Eukarioto
| Eukarioto | |
|---|---|
 | |
| Sailkapen zientifikoa | |
| Goierreinua | Eukaryota |
| Azpibanaketa | |
| |
Eukariotoak zelula nukleoa eta beste organulu batzuk zelula mintzaz inguratuta duten izaki bizidunak dira. Eukariotoak Eukarya domeinuaren parte dira, eta zelulabakar edo zelulaniztunak izan daitezke. Zelula eukariotoa prokariototik (hau da, bakterioak eta arkeoak) bereizten duen ezaugarria organuluak mintzaz babestuak egotea da, bereziki nukleoa. Nukleo horretan material genetikoa biltzen da, gaineztadura nuklearrren babesean[1][2][3]. Nukleoa izateak ematen die eukariotoei euren izena, antzinako grezierako εὖ (eu, "ondo" edo "benetakoa") eta κάρυον (karyon, "muin", "hazi" edo "mami")[4]. Gaineztadura edo mintzaz babestutako beste organulo batzuk ere badituzte zelula eukariotoek, adibidez mitokrondrioak eta Golgi aparatua. Gainera, landare eta algek kloroplastoak ere badituzte. Arkeobakterio eta bakterioek ez bezala, eukariotoak elkartu eta izaki zelulaniztunak sor ditzakete, ehun eta zelula mota ezberdinekin.
Eukarya domeinuaren barruan Protista, Fungi (onddoak), Plantae (landareak) eta Animalia (animaliak) izeneko erreinuak daude[5][6]. Eukariotoak asexualki ugal daitezke mitosiaren bidez, edo sexualki, meiosi eta gameto fusioaren bidez. Mitosian, zelula zatitzen da eta genetikoki identikoak diren beste bi zelula sortzen ditu. Meiosian, DNAren erreplikazioa bi aldiz egiten du zelula zatiketa, lau zelula haploide sortuz. Zelula hauek sexuaren bidez elkartzen dira, eta gameto izena dute. Gameto bakoitzak kromosoma multzo bakarra du, meiosiaren birkonbinazioan zehar sortutakoak[7].
Eukariotoak monofiletikoak direla ematen du, hau da, jatorri bera duten izakiz osatutako domeinua da. Hiru domeinu sistemako domeinuetako bat da, baina beste biek ez dute goian aipatutako jokamoldea[8]. Eukariotoak izaki bizidun guztien multzoaren zati txiki bat baino ez dira[9].
Egitura
[aldatu | aldatu iturburu kodea]Zelula eukariotiko gehienak mikroskopikoak badira ere, prokariotikoak baino askoz handiagoak dira: tipikoki hamar aldiz handiagoak luzeraz, eta gutxienez mila aldiz handiagoak bolumenari dagokionez. Badirudi tamaina maximoa elikagaien eta oxigenoaren difusio-tasak mugatzen duela. Tamainarekin batera noski osagai-kopurua ere oso altua da: zelula eukariotiko batean 10.000-20.000 proteina desberdin egon daitezke, eta proteinen kopuru osoa 10.000 milioi inguruko izan daiteke.
Egiturari dagokionez, zelula eukariotiko guztietan bi konpartimentu bereizten dira: zelula nukleoa eta zitoplasma. Zitoplasma gehienetan nukleoa baino handiagoa eta oso ondo antolatuta dago: hemen hainbat organulu eta egitura ezberdin bereizten dira eta prozesu anitz burutzen dira. Organulu zitoplasmatikoak mintz batez mugatutako barrunbeak dira, bakoitzak bere osagai espezifikoekin. Organulu bakoitzean funtzio konkretu bat (edo batzuk) burutzen da: zelula lantegi bat izango balitz organuluak funtzio edo prozesu ezpezifikoetaz arduratzen diren atalak izango lirateke.
Organuluak
[aldatu | aldatu iturburu kodea]
(2) Nukleoa
(3) Erribosomak (puntutxoak)
(4) Besikula
(5) Erretikulu endoplasmatiko bikortsua
(6) Golgiren aparatua
(7) Zitoeskeletoa
(8) Erretikulu endoplasmatikoa laua
(9) Mitokondrioa
(10) Bakuoloa
(11) Zitoplasma
(12) Lisosoma
(13) Zentrioloak bere zentrosomarekin
Animali zelula batean, organulu zitoplasmatikoek zelularen bolumenaren %50 inguru okupatzen dute. Orokorrean antzeko funtzioak betetzen dituzte zelula mota guztietan baina organulu bakoitzaren kopurua eta zenbait ezaugarri desberdinak dira zelula-motaren arabera. Organuluen garrantziaz ohartzeko kontuan hartu behar dugu organuluen mintzen azalera mintz plasmatikoarena baino 20 aldiz handiagoa izan daitekeela.
Erretikulu endoplasmatikoa
[aldatu | aldatu iturburu kodea]Erretikulu endoplasmatikoa (EE) bere izenak adierazten duen moduan sare konplexu bat da hodiz eta sakuluz osatua. Labirinto itxura duen organulu honen mintzak zelularen mintz guztien erdia izan daiteke. Zentro biosintetiko bat da: hemen lipido zelular gehienak eta proteina asko sintetizatu eta modifikatzen dira.
Golgiren aparatua
[aldatu | aldatu iturburu kodea]Multzoka agertzen diren zisterna zapalez osatuta dago. Hemen erretikulu endoplasmatikotik datozen produktuen biosintesiak jarraitzen du eta gainera produktu horien sailkapena burutzen da zelularen hemuga desberdinetara bidaltzeko.
Lisosomak
[aldatu | aldatu iturburu kodea]Liseri-entzimak gordetzen dituzten poltsak dira. Organulu hauetan molekula eta egitura anitz apurtzen dira; bai endozitosiaren bidez sartzen direnak, bai barneko osagaiak, eliminatuak izan behar direnak.
Endosomak
[aldatu | aldatu iturburu kodea]Endozitaturiko molekulak lisosometara iritsi baino lehen beste barrunbeetatik pasatzen dira, hauek endosomak ditugu: hemen kanpotik datozen produktuak sailkatzen dira.
Mitokondrioak
[aldatu | aldatu iturburu kodea]Mitokondrioak organulu handiak dira, mintz bikoitz batez inguraturik[10]. Hemen arnasketa burutzen da eta ATP molekulak ekoizten dira; gero ATPa zelula osoan erabiliko da hainbat prozesu zelularratan (nukleoan ere bai). Mitokondrioek oso ezaugarri berezia aurkezten dute: bere gai genetiko propioa dauka, proteina mitokondrial batzuk sintetizatzeko ezinbestekoa dena[11].
Peroxisomak
[aldatu | aldatu iturburu kodea]Entzimak gordetzen dituzten poltsak dira non hainbat erreakzio oxidatibo burutzen diren.
Zitosola
[aldatu | aldatu iturburu kodea]Zitoplasmatik organuluak eliminatzen baditugu geratzen den barrunbeari (eta hor dauden osagaiei) zitosola deitzen diogu. Zitosola askotan zelularen barrunberik handiena da: zelularen bolumenaren erdia baino gehiago hartzen du. Hemen zelularentzat funtsezkoak diren prozesu asko etengabe gertatzen ari dira.
Hasteko esan behar dugu zelularen proteina guztien sintesia zitosolean hasten dela. Hori dela eta erribosomak, itzulpenaz arduratzen diren egiturak zitosolean daude. Animalia zelula batean 10 milioi erribosoma egon daitezke, hauetariko batzuk aske daude eta beste batzuk EE-aren mintzei itsatsita. Gainera proteinen degradazioa ere hemen gertatzen da. Prozesu honetaz arduratzen diren egiturak proteasomak deritze; hauek, erribosomak bezala, elkarte makromolekularrak dira.
Bestalde zitosolean sintesi eta degradazio-erreakzio asko gertatzen ari dira etengabe eta aldiberean. Degradazioa batez ere energia lortzeko (ATP molekuletan metatzen dena) eta sintesia (aminoazido, nukleotido eta gantz azidoen biosintesia), makromolekulen monomeroak sortzeko. Dakigun bezala bide metabolikoak ez dira independenteak, gurutzatzen dira. Erreakzio hauen artean garrantzi berezia dauka glukolisiak: ATP lortzeko bide kataboliko non glukosaren degradazioa gertatzen den. Bestalde zitosola erregaien metalekua da. Zelulak erregaiak behar ditu energia sortzeko. Erregaiak disoluzioan dauden molekula txikiak izan daitezke, glukosa, alegia; baina erregaien erreserbak batez ere egitura ez-solugarrien moduan metatzen dira. Horrelako moduan energia-kopuru handia oso bolumen txikian metatzea lortzen da.
Erreserba-egitura garrantzitsuenak bi dira:
- glukogeno pikorrak, glukosaren polimero bat
- lipido-tantak, non gantz azidoak triglizerido moduan metatzen diren
Oso ugariak izan daitezke zenbait zelula-motatan; bereziki lipido-tantek adipozitoetan ia zitoplasma osoa okupatu ahal dute eta organulu bat baino askoz handiagoak izan daitezke. Orokorrean egitura hauek inklusio zitoplasmatikoak deitzen dira: gai espezfikoen metaketak, gehienetan zelularen jarduera biosintetikoaren ondorioz sortuak direnak.
Zitoeskeletoa
[aldatu | aldatu iturburu kodea]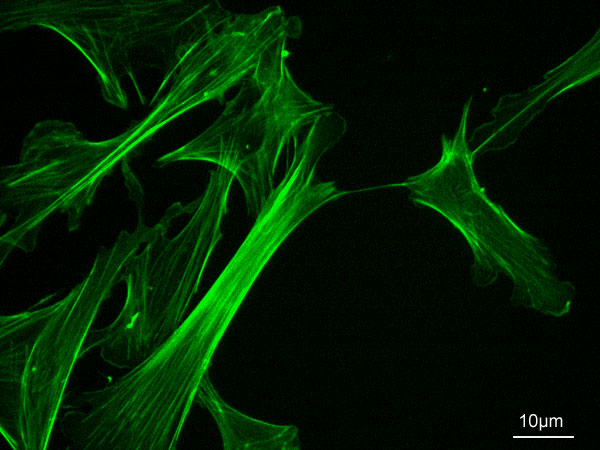
Hasieran ur disoluzioa bat besterik ez zela pentsatu zen; zehazki, ia likidoa den gel moduko disoluzio bat. Disoluzio honetan makromolekulak eta molekula txikiak disolbaturik zeuden. Gaur egun badakigu zitosola oso ondo antolatuta dagoela: sare konplexu bat osatzen duten zuntzez zeharkaturik dago; sare hau zitoeskeletoa da. Zitoeskeletoa beraz, zitosolaren funtsezko osagaia da: zitoeskeletoari esker zitosoleko osagaiak eta barrunbe honetan gertatzen diren erreakzio eta prozesuak espazialki antolatuta daude, hau da, kokapen espezifikoetan agertzen dira. Gainera itxura ematen dio zelulari, mugimendu zelularrez arduratzen da eta funtsezkoa da zatiketa zelularra burutzeko. Zitoeskeletoa firu proteiko ezberdinez osatuta dago; zehazki hiru firu-mota daude:
Garraio mekanismoak
[aldatu | aldatu iturburu kodea]Zelula eukariotoaren barrunbeak (zelula nukleoa, organulu zitoplasmatikoak eta zitosola) independenteak badira ere, ez dira egitura isolaturik; haien artean ordea elkartrukeak gertatzen ari dira etengabe. Hau ulertzeko kontuan hartu behar dugu barrunbe bakoitzaren osagaiak ez direla sortzen barrunbean bertan, alderantziz, askotan beste lekutan sintetizatzen dira; horregatik mekanismo espezifikoak beharrezkoak dira emandako helmuga batetara heltzeko.
Nukleoaren proteina guztiak esateko zitosolean ekoizten dira eta gero gaineztadura nuklearra zeharkatu behar dute nukleoan sartzeko. Zelulak garraio-sistema desberdinak erabiltzen ditu molekulak barrunbeen artean elkartrukatzeko. Zenbait organulu, erretikulu endoplasmatikoa, Golgiren aparatua, lisosomak eta endosomak hain zuzen, besikulen bidez komunikatuta daude:
- garraio-besikulak askatzen dira barrunbe batetatik (emailetik)
- fusionatzen dira beste barrunbe batekin (hartzailearekin).
Fenomeno honi trafiko besikularra deitzen diogu; oso konplexua da eta oso ondo erregulatuta dago. Garraio-xixkuak beraz, oso ugariak dira edozein animalia zelulatan. Trafiko honetan inplikatutako organuluak etengabe mintz-zatiak elkartrukatzen ari dira, hori dela eta esaten da barne-mintz sistema osatzen dutela[12]. Barnemintz sistematik kanpo mitokondrioak eta peroxisomak geratzen dira, hori dela eta, organulu hauen mintzetan translokadoreak daude osagai espezifikoen inportazioa burutzeko. Bestalde nukleoa eta zitosolaren arteko elkartrukeak nukleoa inguratzen duen gaineztaduraren egituren bidez burutzen da; horiek nukleo-poroak dira.
Mintz plasmatikoa
[aldatu | aldatu iturburu kodea]Zelula eukariotoaren zitoplasma mintz plasmatikoak mugatzen du. Mintz plasmatikoa, egiturari dagokionez, berdina da zelula guztietan, baina eukariotoetan mintz plasmatikoak oso espezializazio-maila altua lortu du. Gogoratu behar dugu prokariotoetan mintz plasmatikoan burutzen diren prozesu asko, ATParen sintesia esateko, eukariotoetan organulu zitoplasmatikoetan burutzen direla. Oro har esan dezakegu eukariotoen mintz plasmatikoa batez ere garraio, komunikazio eta adhesio prozesuetan inplikatuta dagoela, hau da, inguruneariko harremanekin zerikusia duten funtzioetan.
Bestalde animali zeluletan mintz plasmatikoak gaitasun bereziki garrantzitsua garatu da: endozitosi eta exozitosiaren gaitasuna; hau da, gaiak barneratzea edo kanporatzea mintz-zatiak erabiliz. Endozitatzen diren produktuak batzuetan oso handiak dira eta kasu honetan fagozitosiaz hitz egiten dugu. Prozesu hauek burutzeko zelularen azala oso malgua izan behar da, beraz, animalia zelulak biluziak dira, hau da, ez daukate zelula hormarik.
Nukleoa
[aldatu | aldatu iturburu kodea]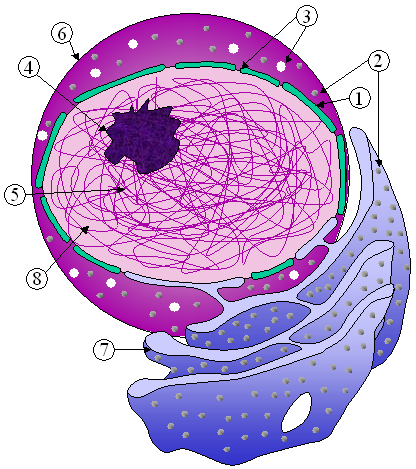
Nukleoan DNA zelular gehiena gordetzen da (baina ez guztia) eta zitoplasmatik banatuta dago gaineztadura nuklearra izeneko bi mintzez osatutako egituraren bidez Egitura honi esker gai genetikoa zitoplasman burutzen den jarduera metabolikotik eta mugimendu zitoplasmatikoetatik babestuta dago[13].
Zelula eukariotikoaren DNA kopurua oso altua da; giza zelula tipiko batek esate baterako, bakterio batek baino 1.000 aldiz DNA gehiago izaten du. Bestalde informazio genetikoa DNA molekula desberdinetan gordetzen da. DNA molekula eukariotikoak oso luzeak dira eta beti proteina espezifikoekin (histonak izeneko proteinekin hain zuzen) elkartuta daude; elkarte honi kromosoma deitzen diogu: DNA molekula bat gehi proteinak. Nukleoan kromosoma batzuk biltzen dira baina zelula zatiketan ez dagoenean hauek ez dira egitura trinkoak eta ez dira bereizten; hori dela eta kromosoma-multzo hau izendatzeko askotan beste termino bat erabiltzen da: kromatina[14][15].
Kromatinaz gain nukleoaren barruan beste egitura bat bereizten da: nukleoloa, erribosomak ekoizteko zentroa. Jarduera nuklearrari dagokionez esan behar dugu nukleoan RNA sintesia burutzen dela baina ez proteinen sintesia; beraz, gene-adierazpenaren bi prozesuak, transkripzioa eta itzulpena, banatuta daude; prokariotoetan ordea aldi berean gertatzen dira.
Ugalketa
[aldatu | aldatu iturburu kodea]Mitosia
[aldatu | aldatu iturburu kodea]Ikuspuntu funtzionaletik ere zelula eukariotoa oso konplexua da. Zelula eukariotoaren funtzioak sakonki aztertuko ditugu ondorengo kapituluetan, hemen bakarrik oinarrizko funtzio bat aipatuko dugu: ugalketa. Zelula eukariotoek prozesu bat asmatu dute gai genetikoa banatzeko: mitosia. Mitosian DNA molekulak paketatzen dira egitura trinkoak (kromosoma mitotikoak, bi kromatidez osatuak) sortzeko. Gero, zitoeskeletoari esker, kromosomak bi multzotan banatzen dira bi zelula alaba genetikoki berdinak sortzeko. Mekanismo honen bidez zelula eukariotoak ugaltzen dira. Hau ugalketa asexuala dugu.
Meiosia
[aldatu | aldatu iturburu kodea]Hala ere eukariotoek beste ugalketa-mota asmatu dute non bi zelula batzen dira zelula alaba genetikoki desberdinak sortzeko: ugalketa sexuala meiosiaren bidez. Fenomeno hau ikuspuntu ebolutibotik oso garrantzitsua izan da izaki eukariotikoen aniztasun harrigarria sortzeko[16][17]. Meiosia zelula diploide bakar batetik lau gameto sortzen dituen zelularen zatiketa mekanismoa da. Gametoek parte hartzen duten ugalketa motari deritzo ere, bertako zelula sexualak meiosi bidez lortzen direlarik, ausaz kromosoma homologoek crossing over eginez eta kromosoma ezberdinak lortuz.
Meiosiaren helburua kromosomen erdiak dituzten zelulak sortaraztea da eta bi zatitan bana daiteke prozesua: lehen zatiketa meiotikoa (I. Meiosia) eta bigarren zatiketa meiotikoa (II. Meiosia). Prozesu meiotikoan zelula diploide batetik (2n) bi zatiketen bidez lau zelula haploide (n) sortzen dira. Lehen zatiketan bi zelula diploide lortzen dira (2n) eta bigarrenean bi zelula diploidetik (2n) lau gameto haploideak (n) agertzen dira.
Eukarioto motak
[aldatu | aldatu iturburu kodea]Landare zelulak
[aldatu | aldatu iturburu kodea]
Zelula eukariotoa deskribatzeko animalia zelula eredutzat hartu egin dugu. Landare-zelularen antolamendua funtsean antzekoa da baina zenbait ezaugarri espezifiko aipa daitezke. Tamainari dagokionez gehienetan landare-zelulak animalia zelulak baino handiagoak dira. Bestalde zelularen egiturari dagokionez landare-zelulek osagai espezifiko batzuk izaten dituzte; garrantzitsuenak zelula horma, bakuolak eta kloroplastoak dira. Zelula horma, mintz plasmatikoaren gainetik dagoen egitura zurruna da, bere osagai nagusia zelulosa izanik (glukosaren polimero bat).
Bakuolak likidoz betetako barrunbeak dira, funtzio askotan inplikatuta daudenak. Bat baino gehiago egon daitezke, baina askotan landare-zelularen zitoplasma osoa bakuola handi batez okupatuta dago. Bakuolaren barruan presio osmotikoa oso altua da eta hori dela eta oso garrantzitsua da zelularen bolumena kontrolatzeko. Horrez gain bakuolak erabiltzen dira hainbat molekula metatzeko eta degradazio-prozesuak burutzeko, hau da, bakuolek jarduera lisosomikoa ere aurkezten dute. Kloroplastoak fotosintesia burutzeko erabiltzen diren organulu zitoplasmatikoak dira.
Egiturari dagokionez kloroplastoak eta mitokondrioak oso antzekoak dira: mintz bikoitz batez mugatuta daude, barruan mintz asko daude, tilakoideak, non konplexu fotosintetikoak kokatuta dauden, eta gainera organuluak bere genoma propioa dauka. Konplexu fotosintetikoetan fotosintesia burutzeko entzima eta eragile guztiak bilduta daude, horien artean klorofila pigmentua. Pigmentu hau ezinbestekoa da argi-energia harrapatzeko eta bere presentziagatik kloroplastoak eta hortaz landare-zelulak ere berde kolorekoak dira. Azpimarratu behar da kloroplastoak ez direla agertzen landare-zelula guztietan, bakarrik fotosintesia gertatzen deneko zeluletan; dena den kloroplastoen antzeko organuluak agertzen dira. Izan ere kloroplastoak organulu-familia handi baten partaideak dira; plastoak izeneko familia hain zuzen.
Eukarioto zelulabakarrak
[aldatu | aldatu iturburu kodea]Izaki eukariotiko asko, prokariotoak bezala, zelula bakar batez osatuta daude, hauek eukarioto zelulabakarrak ditugu. Eukarioto zelulabakarrek osatzen duten taldeak protistoak du izena. Protistoak bakterioak baino askoz konplexuagoak dira, hala ere mikroskopikoak dira, beraz, bakterioak bezala, mikroorganismoak dira.
Protistoek aniztasun harrigarria aurkezte dute. Konplexuenak protozooak dira[18]. Protozooek, zelulabakarrak izan arren, animaliak gogorarazten dizkigute: mugikorrak, oso aktiboak, eta asko harrapakariak dira. Nahiko handiak dira (zelula eukariotio tipiko batekin konparatuz) eta euren anatomia oso konplexua da: oso egitura espezializatuak azaltzen dituzte (flageloak, ile sentikorrak, hanka itxurako apendizeak, aho-egiturak, muskuluen antzeko egitura uzkurkorrak, eta abar).
Diatomeak, beste protisto talde bat, ez dira mugikorrak, fotosintetizatzaileak dira, eta zelula silizez osatutako kutxa batean sartuta dago. Legamiak onddoen antzekoak dira (mugiezinak, ezfotosintetizatzaileak) eta egiturari dagokionez oso sinpleak dira; izan ere legamiak zelula eukariotorik sinpleentzat har ditzakegu. Ezaugarri honengatik hain zuzen, legamiak, eta zehazki Saccharomyces cerevisiae legamia (ogia egiteko erabiltzen dena), oso erabilgarriak suertatu dira zelula eukariotoaren prozesu konplexuak ikertzeko: ziklo zelularraren kontrola esate baterako zelula hauetan agerian jartzea lortu da.
Organismo zelula anitzak
[aldatu | aldatu iturburu kodea]Organismo zelulabakarrez gain, zelula eukariotoa izaki multizelularrak sortzeko gai izan da; izaki hauek eboluzioan zehar gero eta konplexuago bihurtu dira goi-mailako animalia eta landareak eman arte. Elkarte zelularrik sinpleenak koloniak dira. Kolonia batean zelula guztiak oso antzekoak dira; gainera kolonia desantola daiteke eta askatzen diren zelulak indibidualki bizi daitezke. Benetako organismo multizelularrak koloniak baino konplexuagoak dira; elkartzen diren zelulek funtzio espezializatuak betetzen baitituzte.
Bestalde organismo zelulanitza elkarte iraunkorra da: zelulak lotuta mantentzen dira haien artean adhesio-mekanismo espezifikoen bidez. Gainera, animalien kasuan, zelulez gai beste elementu bat oso garrantzitsua da: matrize estrazelularra, zelulen artean dagoen gai azelularra. Zelulak eta matrize estrazelularra modu espezifikoetan antolatzen dira ehunak sortzeko. Ehunak funtsean mota bereko zelulez osatuta daude eta matrize estrazelular espezifiko bat aurkezten dute. Oinarrizko ehuna, animalia guztietan agertzen dena, epitelio-ehuna da; izan ere animalia sinpleenak (koralak esate baterako) bakarrik orri epitelialez daude osatuta. Epitelioak orri zelularrak dira non zelulak modu ordenatu batean kokatuta dauden.
Goi-mailako animalietan ehun-mota asko eta zelula-mota asko daude: ugaztunetan 200 zelula-mota desberdin baino gehiago deskribatu egin dira. Zelula-mota bakoitzak bere funtzio espezifikoa izaten du eta hau bere morfologian islatzen da. Argi dago eukariotoek lortu egin duten konplexutasun-maila harrigarria dela. Hala ere ez dugu ahaztu behar, hain izaki konplexuak izan arren, azken finean zelula bakar batetatik sortuak direla, zelula hori zigotoa da. Hau da, organismo osoa garatzeko informazioa zelula eukarioto bakar batean gordetzen da.
Eboluzioa eta jatorriak
[aldatu | aldatu iturburu kodea]Zelula eukariotoak duela mila miloi urte sortu zirela pentsatzen da[19][20]. Lehenengo izaki eukariotoak zelulabakarrak ziren (protistak). Eukariotoen sailkapena asko aldatu da azken urteetan[21][22].
Fosilak
[aldatu | aldatu iturburu kodea]
Zelula eukariotikoaren jatorria bizitzaren eboluzioaren barruko mugarri bat izan zen, eukariotoak zelula konplexu eta organismo zelulaniztun guztiak barnebiltzen dituztelako. Hala ere, gertakarien kronologia ezartzea oso ariketa zaila da; Knollek 2006an proposatu zuen orian dela 1.600 eta 2.100 milioi urte artean gertatu zela. Akritarko batzuk orain dela 1.650 milio urte jada existitzen ziren, eta baliteke Grypania alga orain dela 2.100 milioi urte bizi izana[24]. Geosiphonen antza duen Diskagma onddo fosila aurkitu da orain dela 2.200 milioi urteko paleozoru batean[25].
Egitura bizidun antolatuak aurkitu dira Gabongo Francevillian B Formazioko eskisto beltz Paleoproterozoikoan, orain dela 2.100 milioi urtekoak. Bizitza eukariotoa garai horretan eboluzionatzea posible da[26]. Gaur egungo taldeekin harreman argia duten fosilak orain dela 1.200 milioi urte agertu ziren, alga gorrien taldekoak, nahiz eta baliteke Vindhya arroan aurkitu diren filamentu batzuk algak izatea. Hala balitz, orain dela 1.600 edo 1.700 milioi urteko fosilak izango lirateke[27].
Biomarkadoreetan oinarrituta oinarrizko eukariotoak lehenago sortu zirela uste da. Australiako eskistoetan aurkitutako ziklopentanoperhidrofenantrenoa dela eta, orain dela 2.700 milioi urte eukariotoak egon zitezkeela proposatu da[28][29].
Arkeobakterioekin harremana
[aldatu | aldatu iturburu kodea]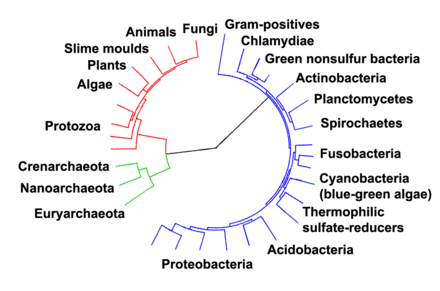

Nukleoko DNA eta eukariotoen makinaria genetikoak antza handiagoa du Arkeobakterioekin bakterioekin baino; horregatik, proposamen batzuetan arkeoak Neomura kladoan batzen dira eukariotoekin. Beste eremu batzuetan, hala nola mintzaren osaketa, eukariotoak bakterioen antzekoak dira. Hiru proposamen daude hau azaltzeko:
- Eukariotoak bi zelula osoren fusioaz sortu ziren, non zitoplasma Eubacterium batetik sortu zen, eta nukleoa arkeo batetik[32], birus batetik[33][34] edo pre-zelula batetik[35][36].
- Eukariotoak Arkaeoatik garatu ziren, eta bakterioen ezaugarriak endosinbiositik eskuratu ziren, bakterio jatorriko proto-mitokondrio batetik[37].
- Eukariotoak eta Archaea modu berezitu batean garatu ziren eubakterio eraldatu batetik.
Beste proposamen alternatiboak ere badaude hau azaltzeko:
- Kronozitoaren hipotesiak esaten du zelula eukariotiko primitibo bat eratu zela arkaea eta bakterio baten endosinbiosiarekin, kronozito deitutako hirugarren zelula baten barruan[38].
- Arbaso komun unibertsal bat dago eukarioto, bakterio eta arkeoen artean, iraungitze masibo bat biziraun zuena eboluzioaren hasierako momentu batean. Eukariotoak eta akariotoak (hau da, bakterio eta arkeoak) ezaugarriak galtzen eboluzionatu zuten[39], eta beraz antzekotasunak jatorrizko ezaugarriak mantentzearen ondorio dira.
Beste inongo talderik ez bada sartzen eukariotoen sorreraren hipotesian, hiru aukera daude bakterio, arkaea eta eukariotoen arteko harremanak emateko, talde monofiletikoak direla pentsatzen badugu. Azpiko taulan batetik hirura arte izendatzen dira. Eozitoaren hipotesia 2. hipotesiaren aldaera bat da, non Archaea parafiletikoa den. Taldeen izenak Harish eta Kurlandek 2017an argitaratutako lanetik hartu dira[40]:
| 1 – Bi inperio | 2 – Hiru domeinu | 3 – Gupta | 4 – Eozito | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
|
|
Erreferentziak
[aldatu | aldatu iturburu kodea]- ↑ M., Youngson, R.. (2006). Collins dictionary of human biology. Collins ISBN 0007221347. PMC 63185739..
- ↑ L.,, Lehninger, Albert. (2005). Lehninger principles of biochemistry. (Fourth edition. argitaraldia) W.H. Freeman ISBN 0716743396. PMC 55476414..
- ↑ Dictionary of life sciences. (2nd ed. argitaraldia) Macmillan 1983 ISBN 0333348672. PMC 10502303..
- ↑ (Ingelesez) «eukaryotic | Origin and meaning of eukaryotic by Online Etymology Dictionary» www.etymonline.com (Noiz kontsultatua: 2018-03-24).
- ↑ Burki, Fabien; Shalchian-Tabrizi, Kamran; Minge, Marianne; Skjaeveland, Asmund; Nikolaev, Sergey I.; Jakobsen, Kjetill S.; Pawlowski, Jan. (2007-08-29). «Phylogenomics reshuffles the eukaryotic supergroups» PloS One 2 (8): e790. doi:. ISSN 1932-6203. PMID 17726520. PMC PMC1949142. (Noiz kontsultatua: 2018-03-24).
- ↑ Parfrey, Laura Wegener; Barbero, Erika; Lasser, Elyse; Dunthorn, Micah; Bhattacharya, Debashish; Patterson, David J.; Katz, Laura A.. (December 2006). «Evaluating support for the current classification of eukaryotic diversity» PLoS genetics 2 (12): e220. doi:. ISSN 1553-7404. PMID 17194223. PMC PMC1713255. (Noiz kontsultatua: 2018-03-24).
- ↑ 1938-2011., Margulis, Lynn,. (1997). What is sex?. Simon & Schuster Editions ISBN 9780684826912. PMC 36798234..
- ↑ (Ingelesez) Zimmer, Carl. (2016-04-11). «Scientists Unveil New ‘Tree of Life’» The New York Times ISSN 0362-4331. (Noiz kontsultatua: 2018-03-24).
- ↑ (Ingelesez) Whitman, W. B.; Coleman, D. C.; Wiebe, W. J.. «Prokaryotes: The unseen majority» Proceedings of the National Academy of Sciences 95 (12): 6578–6583. doi:. (Noiz kontsultatua: 2018-03-24).
- ↑ Tovar J, Fischer A, Clark CG. (1999). «The mitosome, a novel organelle related to mitochondria in the amitochondrial parasite Entamoeba histolytica» Mol. Microbiol. 32 (5): 1013–21. doi:. PMID 10361303..
- ↑ Friedrich, T.; Veenhuis, M.; Huynen, M.A.; Hackstein, J.H. (2005). «An anaerobic mitochondrion that produces hydrogen» Nature 434 (7029): 74–9. doi:. PMID 15744302..
- ↑ Jékely G. (2007). «Origin of eukaryotic endomembranes: a critical evaluation of different model scenarios» Adv. Exp. Med. Biol. 607: 38–51. doi:. PMID 17977457..
- ↑ Martin, William. «Archaebacteria (Archaea) and the origin of the eukaryotic nucleus» Current Opinion in Microbiology 8 (6): 630–637. doi:. (Noiz kontsultatua: 2018-03-24).
- ↑ Takemura M. (2001). «Poxviruses and the origin of the eukaryotic nucleus» J. Mol. Evol. 52 (5): 419–25. doi:. PMID 11443345..
- ↑ Bell PJ. (2001). «Viral eukaryogenesis: was the ancestor of the nucleus a complex DNA virus?» J. Mol. Evol. 53 (3): 251–6. doi:. PMID 11523012..
- ↑ Wächtershäuser G. (2003). «From pre-cells to Eukarya--a tale of two lipids» Mol. Microbiol. 47 (1): 13–22. doi:. PMID 12492850..
- ↑ Wächtershäuser G. (2006). «From volcanic origins of chemoautotrophic life to Bacteria, Archaea and Eukarya» Philos. Trans. R. Soc. Lond. B Biol. Sci. 361 (1474): 1787–1808. doi:. PMID 17008219..
- ↑ Cavalier-Smith T. (2002). «The phagotrophic origin of eukaryotes and phylogenetic classification of Protozoa» Int. J. Syst. Evol. Microbiol. 52 (Pt 2): 297–354. PMID 11931142..
- ↑ (Ingelesez) Knoll, A. H.; Javaux, E. J.; Hewitt, D.; Cohen, P.. (2006-06-29). «Eukaryotic organisms in Proterozoic oceans» Philosophical Transactions of the Royal Society of London B: Biological Sciences 361 (1470): 1023–1038. doi:. ISSN 0962-8436. PMID 16754612. (Noiz kontsultatua: 2018-03-24).
- ↑ Bengtson S, Belivanova V, Rasmussen B, Whitehouse M. (2009). "The controversial "Cambrian" fossils of the Vindhyan are real but more than a billion years older". Proc Natl Acad Sci U S A. 106: 7729–7734 PMID 19416859
- ↑ Brocks JJ, Logan GA, Buick R, Summons RE. (1999). «Archean molecular fossils and the early rise of eukaryotes» Science 285 (5430): 1033–6. doi:. PMID 10446042..
- ↑ Peter Ward. (2008-02-09). «Mass extinctions: the microbes strike back» New Scientist: 40–3..
- ↑ (Ingelesez) Cox, Cymon J.; Foster, Peter G.; Hirt, Robert P.; Harris, Simon R.; Embley, T. Martin. (2008-12-23). «The archaebacterial origin of eukaryotes» Proceedings of the National Academy of Sciences 105 (51): 20356–20361. doi:. ISSN 0027-8424. PMID 19073919. (Noiz kontsultatua: 2018-03-26).
- ↑ (Ingelesez) Knoll, A. H.; Javaux, E. J.; Hewitt, D.; Cohen, P.. (2006-06-29). «Eukaryotic organisms in Proterozoic oceans» Philosophical Transactions of the Royal Society of London B: Biological Sciences 361 (1470): 1023–1038. doi:. ISSN 0962-8436. PMID 16754612. (Noiz kontsultatua: 2018-03-26).
- ↑ Retallack, Gregory J.; Krull, Evelyn S.; Thackray, Glenn D.; Parkinson, Dula. «Problematic urn-shaped fossils from a Paleoproterozoic (2.2Ga) paleosol in South Africa» Precambrian Research 235: 71–87. doi:. (Noiz kontsultatua: 2018-03-26).
- ↑ (Ingelesez) Albani, Abderrazak El; Bengtson, Stefan; Canfield, Donald E.; Bekker, Andrey; Macchiarelli, Roberto; Mazurier, Arnaud; Hammarlund, Emma U.; Boulvais, Philippe et al.. (2010/07). «Large colonial organisms with coordinated growth in oxygenated environments 2.1 Gyr ago» Nature 466 (7302): 100–104. doi:. ISSN 1476-4687. (Noiz kontsultatua: 2018-03-26).
- ↑ (Ingelesez) Bengtson, Stefan; Belivanova, Veneta; Rasmussen, Birger; Whitehouse, Martin. (2009-05-12). «The controversial “Cambrian” fossils of the Vindhyan are real but more than a billion years older» Proceedings of the National Academy of Sciences 106 (19): 7729–7734. doi:. ISSN 0027-8424. PMID 19416859. (Noiz kontsultatua: 2018-03-26).
- ↑ (Ingelesez) Brocks, Jochen J.; Logan, Graham A.; Buick, Roger; Summons, Roger E.. (1999-08-13). «Archean Molecular Fossils and the Early Rise of Eukaryotes» Science 285 (5430): 1033–1036. doi:. ISSN 0036-8075. PMID 10446042. (Noiz kontsultatua: 2018-03-26).
- ↑ (Ingelesez) «Life» New Scientist (Noiz kontsultatua: 2018-03-26).
- ↑ (Ingelesez) Ciccarelli, Francesca D.; Doerks, Tobias; Mering, Christian von; Creevey, Christopher J.; Snel, Berend; Bork, Peer. (2006-03-03). «Toward Automatic Reconstruction of a Highly Resolved Tree of Life» Science 311 (5765): 1283–1287. doi:. ISSN 0036-8075. PMID 16513982. (Noiz kontsultatua: 2018-03-26).
- ↑ (Ingelesez) Egel, Richard. (2012-01-23). «Primal Eukaryogenesis: On the Communal Nature of Precellular States, Ancestral to Modern Life» Life 2 (1): 170–212. doi:. (Noiz kontsultatua: 2018-03-26).
- ↑ Martin, William. «Archaebacteria (Archaea) and the origin of the eukaryotic nucleus» Current Opinion in Microbiology 8 (6): 630–637. doi:. (Noiz kontsultatua: 2018-03-26).
- ↑ (Ingelesez) Takemura, Masaharu. (2001-05-01). «Poxviruses and the Origin of the Eukaryotic Nucleus» Journal of Molecular Evolution 52 (5): 419–425. doi:. ISSN 0022-2844. (Noiz kontsultatua: 2018-03-26).
- ↑ (Ingelesez) Bell, Philip John Livingstone. (2001-09-01). «Viral Eukaryogenesis: Was the Ancestor of the Nucleus a Complex DNA Virus?» Journal of Molecular Evolution 53 (3): 251–256. doi:. ISSN 0022-2844. (Noiz kontsultatua: 2018-03-26).
- ↑ (Ingelesez) Wächtershäuser, G.. (2003-01-01). «From pre-cells to Eukarya - a tale of two lipids» Molecular Microbiology 47 (1): 13–22. doi:. ISSN 1365-2958. (Noiz kontsultatua: 2018-03-26).
- ↑ (Ingelesez) Wächtershäuser, Günter. (2006-10-29). «From volcanic origins of chemoautotrophic life to Bacteria, Archaea and Eukarya» Philosophical Transactions of the Royal Society of London B: Biological Sciences 361 (1474): 1787–1808. doi:. ISSN 0962-8436. PMID 17008219. (Noiz kontsultatua: 2018-03-26).
- ↑ 1967-, Lane, Nick,. The vital question : why is life the way it is?. ISBN 9781781250372. PMC 908425194..
- ↑ (Ingelesez) Hartman, Hyman; Fedorov, Alexei. (2002-02-05). «The origin of the eukaryotic cell: A genomic investigation» Proceedings of the National Academy of Sciences 99 (3): 1420–1425. doi:. ISSN 0027-8424. PMID 11805300. (Noiz kontsultatua: 2018-03-26).
- ↑ Harish, Ajith; Tunlid, Anders; Kurland, Charles G.. «Rooted phylogeny of the three superkingdoms» Biochimie 95 (8): 1593–1604. doi:. (Noiz kontsultatua: 2018-03-26).
- ↑ Harish, Ajith; Kurland, Charles G.. «Akaryotes and Eukaryotes are independent descendants of a universal common ancestor» Biochimie 138: 168–183. doi:. (Noiz kontsultatua: 2018-03-26).
Kanpo estekak
[aldatu | aldatu iturburu kodea]| Autoritate kontrola |
|
|---|