Saccharomyces cerevisiae
| Saccharomyces cerevisiae | ||
|---|---|---|
 | ||
| Taxonomía | ||
| Reino: | Fungi | |
| División: | Ascomycota | |
| Clase: | Saccharomycetes | |
| Orden: | Saccharomycetales | |
| Familia: | Saccharomycetaceae | |
| Género: | Saccharomyces | |
| Especie: |
S. cerevisiae Meyen ex E.C.Hansen | |
La levadura de cerveza (Saccharomyces cerevisiae) Meyen ex E.C.Hansen, de Saccharo azúcar, myces hongo y cerevisiae cerveza es un hongo unicelular, un tipo de levadura utilizado industrialmente en la fabricación de pan, cerveza y vino. El genoma de saccharomyces cerevisiae fue el primero de entre los eucariotas en ser secuenciado. En su ciclo de vida alternan dos formas, una haploide y otra diploide. Ambas formas se reproducen de forma asexual por gemación. En condiciones muy determinadas la forma diploide es capaz de reproducirse sexualmente. En estos casos se produce la meiosis en la célula formándose un asca que contiene cuatro ascosporas haploides.
S. cerevisiae es uno de los modelos más adecuados para el estudio de problemas biológicos. Es un sistema eucariota, con una complejidad solo ligeramente superior a la de la bacteria pero que comparte con ella muchas de sus ventajas técnicas. Además de su rápido crecimiento, la dispersión de las células y la facilidad con que se replican cultivos y aíslan mutantes, destaca por un sencillo y versátil sistema de transformación de ADN. Por otro lado, la ausencia de patogenicidad permite su manipulación con las mínimas precauciones.
S. cerevisiae es un sistema genético que, a diferencia de la mayoría de los otros microorganismos, presenta dos fases biológicas estables: haploide y diploide. La fase haploide permite generar, aislar y caracterizar mutantes con mucha facilidad, mientras que en la diploide se pueden realizar estudios de complementación. Una levadura haploide contiene 16 cromosomas que varían en tamaño de 200 a 2200 kilobases (kb).
Una ventaja adicional de este microorganismo consiste en que se conoce la secuencia completa de su genoma y se mantiene en constante revisión. Ello ha permitido la manipulación genética de los casi 6600 genes que codifica el genoma de levadura, el uso extensivo de micromatrices de ADN para investigar el transcriptoma y estudios a escala genómica de, entre otros muchos aspectos, la expresión génica, localización de proteínas y la organización funcional del genoma y el proteoma.
La maquinaria molecular de muchos procesos celulares se encuentra conservada tanto en levaduras como en plantas y en mamíferos. Esto se ilustra con el hecho de que rutinariamente se han introducido genes de eucariotas superiores en levaduras para el análisis sistemático de su función.
Por estas razones, S. cerevisiae se ha convertido en una importante herramienta a gran escala de análisis de genómica funcional, proporcionando un punto de partida para el análisis de organismos eucariotas más complejos. Al ser un organismo unicelular con una tasa de crecimiento rápida, la levadura se puede utilizar para los estudios de células que resultarían muy complicados o costosos en organismos multicelulares.
Utilidad
[editar]Las utilidades industriales más importantes de esta levadura son la producción de cerveza, pan, vino y kumis, gracias a su capacidad de generar dióxido de carbono y etanol durante el proceso de fermentación. Básicamente este proceso se lleva a cabo cuando esta levadura se encuentra en un medio muy rico en azúcares (como la D-glucosa). En condiciones de escasez de nutrientes, la levadura utiliza otras rutas metabólicas que le permiten obtener un mayor rendimiento energético, y por tanto no realiza la fermentación.
Desde el punto de vista científico, este microorganismo se ha empleado como modelo simple de la célula eucariota. Esto se debe a una serie de ventajas como su facilidad de cultivo y su velocidad de división celular (aproximadamente dos horas).
Nutrición de S. cerevisiae
[editar]
Las fuentes de carbono utilizadas por las levaduras varían desde los carbohidratos hasta los aminoácidos. Además, la capacidad de utilizar ciertos tipos de azúcares ha sido tradicionalmente empleada para la caracterización de las distintas razas que esta especie presenta. Entre los azúcares que puede utilizar están monosacáridos como la glucosa, fructosa, y manosa, entre otros. También son capaces de utilizar disacáridos como la maltosa y la sacarosa y trisacáridos como la rafinosa. Uno de los azúcares que no puede metabolizar es la lactosa, utilizándose este azúcar para distinguir esta especie de Kluyveromyces lactis . También es capaz de utilizar otras fuentes de carbono distintas a carbohidratos y aminoácidos. Entre las más destacadas se encuentra la capacidad de utilizar tanto etanol como glicerol. Por norma general, las levaduras mantienen dos tipos de metabolismo muy bien diferenciados. Por una parte, en condiciones en las que existen altas concentraciones de glucosa, fructosa o maltosa, la tendencia es a realizar una fermentación alcohólica de estos, es decir, se realiza la glucólisis y posteriormente se forma etanol. Una vez que estos azúcares escasean, se produce la respiración del etanol, vía ciclo de Krebs. Evolutivamente esto es un proceso que, a priori, no es ventajoso por ser energéticamente desfavorable para la reproducción del organismo, dado que se obtiene mucha menos energía en el primer proceso que en el segundo. No obstante, la gran mayoría de los organismos son muy sensibles al etanol, por lo que se ha entendido como un proceso de competencia por sustrato. Las levaduras, además de necesitar una fuente de carbono, necesitan tanto fuentes de nitrógeno —como podrían ser el amonio, la urea o distintos tipos de aminoácidos— como fuentes de fósforo. Además, son necesarias vitaminas como la Biotina, también llamada Vitamina H, y distintos elementos traza
Apareamiento en levaduras
[editar]El apareamiento sexual de las levaduras solo puede ocurrir entre células haploides de distinto sexo. Se definen por tanto dos tipos sexuales de levaduras, las células a y las células alfa. En el caso de las levaduras, la determinación sexual no se debe a un cromosoma distinto entre sexos sino más bien a una diferencia en un único locus. Dicho locus se conoce con el nombre de MAT y gobierna el comportamiento sexual entre células haploides y células diploides.
Ciclo sexual de Saccharomyces cerevisiae
[editar]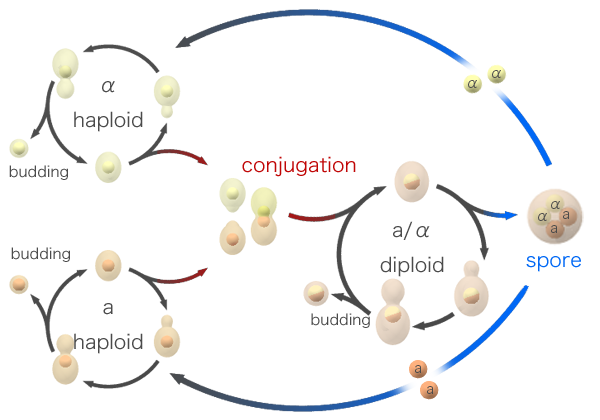
Las levaduras pueden ser haploides o diploides según el estado del ciclo. No obstante, ambos tipos celulares son estables y se pueden reproducir de forma asexual mediante mitosis. La división es por gemación, es decir, las células hijas son de tamaño inferior al de las células madre. Como ya se ha comentado antes, solo las células haploides se pueden reproducir sexualmente, por lo que si una célula de tipo a se encuentra con una célula de tipo α se fusionarán en una sola célula, la cual también sufrirá una fusión de núcleos, formándose un diploide estable que también es capaz de reproducirse de forma asexual. Cuando las condiciones exteriores son desfavorables para las células diploides, sobreviene la meiosis, que provocará la aparición de cuatro esporas haploides, dos de las cuales serán de tipo sexual a y las otras dos de tipo sexual α.
Diferencias entre células a y α
[editar]Las células a producen el "Factor a", que es una feromona peptídica que indica la presencia de células de ese mismo tipo a células del sexo opuesto. Las células a no responderán en ningún caso al factor a, pero sí lo harán si en las inmediaciones existe Factor α. Este tipo de respuesta desencadena la formación de una protuberancia en las células hacia la fuente de las feromonas de sexo contrario y es recíproco. En la actualidad se conocen las bases moleculares que rigen este comportamiento, el cual se debe a la transcripción o represión de genes en los dos tipos sexuales de levaduras. Las células a transcriben los genes que producirán el factor a, además de un receptor de membrana que se conoce con el nombre de Ste2p. Dicho receptor es capaz de unirse al factor α y desencadenar una serie de señales intracelulares mediadas por la proteína G. Además, las células a reprimen la expresión de los genes que formarán las proteínas necesarias para la síntesis del factor α y el receptor de membrana Ste3p. En las células α ocurre exactamente lo contrario a lo descrito. Todas estas diferencias entre activación y represión transcripcional son causadas por la presencia de uno de los dos alelos de un locus denominado MAT: MATa o Matα. El alelo Mata codifica para una única proteína denominada a1. El alelo Matα codifica para α1 y α2, que en los haploides dirigen la transcripción del programa específico de las células α.
Diferencias entre células haploides y diploides
[editar]Las células haploides de cualquiera de los sexos responde a la feromona producida por el sexo contrario. Las células de sexo opuesto podrán fusionarse, formando una célula diploide. Las células haploides nunca podrán realizar la meiosis en condiciones normales. Por el contrario, las células diploides no producen ni responden a ninguno de los dos tipos de feromonas, pero sí pueden realizar meiosis bajo condiciones ambientales muy determinadas. Al igual que existen patrones de expresión génica entre células a y α, también existen diferencias entre la expresión génica entre células haploides y diploides. Un ejemplo de esto último es el caso de la endonucleasa HO, que es expresada en las células haploides, o el caso de IME1, cuya expresión está reprimida en los diploides. Las diferencias entre los patrones de expresión entre haploides y diploides son producidas por el locus MAT. Las células haploides solo contienen una copia del locus MAT, en cualquiera de sus variantes alélicas, y esta determinará el sexo de la célula. Los diploides resultan de la fusión celular entre células de distinto sexo, por lo que presentan los dos loci.
Cambio sexual en levaduras
[editar]Una levadura haploide es capaz de cambiar de sexo. De tal forma que si una única célula de tipo a o α está en un medio sin la presencia del sexo contrario, al cabo de unas cuantas generaciones se advierte la presencia de la feromona contraria y un incremento en células diploides. Esta aparición de diploides puede ser tan alta que desplaza la población de haploides, ya que esta última población tiene una alta tendencia a aparearse. Las cepas de levaduras utilizadas en los laboratorios no suelen realizar este cambio de sexo debido a que están alteradas en el gen HO, que es determinante para el cambio de sexo. Esto genera una propagación estable de cualquiera de los tipos celulares de los haploides, y nunca se llegan a formar diploides, en condiciones normales.
HML y HMR
[editar]
¿Cómo pueden cambiar las levaduras de sexo si este fenotipo viene dado por un único locus MAT? La respuesta es simple: las levaduras poseen copias del locus MAT que están silenciadas y por tanto no interfieren en la determinación sexual. Cuando se produce un cambio en el sexo de las levaduras, se produce un reemplazamiento génico del locus MAT por una de las copias adicionales. Las copias silenciosas se denominan HML (que generalmente llevan una copia silenciosa del alelo MATα) y HMR (que generalmente lleva una copia silenciosa del alelo MATa). Ambos loci se encuentran en el cromosoma III y están situados a derecha (HMR, donde la R es de right) y a izquierda (HML, donde la L es de left) del locus MAT en cualquiera de sus variantes alélicas.
Mecanismo del cambio sexual
[editar]El proceso de cambio sexual en las levaduras viene dado por la conversión génica iniciada por la endonucleasa HO. La expresión de dicha endonucleasa está regulada específicamente en los haploides y solo es activa en las células haploides durante la fase de ciclo celular G1. La endonucleasa HO genera un corte específico en el ADN del locus MAT. Una vez se realiza el corte, los extremos libres generados son atacados por exonucleasas, produciéndose la degradación del locus MAT en ambos sentidos. Esta ausencia de parte de un locus genera la activación de sistemas de reparación del ADN que conllevan el reemplazamiento del locus ausente por una de las copias adicionales HMR o HML.
Direccionalidad del cambio sexual
[editar]La reparación del locus MAT cortado por la endonucleasa HO permite el cambio sexual, ya que, por norma general, se reemplaza por el alelo contrario al que estaba en un principio. De esta forma cuando una célula a decide realizar un cambio sexual, el alelo MATa es degradado y reemplazado por la copia HML. Esto da como resultado el cese de la expresión del antiguo MATa y el inicio de la expresión del nuevo MATα, con todo lo que esto conlleva. Esta direccionalidad es en parte debida a la actividad de un enhancer de recombinación (RE)[1] ubicado en el brazo izquierdo del cromosoma III. La eliminación de esta región provoca que una célula a repare incorrectamente el locus MAT usando la región HMR. En las células a la proteína Mcm1 se une a RE y promueve la recombinación de la región HML. En células α, en cambio, el factor α2 se une a la región RE y reprime la recombinación. Existiría un sesgo innato por defecto hacia la recombinación con HMR que promovería la recombianción con esa región en este último caso. Estos mecanismos se encuentran actualmente bajo investigación
Genoma de S. cerevisiae
[editar]El genoma de esta levadura contiene aproximadamente 12.156.677 pares de bases (12 Mb) con 6.275 marcos abiertos de lectura, o genes, de los que se cree que solo 5.800 son genes realmente funcionales. Está organizado en un conjunto de dieciséis cromosomas completamente caracterizados con tamaños entre 200 a 2200 kb. Se estima que comparte aproximadamente el 23 % del genoma con el ser humano.
Patogenia
[editar]Saccharomyces cerevisiae no se considera un patógeno común. Actualmente cobra importancia su papel oportunista en sepsis en enfermos de leucemia y otras infecciones oportunistas en enfermos de sida. Ha sido reportado recientemente como causante del Auto-brewery syndrome o Síndrome de fermentación Intestinal.[2]
Se han encontrado anticuerpos frente a S. cerevisiae en el 60-70 % de los afectados de enfermedad de Crohn y en el 10-15 % de los enfermos de colitis ulcerativa (y en el 8 % de personas sanas).[3]
Véase también
[editar]Referencias
[editar]- ↑ «The Saccharomyces cerevisiae recombination enhancer biases recombination during interchromosomal mating-type switching but not in interchromosomal homologous recombination». Genetics 166 (3): 1187-97. March 2004. PMC 1470794. PMID 15082540. doi:10.1534/genetics.166.3.1187.
- ↑ Cristine Souza Goebel, y otros (2013). «Infección por Saccharomyces cerevisiae». Revista Iberoamericana de Micología.
- ↑ Walker, L. J.; Aldhous, M. C.; Drummond, H. E.; Smith, B. R. K.; Nimmo, E. R.; Arnott, I. D. R.; Satsangi, J. (2004). «Anti-Saccharomyces cerevisiae antibodies (ASCA) in Crohn's disease are associated with disease severity but not NOD2/CARD15 mutations». Clinical and Experimental Immunology 135 (3): 490-6. PMC 1808965. PMID 15008984. doi:10.1111/j.1365-2249.2003.02392.x.
Bibliografía
[editar]- Lewin B (2001). Genes VII. Marban
- Cordell, B.; McCarthy, J. (2013). "A Case Study of Gut Fermentation Syndrome (Auto-Brewery) with Saccharomyces cerevisiae as the Causative Organism". International Journal of Clinical Medicine 04 (7): 309. doi:10.4236/ijcm.2013.47054.
Enlaces externos
[editar]Wikimedia Commons alberga una galería multimedia sobre Saccharomyces cerevisiae.
| Control de autoridades |
|
|---|
Datos: Q719725
Especies: Saccharomyces cerevisiae