- Incremento da capacidade respiratoria. Esta é probablemente a hipótese máis común, e concorda ben coa idea de que moitos dinosauros teñen taxas metabólicas bastante altas.
- Mellora do equilibrio e manobrabilidade ao facer descender o centro de gravidade e reducir a inercia rotacional. Porén, isto non explica a expansión dos sacos aéreos nos saurópodos cuadrúpedes.
- Como mecanismos de arrefriamento corporal. Parece que os sacos aéreos e as plumas evolucionaron aproximadamente ao mesmo tempo en coelurosauros. Se as plumas retiñan a calor, os dinosauros emplumados tiveron que ter algún medio para disipar o exceso de calor. Esta idea é plausible pero necesita de maior apoio empírico.
Os cálculos dos volumes de varias partes do sistema respiratorio do saurópodo Apatosaurus apoian as evidencias que que os saurópodos tiñan sacos aéreos similares aos das aves:
- Asumindo que Apatosaurus, igual que os parentes superviventes máis próximos aos dinosauros, que son os crocodilianos e as aves, non tiñan un músculo diafragma, o volme de espazo morto dun espécime de 30 toneladas sería duns 184 litros. Este é o volume total da boca, traquea e vías aéreas. Se o animal exhala menos volume de aire que este, o aire estancado non será expulsado e será enviado cara aos pulmóns na seguinte inhalación.
- As estimacións do seu volume mareal (a cantidade de aire movido cara a fóra e a dentro dos pulmóns nunha soa respiración) depende do tipo de sistema respiratorio que teña o animal: 904 litros se é ave; 225 litros nos mamíferos, e 19 litros en réptiles.
Baseándose nisto, Apatosaurus non podería ter un sistema respiratorio reptiliano, xa que o volume mareal tería sido menor que o seu volume de espazo morto, polo que o aire estancado non podería ser explulsado, senón que acabaría pasando aos pulmóns. Igualmente, un sistema como o dos mamíferos só proporcionaría aos pulmóns uns 225 − 184 = 41 litros de aire fresco oxixenado en cada respiración. Apatosaurus debeu, por tanto, ter ou un sistema descoñecido no mundo moderno ou un similar ao das aves, con moitos sacos aéreos e un fluxo a través dos pulmóns. Ademais, un sistema aviario necesitaría só un volume pulmonar duns 600 litros mentres que un de mamífero requiriría uns 2 950 litros, o que excedería os 1 700 litros estimados de espazo dispoñible no peito dun Apatosaurus′ de 30 toneladas.[22]
Os sistemas respiratorios dos dinosauros con sacos aéreos de tipo aviario puideron ter a capacidade de soster maiores niveis de actividade que a dos mamíferos de similar tamaño. Ademais de fornecer do oxíxeno necesario de forma moi eficaz, o rápido fluxo de aire tería sido un mecanismo de arrefriamento moi efectivo, que é esencial para animais que son activos pero demasiado grandes como para eliminar todo o exceso de calor pola súa pel.[21]
O paleontólogo Peter Ward argumentou que a evolución do sistema de sacos aéreos, que apareceu primeiro nos dinosauros máis iniciais, puido ser unha resposta á atmosfera moi baixa en oxíxeno (11%) dos pisos Carniano e Noriano do período Triásico.[23]
Notas
[editar | editar a fonte]- ↑ Romer, A.S.; Parsons, T.S. (1977). The Vertebrate Body. Holt-Saunders International. pp. 330–334. ISBN 0-03-910284-X.
- ↑ Sereno, P.C.; Martinez, R.N.; Wilson, J.A.; Varricchio, D.J.; Alcober, O.A.; Larsson, H.C.E. (2008). Kemp, Tom, ed. "Evidence for Avian Intrathoracic Air Sacs in a New Predatory Dinosaur from Argentina". PLOS ONE 3 (9): e3303. Bibcode:2008PLoSO...3.3303S. PMC 2553519. PMID 18825273. doi:10.1371/journal.pone.0003303. Arquivado dende o orixinal o 08 de outubro de 2008. Consultado o 10 de agosto de 2017.
- ↑ Lee, Y.N.; Barsbold, R.; Currie, P.J.; Kobayashi, Y.; Lee, H.J.; Godefroit, P.; Escuillié, F.O.; Chinzorig, T. (2014). "Resolving the long-standing enigmas of a giant ornithomimosaur Deinocheirus mirificus". Nature 515 (7526): 257–260. Bibcode:2014Natur.515..257L. PMID 25337880. doi:10.1038/nature13874.
- ↑ 4,0 4,1 K. V. Kardong. Vertebrados. Anatomía comparada, función, evolución. McGraw Hill-Interamericana. 2ª edición (1999) Páxinas 402-403. ISBN 84-486-0253-6
- ↑ Bakker, Robert T. (1972). "Anatomical and ecological evidence of endothermy in dinosaurs". Nature 238 (5359): 81–85. Bibcode:1972Natur.238...81B. doi:10.1038/238081a0.
- ↑ Ruben, J.A.; Jones, T.D.; Geist, N.R. & Hillenius, W. J. (November 1997). "Lung structure and ventilation in theropod dinosaurs and early birds". Science 278 (5341): 1267–1270. Bibcode:1997Sci...278.1267R. doi:10.1126/science.278.5341.1267.
- ↑ Ruben, J.A.; Jones, T.D.; Geist, N.R.; Leitch, A. & Hillenius, W.J. (1997). "Lung ventilation and gas exchange in theropod dinosaurs". Science 278 (5341): 1267–1270. Bibcode:1997Sci...278.1267R. doi:10.1126/science.278.5341.1267.
- ↑ Ruben, J.A.; Dal Sasso, C.; Geist, N.R.; Hillenius, W. J.; Jones, T.D. & Signore, M. (January 1999). "Pulmonary function and metabolic physiology of theropod dinosaurs". Science 283 (5401): 514–516. Bibcode:1999Sci...283..514R. PMID 9915693. doi:10.1126/science.283.5401.514.
- ↑ Ruben, J. A.; Jones, T. D. & Geist, N. R. (2003). "Respiration and reproductive paleophysiology of dinosaurs and early birds". Physiol. Biochem. Zool. 76 (2): 141–164. PMID 12794669. doi:10.1086/375425.
- ↑ Hillenius, W. J.; Ruben, J.A. (November–December 2004). "The Evolution of Endothermy in Terrestrial Vertebrates: Who? When? Why?". Physiological and Biochemical Zoology 77 (6): 1019–1042. PMID 15674773. doi:10.1086/425185.
- ↑ Hicks, J.W.; Farmer, C.G. (November 1997). "Lung Ventilation and Gas Exchange in Theropod Dinosaurs". Science 278 (5341): 1267–1270. Bibcode:1997Sci...278.1267R. doi:10.1126/science.278.5341.1267.
- ↑ Hicks, J.W.; Farmer, C.G. (September 1999). "Gas exchange potential in reptilian lungs: implications for the dinosaur–avian connection". Respiration Physiology 117 (2–3): 73–83. PMID 10563436. doi:10.1016/S0034-5687(99)00060-2. Arquivado dende o orixinal o 08 de decembro de 2012. Consultado o 10 de agosto de 2017.
- ↑ Currie, P.J.; Chen, P-j. (December 2001). "Anatomy of Sinosauropteryx prima from Liaoning, northeastern China". Canadian Journal of Earth Sciences 38 (12): 1705–1727. Bibcode:2001CaJES..38.1705C. doi:10.1139/cjes-38-12-1705. Arquivado dende o orixinal o 18 de agosto de 2016. Consultado o 10 de agosto de 2017.
- ↑ 14,0 14,1 O'Connor, P.; Claessens, L. (July 2005). "Basic avian pulmonary design and flow-through ventilation in non-avian theropod dinosaurs". Nature 436 (7048): 253–256. Bibcode:2005Natur.436..253O. PMID 16015329. doi:10.1038/nature03716.
- ↑ Wedel, M.J. (2003). "Vertebral pneumaticity, air sacs, and the physiology of sauropod dinosaurs". Paleobiology 29 (2): 243–255. doi:10.1666/0094-8373(2003)029<0243:VPASAT>2.0.CO;2. Texto completo en liña en "Findarticles.com: Vertebral pneumaticity, air sacs, and the physiology of sauropod dinosaurs". Paleobiology. 2003. e "Vertebral pneumaticity, air sacs, and the physiology of sauropod dinosaurs" (PDF). Arquivado dende o orixinal (PDF) o 15 de febreiro de 2015. Consultado o 10 de agosto de 2017. Detailed anatomical analyses can be found at Wedel, M.J. (2003). "The Evolution of Vertebral Pneumaticity in Sauropod Dinosaurs". Journal of Vertebrate Paleontology 23 (2): 344–357. doi:10.1671/0272-4634(2003)023[0344:TEOVPI]2.0.CO;2.
- ↑ 16,0 16,1 Wedel, M.J. (June 2006). "Origin of postcranial skeletal pneumaticity in dinosaurs". Integrative Zoology 1 (2): 80–85. doi:10.1111/j.1749-4877.2006.00019.x. Arquivado dende o orixinal o 15 de setembro de 2019. Consultado o 10 de august de 2017.
- ↑ Naish, D.; Martill, D. M. & Frey, E. (June 2004). "Ecology, systematics and biogeographical relationships of dinosaurs, including a new theropod, from the Santana Formation (?Albian, Early Cretaceous) of Brazil". Historical Biology 16 (2–4): 57–70. doi:10.1080/08912960410001674200. Tamén se trata disto no blog de Naish, "Basal tyrant dinosaurs and my pet Mirischia". - nótese queMirischia era un celurosauro, que Naish cre que está relacionado con Compsognathus.
- ↑ 18,0 18,1 Sereno PC, Martinez RN, Wilson JA, Varricchio DJ, Alcober OA, et al. (September 2008). Kemp T, ed. "Evidence for Avian Intrathoracic Air Sacs in a New Predatory Dinosaur from Argentina". PLOS ONE 3 (9): e3303doi=10.1371/journal.pone.0003303. Bibcode:2008PLoSO...3.3303S. PMC 2553519. PMID 18825273. doi:10.1371/journal.pone.0003303. Consultado o 2008-09-29.
- ↑ Wedel, Mathew (2007). "What pneumaticity tells us about 'prosauropods', and vice versa" (PDF). Special Papers in Palaeontology 77: 207–222. Arquivado dende o orixinal (PDF) o 05 de xullo de 2008. Consultado o 2007-10-31.
- ↑ Mallison, H. (2010). "The digital Plateosaurus II: an assessment of the range of motion of the limbs and vertebral column and of previous reconstructions using a digital skeletal mount". Acta Palaeontologica Polonica 55 (3): 433–458. doi:10.4202/app.2009.0075.
- ↑ 21,0 21,1 Reid, R.E.H. (1997). "Dinosaurian Physiology: the Case for "Intermediate" Dinosaurs". En Farlow, J.O.; Brett-Surman, M.K. The Complete Dinosaur. Bloomington: Indiana University Press. pp. 449–473. ISBN 0-253-33349-0. Consultado o 2008-09-12.
- ↑ Paladino, F.V.; Spotila, J.R. & Dodson, P. (1997). "A Blueprint for Giants: Modeling the Physiology of Large Dinosaurs". En Farlow, J.O. & Brett-Surman, M.K. The Complete Dinosaur. Bloomington, Ind.: Indiana University Press. pp. 491–504. ISBN 0-253-21313-4.
- ↑ Ward, Peter (2006) Out of thin air: Dinosaurs, birds, and earth's ancient atmosphere Pages 159–198, National Academies Press. ISBN 9780309141239.
| Control de autoridades |
|
|---|