Cromatóforo

Os cromatóforos son células[1] (ou grupos de células nalgúns taxons) que conteñen pigmentos e reflicten a luz dando cor, que se encontran nunha ampla variedade de animais, como peixes, anfibios, réptiles, crustáceos ou cefalópodos. Tamén se usa o termo en bacterias fotosintetizadoras[2][3][4], diatomeas[5] e dinoflaxelados[6] para referirse a vesículas ou plastidios na célula con concentración de pigmentos. Os mamíferos e aves, en vez de cromatóforos teñen células chamadas melanocitos que lles dan a súa coloración.
Os cromatóforos son en gran medida responsables de xerar a cor da pel e dos ollos en animais de sangue frío. Xéranse na crista neural durante o desenvolvemento embrionario. Os cromatóforos maduros agrúpanse en subclases baseándose na súa cor (máis propiamente o "ton") baixo a luz branca, que son: xantóforos (amarelos), eritróforos (vermellos), iridóforos (reflectivos/iridescentes), leucóforos (brancos), melanóforos (negros/castaños) e cianóforos (azuis).
Algunhas especies poden cambiar rapidamente de cor por medio de mecanismos que translocan o pigmento e reorientan placas reflectoras nos cromatóforos. Este proceso, que a miúdo se usa como un tipo de camuflaxe, denomínase cambio de cor fisiolóxico ou metacrose.[7] Os cefalópodos como o polbo teñen órganos cromatóforos complexos controlados por músculos para mudar de cor, mentres que os vertebrados como os camaleóns xeran un efecto similar por sinalización celular. Eses sinais poden ser hormonas ou neurotransmisores e poden ser iniciados por cambios no estado de ánimo, temperatura, estrés ou cambios visibles no ambiente local. Os científicos estudan os cromatóforos para comprender certas doenzas humanas e como unha ferramenta para descubrir novos fármacos.
Clasificación
[editar | editar a fonte]O termo cromatóforo foi adoptado (a partir do precedente cromoforo usado polo italiano G. Sangiovanni no século XIX[8]) como o nome das células que levan pigmento derivadas da crista neural de vertebrados de sangue frío e cefalópodos. A propia palabra deriva das palabras gregas khrōma (χρωμα) que significa 'cor', e phoros (φορος) que significa 'portador'.[9] Por outra parte, a palabra cromatocito utilízase ás veces para as células responsables da cor de mamíferos e aves, pero só está identificado un tipo de cromatocito nesta clase de animais, o melanocito.[10]
Os cromatóforos empezaron a comprenderse o suficientemente ben na década de 1960, o que permitiu clasificalos segundo a súa aparencia. Este sistema de clasificación daquela época persiste ata hoxe, aínda que a bioquímica dos pigmentos pode ser máis útil para comprender cientificamente como funcionan as células.[11]
As moléculas que producen cor poden ser de dúas clases: biocromos e cores estruturais ou "esquemocromos".[12] Os biocromos son verdadeiros pigmentos como os carotenoides e pteridinas. Estes pigmentos absorben selectivamente partes do espectro visible da luz (luces de certas lonxitudes de onda) que forman parte da luz branca, mentres que permiten que outras lonxitudes de onda cheguen ao ollo do observador e produzan a sensación de cor. As cores estruturais prodúcense por combinacións de difraccións, reflexións ou dispersións da luz cando esta incide en estruturas que teñen unha escala dunha fracción da lonxitude de onda da luz. Moitas destas estruturas interfiren con algunhas lonxitudes de onda (cores) da luz e transmiten outras, simplemente debido á súa escala, polo que a miúdo producen iridescencia, xerando diferentes cores cando se observan en diferentes direccións.[13][14] Moitos animais teñen á vez cores estruturais e cores pigmentarias (biocromos); por exemplo as plumas do pavo teñen melanina castaña pero tamén iridescencia.[15][16]
Aínda que todos os cromatóforos conteñen pigmentos ou estruturas reflectantes (excepto cando se produciu unha mutación como o albinismo), non todas as células que conteñen pigmentos son cromatóforos. O grupo hemo da proteína hemoglobina, por exemplo, é un biocromo responsable de que o sangue teña cor vermella (aínda que a súa función é transportar oxíxeno). Atópase fundamentalmente nos eritrocitos, que son células que se forman ao longo de toda a vida na medula ósea, e non só durante o desenvolvemento embrionario. Por tanto, os eritrocitos son células pigmentadas que non se clasifican como cromatóforos.[17]

Xantóforos e eritróforos
[editar | editar a fonte]Os cromatóforos que conteñen grandes cantidades de pigmentos de pteridina amarelos denomínanse xantóforos; e os que son principalmente carotenoides vermellos ou laranxas denomínanse eritróforos.[11] Porén, as vesículas que conteñen pteridinas e os carotenoides ás veces encóntranse dentro da mesma célula, e nese caso a cor global depende da proporción de pigmentos vermellos e amarelos.[18] Por tanto, a distinción entre estes dous tipos de cromatóforos non sempre é clara.
A maioría dos cromatóforos poden xerar pteridinas a partir da guanosina trifosfato, pero os xantóforos parecen ter vías bioquímicas suplementarias que lles permiten acumular pigmentos amarelos. Ao contrario, os carotenoides son metabolizados e transportados a eritróforos. Isto foi demostrado primeiramente criando ras normalmente verdes cunha dieta restrinxida en caroteno a base de grilos. A ausencia de caroteno na dieta das ras supuxo que o "filtro" de cor vermella/laranxa de carotenoide non estaba presente nos seus eritróforos, e isto fixo que as ras tivesen unha aparencia azul en vez de verde.[19]
Iridóforos e leucóforos
[editar | editar a fonte]Os iridóforos, ás veces chamados guanóforos, son células pigmentarias que reflicten a luz usando placas cristalinas de quimiocromos formados por guanina.[20] Cando son iluminados xeran cores iridescentes debido á difracción da luz nas placas amontoadas. A orientación dos esquemocromos determina a natureza da cor observada.[21] Ao usar biocromos como filtros coloreados, os iridóforos crean uns efectos ópticos coñecidos como efecto Tyndall ou dispersión Rayleigh, producindo cores brillantes azuis ou verdes.[22]
Un tipo de cromatóforo relacionado é o leucóforo, que se encontra nalgúns peixes, especialmente no tapetum lucidum. Igual que os iridóforos, utilizan purinas (xeralmente guanina) cristalizadas para reflectir a luz. A diferenza dos iridóforos, os leucóforos presentan cristais máis organizados que reducen a difracción. Se reciben luz branca, producen un brillo branco. Igual que pasaba cos xantóforos e eritróforos, nos peixes a distinción entre iridóforos e leucóforos non sempre é evidente, mais, en xeral, considérase que os iridóforos xeran cores metálicas ou iridescentes, mentres que os leucóforos producen tons brancos reflexivos.[22]
Melanóforos
[editar | editar a fonte]
- Véxase tamén: Melanocito.
Os melanóforos conteñen eumelanina, un tipo de melanina, que ten cor negra ou castaña escura debido ás súas cualidades de absorción de luz. O pigmento está empaquetado en vesículas chamadas melanosomas e distribuído pola célula. A eumelanina xérase a partir do aminoácido tirosina nunha serie de reaccións químicas catalizadas encimaticamente. É un complexo químico que contén unidades de dihidroxiindol e dihidroxiindol-2-ácido carboxílico con algúns aneis pirrol.[23] O encima clave na síntese de melanina é a tirosinase. Cando esta proteína é defectiva, non se pode xerar melanina, o que dá lugar a certo tipo de albinismo. Nalgunhas especies de anfibios empaquétanse outros pigmentos xunto coa eumelanina. Por exemplo, identificouse un novo pigmento de cor vermella escura (cor viño) nos melanóforos de ras filomedusinas.[24] Posteriormente, este foi identificado como a pterorrodina, unha pteridina dímera que se acumula arredor dun núcleo de eumelanina, e está tamén presente nunha variedade de especies de ras arborícolas de Australia e Papúa Nova Guinea. Aínda que é probable que outras especies menos estudadas teñan pigmentos melanóforos complexos, é malia todo certo que a maioría dos melanóforos estudados ata agora conteñan exclusivamente eumelanina.[25]
Os humanos teñen unha soa clase de célula pigmentaria, que é o equivalente nos mamíferos dos melanóforos, que crea a cor da pel, pelos e ollos chamada melanocito. Por esta razón e polo gran número e cor contrastada das células que fai que xeralmente sexan fáciles de visualizar, estes son con diferenza os cromatóforos máis estudados. Porén, hai diferenzas entre a bioloxía dos melanóforos e a dos melanocitos. Ademais da eumelanina, os melanocitos poden xerar un pigmento amarelo/vermello chamado feomelanina.[26]

Cianóforos
[editar | editar a fonte]Case todos os azuis brillantes nos animais e plantas son orixinados por coloración estrutural en vez de por pigmentos. Porén, algúns tipos de peixe mandarín (Synchiropus splendidus) posúen vesículas de biocromo de cor ciano de estrutura química descoñecida en células chamadas cianóforos.[22] Aínda que parecen infrecuentes polo seu limitado rango taxonómico, pode haber cianóforos (e outros tipos de cromatóforos raros) noutros peixes e anfibios. Por exemplo, os cromatóforos de cores brillantes con pigmentos non definidos poden encontrarse en ras dardo velenosas e ras de cristal,[27] e describíronse cromatóforos dicromáticos atípicos, chamados eritro-iridóforos no peixe Pseudochromis diadema.[28]
Translocación do pigmento
[editar | editar a fonte]
Moitas especies poden translocar o pigmento dentro dos seus cromatóforos, o que dá lugar a un cambio na cor do corpo. Este proceso, coñecido como cambio de cor fisiolóxico, foi estudado con maior amplitude nos melanóforos, xa que a melanina é o pigmento máis escuro e visible. Na maioría das especies cunha derme relativamente fina, os melanóforos dérmicos tenden a ser aplanados e cubrir unha área superficial grande. Porén, nos animais con capas dérmicas grosas, como os réptiles adultos, os melanóforos dérmicos a miúdo forman unidades tridimensionais xunto con outros cromatóforos. Estas unidades de cromatóforos dérmicos (UCD ou, en inglés, DCU) constan dunha capa superior de xantóforos ou eritróforos, debaixo unha capa de iridóforos, e finalmente unha capa de melanóforos con forma de cesta con procesos que cobren os iridóforos.[29]
Ambos os tipos de melanóforos son importantes no cambio de cor fisiolóxico. Os melanóforos dérmicos aplanados a miúdo cobren outros cromatóforos, de modo que se o pigmento está disperso pola célula, a pel aparece escura. Cando o pigmento está agregado na parte central da célula, os pigmentos noutros cromatóforos están expostos á luz e a pel toma o seu ton de cor. Igualmente, despois de que se produce a agregación da melanina en unidades de cromatóforos dérmicos, a pel aparece verde pola filtración que fan os xantóforos (amarelos) da luz que se dispersou na capa de iridóforos. Coa dispersión da melanina, a luz xa non se dispersa e a pel aparece escura. Como os outros cromatóforos biocromáticos poden tamén translocar pigmentos, os animais con varios tipos de cromatóforos poden xerar un conxunto espectacular de cores de pel ao facer un bo uso do efecto divisional.[30][31]
O control e mecánica das translocacións rápidas de pigmento foi ben estudada en varias especies, en particular os anfibios e peixes teleósteos.[22][32] Demostrouse que ese proceso pode estar baixo control hormonal ou neuronal ou ambos os dous. Entre os compostos neuroquímicos que se sabe que translocan pigmentos están a noradrenalina, por medio do seu receptor na superficie dos melanóforos.[33] As principais hormonas implicadas na regulación da translocación parecen ser as melanocortinas, melatonina e hormona concentradora da melanina (MCH), que se producen principalmente na hipófise, glándula pineal e hipotálamo, respectivamente. Estas hormonas poden tamén ser xeradas de modo parácrino por células da pel. Na superficie do melanóforo, as hormonas activan receptores acoplados á proteína G específicos que, á súa vez, transducen o sinal ao interior da célula. As melanocortinas causan a dispersión do pigmento, mentres que a melatonina e a MCH causan a súa agregación.[34]
Identificáronse numerosos receptores de melanocortina, MCH e melatonina nos peixes[35] e ras,[36] incluíndo un homólogo do MC1R,[37] un receptor da melanocortina que regula a cor do pelo e da pel nos humanos.[38] Demostrouse que o MC1R é necesario no peixe cebra para a dispersión da melanina.[39] Dentro da célula, o AMP cíclico (AMPc) é un importante segundo mensaxeiro na translocación do pigmento. Por medio dun mecanismo que aínda non se comprende ben, o AMPc influencia outras proteínas como a proteína quinase A para dirixir motores moleculares levando vesículas que conteñen pigmentos ao longo de microtúbulos e microfilamentos.[40][41][42]
Adaptación ao fondo
[editar | editar a fonte]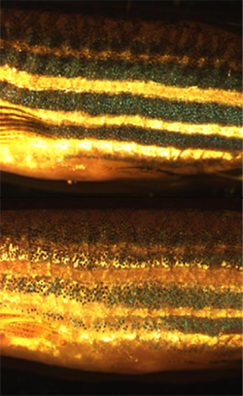
A maioría dos peixes, réptiles e anfibios poden realizar cambios de cor fisiolóxicos limitados en resposta aos cambios no seu ambiente. Este tipo de camuflaxe, coñecido como adaptación ao fondo, aparece máis comunmente como un ton lixeiramente máis escuro ou claro na pel para imitar aproximadamente o ton das contornas inmediatas nas que vive o animal. O proceso de adaptación ao fondo depende da visión (o animal necesita ver o aspecto do ambiente que o rodea para adaptarse a el)[43] e a translocación da melanina nos melanóforos é o principal factor no cambio de cor.[34] Algúns animais, como os camaleóns e os iguánidos anolis, desenvolveron unha resposta de adaptación ao fondo na que poden xerar moi rapidamente varias cores diferentes. Adaptaron a súa capacidade de mudar de cor como resposta a cambios de temperatura, estado de ánimo, niveis de estrés e comportamentos sociais, en vez de simplemente imitar o aspecto do seu medio ambiente.
Desenvolvemento
[editar | editar a fonte]
Durante o desenvolvemento embrionario de vertebrados, os cromatóforos son un dos tipos de células xeradas na crista neural, unha banda par de células que se orixinan nas marxes do tubo neural. Estas células teñen a capacidade de migrar a grandes distancias, o que permite que os cromatóforos poboen moitos órganos do corpo, como a pel, ollos, oídos e cerebro. Os cromatóforos abandonan a crista neural en ondadas e toman a ruta dorsolateral a través da derme, entrando no ectoderma a través de pequenos furados da lámina basal, ou ben toman unha ruta ventromedial entre os somitas e o tubo neural. A excepción a isto son os melanóforos do epitelio pigmentado da retina do ollo. Estes non derivan da crista neural, senón que unha bolsa que sobresae do tubo neural xera a copa óptica, a cal, á súa vez, forma a retina.
Investígase actualmente cando e como as células multipotentes precursoras dos cromatóforos, chamadas cromatoblastos se desenvolven nos seus subtipos fillos. Sábese que nos embrións do peixe cebra, por exemplo, uns 3 días despois da fertilización cada unha das clases de células que se encontran no peixe adulto (melanóforos, xantóforos e iridóforos) están xa presentes. En estudos con peixes mutantes demostrouse que factores de transcrición como son kit, sox10 e mitf son importantes para controlar a diferenciación dos cromatóforos.[44] Se estas proteínas son defectuosas, os cromatóforos poden estar ausentes rexionalmente ou completamente, o que orixina un trastorno leucístico.
Aplicacións prácticas
[editar | editar a fonte]Os cromatóforos utilízanse ás veces en investigación aplicada. Por exemplo, os individuos inmaturos de peixe cebra utilízanse para estudar como se organizan os cromatóforos e se comunican para xerar con precisión o patrón de bandas horizontais regular que ten o peixe adulto.[45] Este considérase un útil sistema modelo para comprender a formación de patróns no campo da bioloxía evolutiva do desenvolvemento. A bioloxía dos cromatóforos tamén se utilizou como modelo de doenzas ou condicións médicas humanas, como o melanoma e o albinismo. Recentemente, atopouse que o xene responsable da cepa de peixe cebra golden (dourada) específica de melanóforos Slc24a5 ten un equivalente humano que se correlaciona fortemente coa cor da pel.[46]
Os cromatóforos úsanse tamén como biomarcadores da cegueira en especies de sangue frío, xa que os animais con certos defectos visuais non poden adaptarse ao fondo do lugar iluminado onde viven.[43] Os homólogos humanos dos receptores que median na translocación do pigmento nos melanóforos pénsase que están implicados en procesos como a supresión do apetito e o bronceado polo sol, o que os fai dianas atractivas para fármacos.[37] Xa que logo, as compañías farmacéuticas desenvolveron unha proba biolóxica para identificar rapidamente potenciais compostos bioactivos utilizando os melanóforos do anfibio Xenopus laevis.[47] Outros científicos desenvolveron técnicas para usar os melanóforos como biosensores,[48] e para a detección rápida de doenzas (baseada no descubrimento de que a toxina pertussis bloquea a agregación dos pigmentos nos melanóforos de peixes).[49] Propuxéronse tamén potenciais aplicacións militares dos cambios de cor mediados por cromatóforos, principalmente como un tipo de camuflaxe activa, a cal podería facer os obxectos case invisibles, igual que fan os chocos.[50][51]
Cromatóforos de cefalópodos
[editar | editar a fonte]
Os cefalópodos Coleoidea (entre os que están os polbos, luras e chocos) teñen órganos multicelulares complexos que utilizan para cambiar de cor rapidamente, producindo unha ampla variedade de cores brillantes e patróns. Cada unidade cromatófora está composta dunha soa célula cromatófora e numerosos músculos, nervios, células da glía e células da vaíña.[52] Dentro da célula cromatófora, os gránulos de pigmento están pechados dentro dun saco elástico, chamado sáculo citoelástico. Para cambiar de cor o animal distorsiona a forma ou tamaño do sáculo por contracción muscular, cambiando o seu grao de transparencia, reflectividade, ou opacidade. Isto é diferente do mecanismo utilizado polos peixes, anfibios e réptiles porque a forma do sáculo cambia, en lugar de producirse unha translocación de vesículas con pigmento dentro da célula. Porén, o efecto conseguido é similar.
Os polbos poden facer funcionar os seus cromatóforos en complexas exhibicións cromáticas con forma de ondas, que dan lugar a unha variedade de esquemas de cambios de cor rápidos. As neuronas que controlan os cromatóforos crese que están situadas no cerebro nun patrón similar ao dos cromatóforos que controlan. Isto significa que os cambios de cor correspóndense cos patróns de activación neuronal. Isto pode explicar por que a cor cambia en ondas, xa que as neuronas se activan unhas a outras.[53] Igual que os camaleóns, os cefalópodos usan cambios de cor fisiolóxicos para a interacción social. Están tamén entre os máis habilidosos para a adaptación ao fondo, e teñen a capacidade de imitar tanto a cor como a textura do seu ambiente local con grande exactitude.
Bacterias e algas
[editar | editar a fonte]Tamén se utiliza o termo cromatóforo para referirse a certas partes coloreadas de bacterias e ás veces dalgunhas algas como os dinoflaxelados[54]. Nas membranas das bacterias fototróficas encóntranse cromatóforos, que se utilizan principalmente para facer a fotosíntese. Conteñen pigmentos bacterioclorofilas e carotenoides.[55] Nas bacterias púrpuras, como Rhodospirillum rubrum, as proteínas captadoras de luz son proteínas intrínsecas das membranas do cromatóforo. Porén, nas bacterias verdes do xofre, están dispostos en complexos antena especializados chamados clorosomas.[56]
Descubrimento
[editar | editar a fonte]Aristóteles xa menciona a capacidade do polbo de cambiar de cor para a camuflaxe e sinalización na súa Historia animalium (cerca de 400 a. de C.):[57]
O polbo ... busca a súa presa cambiando a súa cor para facerse da cor das pedras que o rodean; tamén o fai cando está asustado.
Giosuè Sangiovanni foi o primeiro que describiu as células de invertebrados que levan pigmentos dándolles o nome de cromoforo nunha revista científica italiana en 1819.[8] Actualmente cromóforo úsase con outro significado.[58]
Charles Darwin describiu as capacidade de cambiar de cor do choco no seu The Voyage of the Beagle (1860):[59]
Estes animais tamén escapan da detección por un extraordinario poder parecido ao do camaleón de mudar de cor. Parecen variar os seus tons de acordo coa natureza do terreo por onde pasan: cando están en augas profundas, o seu ton xeral é púrpura acastañado, pero cando se sitúan en terra, ou en augas pouco profundas, este ton escuro cambia a verde amarelado. A cor, examinada máis coidadosamente, era un verde francés, con numerosas pintas diminutas amarelas brillantes: a primeira destas variaba en intensidade; a última desaparecía completamente e aparecía outra vez alternadamente. Estes cambios efectuábanse de tal maneira que nubes de cor, que variaban en ton entre a cor do xacinto e a da castaña, estaban pasando continuamente polo seu corpo. Calquera parte, ao ser sometida a un lixeiro choque de galvanismo, facíase case branca: un efecto similar, pero en menor grao, producíase ao rañar a pel cunha agulla. Estas nubes de cor, ou rubores como se lles pode chamar, dise que se producen pola expansión e contracción alternada de diminutas vesículas que conteñen fluídos de cores variadas.
Notas
[editar | editar a fonte]- ↑ Dictionary.com Chromatophore in zoology
- ↑ Nas bacterias fotosintetizadoras os cromatóforos son vesículas de cor asociadas á membrana e nas que se realiza a fotosíntese.
- ↑ Lascelles, June (1962). J. gen. Microbiology, ed. "The Chromatophores of Photosynthetic Bacteria". Gran Bretaña.
- ↑ Salton, MR (1987). "Bacterial membrane proteins". Microbiological sciences (4 ed.) 4: 100–5. PMID 3153178.
- ↑ Werner, Dietrich (1977). University of California Press, eds. The Biology of Diatoms. ISBN 978-0-520-03400-6.
- ↑ Tait, R. V. (2013-10-22). Elsevier, eds. Elements of Marine Ecology: An Introductory Course. ISBN 978-1-4831-4188-6.
- ↑ Scott M. Boback & Lynn M. Siefferman (2010). "Variation in Color and Color Change in Island and Mainland Boas (Boa constrictor)". Journal of Herpetology 44 (4): 506–515. doi:10.1670/09-026.1.
- ↑ 8,0 8,1 Sangiovanni G. Descrizione di un particolare sistema di organi cromoforo espansivo-dermoideo e dei fenomeni che esso produce, scoperto nei molluschi cefaloso. G. Enciclopedico Napoli. 1819; 9:1–13.
- ↑ "Chromatophore". Online Etymology Dictionary. Consultado o 10 de marzo de 2016.
- ↑ Lapedriza, Alberto; Petratou, Kleio; Kelsh, Robert N. "14. Neural Crest Cells and Pigmentation". En Paul Trainor. Neural Crest Cells: Evolution, Development and Disease. Academic Press. p. 288. ISBN 9780124017306.
- ↑ 11,0 11,1 Bagnara, JT (1966). "Cytology and cytophysiology of non-melanophore pigment cells". International Review of Cytology. International Review of Cytology 20: 173–205. ISBN 978-0-12-364320-9. PMID 5337298. doi:10.1016/S0074-7696(08)60801-3.
- ↑ Fox, DL. (1976). Animal Biochromes and Structural Colors: Physical, Chemical, Distributional & Physiological Features of Colored Bodies in the Animal World. Berkeley: University of California Press. ISBN 0-520-02347-1.
- ↑ University of Exeter, ed. (setembro 1998). "Iridescence in Lepidoptera". Photonics in Nature (originally in Physics Review). Arquivado dende o orixinal o 19 de novembro de 2015. Consultado o 27 de abril de 2012.
- ↑ Wallin, Margareta (2002). "Nature's Palette: How animals, including humans, produce colours" (PDF). Bioscience explained 1 (2): 1–12. Consultado o 17 de novembro de 2011.
- ↑ Smyth, S. (2007). "What Makes the Peacock Feather Colorful?" (PDF). NNIN REU Journal.)
- ↑ Smyth, S. (2009). University of Alaska, Fairbanks (Honors Thesis), ed. "What Makes the Peacock Feather Bright and Colorful". Arquivado dende o orixinal o 04 de marzo de 2016. Consultado o 09 de marzo de 2016.
- ↑ Masato Kinoshita, Kenji Murata, Kiyoshi Naruse, Minoru Tanaka. Medaka: Biology, Management, and Experimental Protocols. Google books. Páxina 119. [1]
- ↑ Matsumoto, J (1965). "Studies on fine structure and cytochemical properties of erythrophores in swordtail, Xiphophorus helleri, with special reference to their pigment granules (pterinosomes)". J Cell Biol 27 (3): 493–504. PMC 2106771. PMID 5885426. doi:10.1083/jcb.27.3.493.
- ↑ Bagnara JT. (1998). Comparative Anatomy and Physiology of Pigment Cells in Nonmammalian Tissues. En: The Pigmentary System: Physiology and Pathophysiology. Oxford University Press. ISBN 0-19-509861-7.
- ↑ Taylor, JD. (1969). "The effects of intermedin on the ultrastructure of amphibian iridophores". Gen Comp Endocrinol 12 (3): 405–16. PMID 5769930. doi:10.1016/0016-6480(69)90157-9.
- ↑ Morrison, RL. (1995). "A transmission electron microscopic (TEM) method for determining structural colors reflected by lizard iridophores". Pigment Cell Res 8 (1): 28–36. PMID 7792252. doi:10.1111/j.1600-0749.1995.tb00771.x.
- ↑ 22,0 22,1 22,2 22,3 Fujii, R. (2000). "The regulation of motile activity in fish chromatophores". Pigment Cell Res 13 (5): 300–19. PMID 11041206. doi:10.1034/j.1600-0749.2000.130502.x.
- ↑ Ito, S; Wakamatsu, K. (2003). "Quantitative analysis of eumelanin and pheomelanin in humans, mice, and other animals: a comparative review". Pigment Cell Res 16 (5): 523–31. PMID 12950732. doi:10.1034/j.1600-0749.2003.00072.x.
- ↑ Bagnara, J.T.; Taylor, JD; Prota, G (1973). "Color changes, unusual melanosomes, and a new pigment from leaf frogs". Science 182 (4116): 1034–5. PMID 4748673. doi:10.1126/science.182.4116.1034.
- ↑ Bagnara, J.T. (2003). "Enigmas of Pterorhodin, a Red Melanosomal Pigment of Tree Frogs". Pigment Cell Research 16 (5): 510–516. PMID 12950730. doi:10.1034/j.1600-0749.2003.00075.x.
- ↑ Hunt G, Kyne S, Ito S, Wakamatsu K, Todd C, Thody A. (1995 agosto). "Eumelanin and phaeomelanin contents of human epidermis and cultured melanocytes". Pigment Cell Res. (8(4)): 202–8. PMID 8610071.
- ↑ Schwalm, PA; Starrett, PH; McDiarmid, RW (1977). "Infrared reflectance in leaf-sitting neotropical frogs". Science 196 (4295): 1225–7. PMID 860137. doi:10.1126/science.860137.
- ↑ Goda M, Ohata M, Ikoma H, Fujiyoshi Y, Sugimoto M, Fujii R (2011). "Integumental reddish-violet coloration owing to novel dichromatic chromatophores in the teleost fish, Pseudochromis diadema". Pigment Cell Melanoma Res 24 (4): 614–7. PMID 21501419. doi:10.1111/j.1755-148X.2011.00861.x.
- ↑ Bagnara, JT; Taylor, JD; Hadley, ME (1968). "The dermal chromatophore unit". J Cell Biol 38 (1): 67–79. PMC 2107474. PMID 5691979. doi:10.1083/jcb.38.1.67.
- ↑ Palazzo, RE; Lynch, TJ; Lo, SJ; Taylor, JD; Tchen, TT (1989). "Rearrangements of pterinosomes and cytoskeleton accompanying pigment dispersion in goldfish xanthophores". Cell Motil Cytoskeleton 13 (1): 9–20. PMID 2543509. doi:10.1002/cm.970130103.
- ↑ Porras, MG; De Loof, A; Breuer, M; Aréchiga, H (2003). "Procambarus clarkii". Peptides 24 (10): 1581–9. PMID 14706537. doi:10.1016/j.peptides.2003.08.016.
- ↑ Deacon, SW; Serpinskaya, AS; Vaughan, PS; Lopez Fanarraga, M; Vernos, I; Vaughan, KT; Gelfand, VI (2003). "Dynactin is required for bidirectional organelle transport". The Journal of Cell Biology 160 (3): 297–301. PMC 2172679. PMID 12551954. doi:10.1083/jcb.200210066.
- ↑ Aspengren, S; Sköld, HN; Quiroga, G; Mårtensson, L; Wallin, M (2003). "Noradrenaline- and melatonin-mediated regulation of pigment aggregation in fish melanophores". Pigment Cell Res 16 (1): 59–64. PMID 12519126. doi:10.1034/j.1600-0749.2003.00003.x.
- ↑ 34,0 34,1 Logan, D. W.; Burn, SF; Jackson, IJ (2006). "Regulation of pigmentation in zebrafish melanophores". Pigment Cell Research 19 (3): 206–213. PMID 16704454. doi:10.1111/j.1600-0749.2006.00307.x.
- ↑ Logan, DW; Bryson-Richardson, RJ; Taylor, MS; Currie, P; Jackson, IJ (2003). "Sequence characterization of teleost fish melanocortin receptors". Ann N Y Acad Sci 994: 319–30. PMID 12851332. doi:10.1111/j.1749-6632.2003.tb03196.x.
- ↑ Sugden, D; Davidson, K; Hough, KA; Teh, MT (2004). "Melatonin, melatonin receptors and melanophores: a moving story". Pigment Cell Res 17 (5): 454–60. PMID 15357831. doi:10.1111/j.1600-0749.2004.00185.x.
- ↑ 37,0 37,1 Logan, DW; Bryson-Richardson, RJ; Pagán, KE; Taylor, MS; Currie, PD; Jackson, IJ (2003). "The structure and evolution of the melanocortin and MCH receptors in fish and mammals". Genomics 81 (2): 184–91. PMID 12620396. doi:10.1016/S0888-7543(02)00037-X.
- ↑ Valverde, P; Healy, E; Jackson, I; Rees, JL; Thody, AJ (1995). "Variants of the melanocyte-stimulating hormone receptor gene are associated with red hair and fair skin in humans". Nat Genet 11 (3): 328–30. PMID 7581459. doi:10.1038/ng1195-328.
- ↑ Richardson, J; Lundegaard, PR; Reynolds, NL; Dorin, JR; Porteous, DJ; Jackson, IJ; Patton, EE (2008). "mc1r Pathway regulation of zebrafish melanosome dispersion". Zebrafish 5 (4): 289–95. PMID 19133827. doi:10.1089/zeb.2008.0541.
- ↑ Snider, J; Lin, F; Zahedi, N; Rodionov, V; Yu, CC; Gross, SP (2004). "Intracellular actin-based transport: How far you go depends on how often you switch". Proceedings of the National Academy of Sciences of the United States of America 101 (36): 13204–9. PMC 516548. PMID 15331778. doi:10.1073/pnas.0403092101. Arquivado dende o orixinal o 10 de xuño de 2008. Consultado o 22 de decembro de 2014.
- ↑ Rodionov, VI; Hope, AJ; Svitkina, TM; Borisy, GG (1998). "Functional coordination of microtubule-based and actin-based motility in melanophores". Current biology : CB 8 (3): 165–8. PMID 9443917. doi:10.1016/S0960-9822(98)70064-8.
- ↑ Kashina, AS; Semenova, IV; Ivanov, PA; Potekhina, ES; Zaliapin, I; Rodionov, VI (2004). "Protein kinase A, which regulates intracellular transport, forms complexes with molecular motors on organelles". Current biology : CB 14 (20): 1877–81. PMID 15498498. doi:10.1016/j.cub.2004.10.003.
- ↑ 43,0 43,1 Neuhauss, SC. (2003). "Behavioral genetic approaches to visual system development and function in zebrafish". J Neurobiol 54 (1): 148–60. PMID 12486702. doi:10.1002/neu.10165.
- ↑ Kelsh, RN; Schmid, B; Eisen, JS (2000). "Genetic analysis of melanophore development in zebrafish embryos". Dev Biol 225 (2): 277–93. PMID 10985850. doi:10.1006/dbio.2000.9840.
- ↑ Kelsh, RN (2004). "Genetics and evolution of pigment patterns in fish". Pigment cell research / sponsored by the European Society for Pigment Cell Research and the International Pigment Cell Society 17 (4): 326–36. PMID 15250934. doi:10.1111/j.1600-0749.2004.00174.x.
- ↑ Lamason, RL; Mohideen, MA; Mest, JR; Wong, AC; Norton, HL; Aros, MC; Jurynec, MJ; Mao, X; et al. (2005). "SLC24A5, a putative cation exchanger, affects pigmentation in zebrafish and humans". Science 310 (5755): 1782–6. PMID 16357253. doi:10.1126/science.1116238.
- ↑ Jayawickreme, CK; Sauls, H; Bolio, N; Ruan, J; Moyer, M; Burkhart, W; Marron, B; Rimele, T; Shaffer, J (1999). "Use of a cell-based, lawn format assay to rapidly screen a 442,368 bead-based peptide library". J Pharmacol Toxicol Methods 42 (4): 189–97. PMID 11033434. doi:10.1016/S1056-8719(00)00083-6.
- ↑ Andersson, TP; Filippini, D; Suska, A; Johansson, TL; Svensson, SP; Lundström, I (2005). "Frog melanophores cultured on fluorescent microbeads: biomimic-based biosensing". Biosens Bioelectron 21 (1): 111–20. PMID 15967358. doi:10.1016/j.bios.2004.08.043.
- ↑ Karlsson, JO; Andersson, RG; Askelöf, P; Elwing, H; Granström, M; Grundström, N; Lundström, I; Ohman, L (1991). "The melanophore aggregating response of isolated fish scales: a very rapid and sensitive diagnosis of whooping cough". FEMS Microbiol Lett 66 (2): 169–75. PMID 1936946.
- ↑ Hansford, Dave (6 de agosto de 2008). "Cuttlefish Change Color, Shape-Shift to Elude Predators". National Geographic News (Wellington, Nova Zelandia).
[...] os chocos polo contrario confiaron na invisibilidade, un talento que pode ter aplicacións na tecnoloxía humana. Norman dixo que os militares mostraron interese na camuflaxe dos chocos con vistas a algún día incorporar mecanismos similares nos uniformes dos soldados.
- ↑ Lee I. Nanotubes for noisy signal processing PhD Thesis. 2005; Universidade do Sur de California.
- ↑ Cloney, RA; Florey, E (1968). "Ultrastructure of cephalopod chromatophore organs". Zeitschrift fur Zellforschung und mikroskopische Anatomie (Vienna, Austria: 1948) 89 (2): 250–80. PMID 5700268. doi:10.1007/BF00347297.
- ↑ Demski, LS (1992). "Chromatophore systems in teleosts and cephalopods: a levels oriented analysis of convergent systems". Brain, behavior and evolution 40 (2–3): 141–56. PMID 1422807. doi:10.1159/000113909.
- ↑ W. A. S. Sarjeant. Fossil and Living Dinoflagellates. Páxina 23. Google books. [2]
- ↑ Salton, MR (1987). "Bacterial membrane proteins". Microbiological sciences 4 (4): 100–5. PMID 3153178.
- ↑ Frigaard, NU; Bryant, DA (2004). "Seeing green bacteria in a new light: genomics-enabled studies of the photosynthetic apparatus in green sulfur bacteria and filamentous anoxygenic phototrophic bacteria". Archives of Microbiology 182 (4): 265–76. PMID 15340781. doi:10.1007/s00203-004-0718-9.
- ↑ Aristotle. Historia Animalium. IX, 622a: 2-10. Aproximadamente 400 BC. Citado en Luciana Borrelli, Francesca Gherardi, Graziano Fiorito. A catalogue of body patterning in Cephalopoda. Firenze University Press, 2006. Abstract Arquivado 06 de febreiro de 2018 en Wayback Machine. Google books
- ↑ "Chromophore". IUPAC Gold Book. Consultado o 10 de marzo de 2016.
- ↑ Darwin, Charles (1860). Journal Of Researches Into The Natural History And Geology Of The Countries Visited During The Voyage Round The World Of H.M.S. 'Beagle' Under The Command Of Captain Fitz Roy, R.N. Capítulo 1. Habits of a Sea-slug and Cuttle-fish. John Murray, Londres. p. 7.