Nucli cel·lular

El nucli (del llatí nucleus o nuculeus, 'núcula') és un orgànul envoltat per una membrana que existeix únicament en les cèl·lules eucariotes. En general, aquestes cèl·lules tenen un sol nucli, però n'hi ha algunes, com els eritròcits dels mamífers, que no en tenen cap, i d'altres, com els osteoclasts, que en tenen molts. Les principals estructures que formen el nucli són l'embolcall nuclear, una doble membrana que envolta l'orgànul sencer i en separa els continguts del citoplasma cel·lular, i la matriu nuclear (incloent-hi la làmina nuclear), una xarxa situada dins el nucli que li ofereix suport mecànic de manera similar a com el citoesquelet suporta la cèl·lula en el seu conjunt.
El nucli conté gairebé tot el genoma cel·lular, arranjat en cromosomes, cadenes llargues d'ADN integrat amb diverses proteïnes, com ara histones, que el protegeixen i l'estructuren. Els gens d'aquests cromosomes estan organitzats per afavorir el funcionament cel·lular. Com a centre neuràlgic de la cèl·lula, el nucli manté la integritat dels gens i regula l'expressió gènica per controlar les activitats de la cèl·lula.
Com que l'embolcall nuclear no permet el pas a la majoria de molècules, calen porus nuclears per regular el transport nuclear de molècules. Aquests porus travessen ambdues membranes, oferint un canal que permet el lliure moviment de molècules petites i ions. El moviment de molècules més grans, com ara proteïnes, està ben controlat, i requereix transport actiu regulat per proteïnes portadores. El transport nuclear és essencial pel funcionament cel·lular, car el moviment a través dels porus és necessari tant per l'expressió gènica com pel manteniment dels cromosomes.
Tot i que l'interior del nucli no conté subcompartiments delimitats per membranes, els seus continguts no són uniformes, i existeixen una sèrie de «cossos subnuclears», formats per proteïnes úniques, molècules d'ARN i determinades parts dels cromosomes. El més conegut d'aquests cossos és el nuclèol, el paper principal del qual és en l'assemblatge dels ribosomes. Els ribosomes, després de ser produïts al nuclèol, són exportats al citoplasma que és on tradueixen l'ARNm.
Història
[modifica]

El nucli fou el primer orgànul a ser descobert. La il·lustració probablement més antiga conservada es remunta a Antonie van Leeuwenhoek, un dels primers microscopistes (1632–1723). Observà un «lumen» al nucli, als glòbuls vermells del salmó.[1] A diferència dels glòbuls vermells dels mamífers, els dels altres vertebrats conserven el nucli. El nucli també fou descrit per Franz Bauer el 1804[2] i amb més detall el 1831, pel botànic escocès Robert Brown en una xerrada a la Societat Linneana de Londres. Brown estava estudiant orquídies amb el microscopi quan observà una àrea opaca, que anomenà arèola o nucli, a les cèl·lules de la capa exterior de la flor.[3] No suggerí cap funció potencial per aquesta estructura. El 1838, Matthias Schleiden suggerí que el nucli té un paper en la generació de cèl·lules, i encunyà el nom "citoblast" ("formador de cèl·lules"). Creia haver observat noves cèl·lules formant-se al voltant dels "citoblasts". Franz Meyen fou un ferm oponent d'aquest punt de vista; descrigué la multiplicació de les cèl·lules per divisió i creia que moltes cèl·lules mancaven de nucli. La idea que les cèl·lules es poden generar de novo, sigui pel "citoblast" o d'una altra manera, contradeia el treball de Robert Remak (1852) i Rudolf Virchow (1855), que propagaren de manera decisiva el nou paradigma que les cèl·lules només són generades per cèl·lules (omnis cellula e cellula). La funció del nucli romania incerta.[4]
Entre el 1876 i el 1878 Oscar Hertwig publicà diversos estudis sobre la fertilització dels ous d'eriçó de mar, demostrant que el nucli de l'esperma penetra a l'oòcit i es fusiona amb el seu nucli. Aquesta fou la primera vegada que se suggerí que un individu es desenvolupa a partir d'una (única) cèl·lula nucleada. Això contradeia la teoria d'Ernst Haeckel que afirmava que durant el desenvolupament embrionari es repeteix el procés complet de filogènesi d'una espècie, incloent-hi la generació de la primera cèl·lula nucleada a partir d'una "monèrula", una massa sense estructura de mucositat primordial (Urschleim). Per consegüent, la necessitat del nucli de l'esperma per la fertilització fou discutida durant bastant de temps. Tanmateix, Hertwig confirmà la seva observació en altres grups d'animals, com ara amfibis i mol·luscs. El 1884, Eduard Strasburger obtingué els mateixos resultats amb les plantes. Això obrí el camí per assignar al nucli un paper important en l'herència. El 1873, August Weismann postulà l'equivalència de les "cèl·lules" germinals materna i paterna per l'herència. La funció del nucli com a portador de la informació genètica no es revelà fins més tard, després del descobriment de la mitosi i del redescobriment de les lleis de Mendel a principis del segle xx; es desenvolupà la teoria cromosòmica de l'herència.[4]
Estructures
[modifica]El nucli és l'orgànul cel·lular més gran en els animals.[5] En les cèl·lules mamíferes, el diàmetre mitjà del nucli és d'aproximadament sis micròmetres (μm), ocupant aproximadament un 10% del volum total de les cèl·lules.[6] El líquid viscós que conté s'anomena nucleoplasma, i té una composició similar a la del citosol que es troba a l'exterior del nucli.[7] Apareix com un orgànul dens i més o menys esfèric.
Embolcall i porus nuclears
[modifica] |
L'embolcall nuclear, també conegut com a membrana nuclear, es compon de dues membranes cel·lulars, una d'interior i una d'exterior, situades en paral·lel i separades per entre 10 i 50 nanòmetres (nm). L'embolcall nuclear envolta completament el nucli i separa el material genètic de la cèl·lula del citoplasma que l'envolta, fent de barrera per evitar que les macromolècules es difonguin lliurement entre el nucleoplasma i el citoplasma.[8] La membrana nuclear exterior és contínua amb la membrana del reticle endoplasmàtic rugós (RER) i també està plena de ribosomes. L'espai entre les membranes rep el nom d'espai perinuclear i és continu amb el lumen del RER.
Els porus nuclears, que ofereixen canals aquosos a través de l'embolcall, es componen de múltiples proteïnes, denominades col·lectivament nucleoporines. Els porus tenen un pes molecular d'uns 125 milions de daltons i es componen d'entre 50 (en els llevats) i 100 proteïnes (en els vertebrats).[5] Els porus tenen un diàmetre total de 100 nm; tanmateix, l'espai a través del qual es difonen lliurement les molècules té una amplada de només uns 9 nm, a causa de la presència de sistemes reguladors al centre del porus. Aquesta mida permet que passin lliurement les petites molècules solubles en aigua, tot evitant que les molècules més grans, com ara els àcids nucleics o les proteïnes més grans, entrin o surtin del nucli de manera inapropiada. Aquestes molècules grans han de ser transportades activament al nucli. El nucli d'una cèl·lula mamífera típica té entre 3.000 i 4.000 porus a l'embolcall,[9] cadascun dels quals té una estructura en forma d'anell octasimètrica a la posició en què es fusionen la membrana interior i l'exterior.[10] Unida a l'anell, hi ha una estructura anomenada "cistell nuclear" que s'estén dins el nucleoplasma, així com una sèrie d'extensions filamentoses que penetren al citoplasma. Ambdues estructures serveixen per mitjançar la unió a proteïnes de transport nuclears.[5]
La majoria de proteïnes, subunitats ribosòmiques i alguns ARN són transportats a través dels complexos de porus en un procés mitjançat per una família de factors de transport coneguts com a carioferines. Les carioferines que mitjancen el moviment vers el nucli també reben el nom d'importines, mentre que les que mitjancen el moviment des del nucli cap enfora s'anomenen exportines. La majoria de carioferines interaccionen directament amb el que porten, tot i que algunes utilitzen proteïnes adaptadores.[11] Les hormones esteroidals com ara el cortisol i l'aldosterona, així com altres petites molècules solubles en lípids implicades en la senyalització intercel·lular, es poden difondre a través de la membrana cel·lular i cap al citoplasma, on s'uneixen a proteïnes receptores nuclears que són transportades al nucli. Allà compleixen la funció de factors de transcripció quan estan unides al seu lligand; en absència del lligand, molts d'aquests receptors funcionen com a histona desacetilases que inhibeixen l'expressió gènica.[5]
Làmina nuclear
[modifica]En les cèl·lules animals, dues xarxes de filaments intermedis proporcionen suport mecànic al nucli: la làmina nuclear forma una malla organitzada a la cara interna de l'embolcall, mentre que hi ha un suport menys organitzat a la cara citosòlica de l'embolcall. Ambdós sistemes proporcionen suport estructural per l'embolcall nuclear i llocs d'ancoratge pels cromosomes i els porus nuclears.[6]
La làmina nuclear es compon majoritàriament de proteïnes anomenades làmines. Com totes les proteïnes, les làmines són sintetitzades al citoplasma i després transportades a l'interior del nucli, on són assemblades abans d'incorporar-se a la xarxa existent de la làmina nuclear.[12][13] Les lamines, també es troben al nucleoplasma, on formen una altra estructura regular coneguda com a "vel nucleoplàsmatic",[14] que és visible al microscopi de fluorescència. La funció exacta del vel és incerta, tot i que és exclòs del nuclèol i és present durant la interfase.[15] Les estructures de làmina que formen el vel s'uneixen a la cromatina, i la disrupció de la seva estructura inhibeix la transcripció dels gens codificadors de proteïnes.[16]
Com els components dels altres filaments intermedis, el monòmer de les làmines conté un domini alfa helicoidal utilitzat per dos monòmers per enrotllar-se l'un al voltant de l'altre, formant una estructura dimèrica anomenada hèlix superenrotllada. Aleshores dues d'aquestes estructures s'uneixen de costat, en una configuració antiparal·lela, per formar un tetràmer anomenat "protofilament". Vuit d'aquests protofilaments formen una configuració lateral que es caragola per formar un "filament" semblant a una corda. Aquests filaments es poden assemblar i desassemblar de manera dinàmica, cosa que significa que els canvis de la longitud del filament depenen de les velocitats competidores d'addició i sostracció de protofilaments.[6]
Les mutacions en els gens de les làmines que causen defectes en l'assemblatge dels filaments són conegudes com a "laminopaties". La laminopati més destacada és la família de malalties conegudes com a progèria, que provoca un envelliment prematur en els malalts. El mecanisme exacte pel qual els canvis bioquímics associats donen origen al fenotip envellit no està ben conegut.[17]
Cromosomes
[modifica]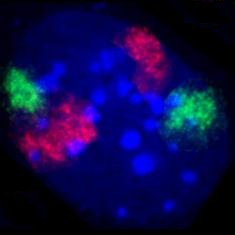
El nucli cel·lular conté la majoria del material genètic de la cèl·lula, en forma de múltiples molècules linears d'ADN organitzades en estructures anomenades cromosomes. Durant la majoria del cicle cel·lular, estan organitzades en un complex ADN-proteïna anomenat cromatina, i durant la divisió cel·lular es pot veure com la cromatina forma els cromosomes ben definits típic d'un cariotip. Una petita part dels gens de la cèl·lula es troben als mitocondris.
Hi ha dos tipus de cromatina. L'eucromatina és la forma menys compacta d'ADN, i conté gens que són expressats sovint per la cèl·lula.[18] L'altre tipus, l'heterocromatina, és la forma més compacta, i conté ADN que no es transcriu gaire sovint. Aquesta estructura se subdivideix en heterocromatina "facultativa", que consisteix en gens que només s'organitzen en heterocromatina en determinats tipus de cèl·lules o determinades fases del desenvolupament, i heterocromatina "constitutiva", que consisteix en components estructurals del cromosoma, com ara telòmers i centròmers.[19] Durant la interfase, la cromatina s'organitza en parts individuals distintes,[20] anomenades "territoris cromosòmics".[21] Els gens actius, que generalment es troben a la regió eucromàtica del cromosoma, tendeixen a situar-se vers el límit del territori cromosòmic.[22]
S'han associat anticossos contra determinats tipus d'organització de la cromatina, especialment els nucleosomes, amb una sèrie de malalties autoimmunitàries, com ara el lupus eritematós sistèmic.[23] Se'ls coneix com a anticossos antinuclears (ANA) i també se'ls ha observat en combinació amb l'esclerosi múltiple, com a part d'una disfunció general del sistema immunitari.[24] Com en el cas de la progèria, el paper que tenen els anticossos a l'hora d'induir els símptomes de les malalties autoimmunitàries no és evident.
Nuclèol
[modifica]El nuclèol és una estructura distinta amb una tinció densa que es troba al nucli. No està envoltat per cap membrana, i a vegades es diu que és un «suborgànul». Es forma al voltant de repeticions en tàndem d'ADNr, l'ADN que codifica l'ARN ribosòmic (ARNr). Aquestes regions són denominades regions organitzants nucleolars (NOR). Les funcions principals del nuclèol són sintetitzar ARNr i assemblar ribosomes. La cohesió estructural del nuclèol depèn de la seva activitat, car l'asemblatge de ribosomes al nucleòl resulta en l'associació passatgera de components nucleolars, facilitant més assemblatges de cromosomes i, per tant, més associació. Aquest model és recolzat per les observacions que indiquen que la desactivació de l'ADNr fa que es barregin les estructures nucleolars.[25]
El primer pas de l'assemblatge ribosòmic és la transcripció de l'ADNr per part d'una proteïna anomenada ARN polimerasa I, formant un gran precursor pre-ARNr. Aquest precursor és fragmentat en les subunitats ARNr 5.8S, 18S i 28S.[26] La transcripció, el processament posttranscripcional i l'assemblatge de l'ARNr es produeix al nuclèol, amb l'ajut de petites molècules d'ARN nucleolar petit (ARNnop), algunes de les quals deriven d'introns empalmats provinents de gens codificants d'ARN missatgers relacionats amb el funcionament ribosòmic. Les subunitats ribosòmiques assemblades són les estructures més grans que passen pels porus nuclears.[5]
En observar-lo al microscopi electrònic, es pot veure que el nuclèol es compon de tres regions distingibles: a la part més interior, els «centres fibril·lars» (CF), envoltats pel «component fibril·lar dens» (CFD), que al seu torn està envoltat pel «component granular» (CG). La transcripció de l'ADNr es fa o bé als CF o al límit CF-CFD; per això, quan s'incrementa la transcripció d'ADNr a la cèl·lula, es detecten més CF. La majoria de la fragmentació i modificació dels ARNr es fa al CFD, mentre que els passos posteriors que impliquen l'assemblatge de les proteïnes a les unitats ribosòmiques es fan al CG.[26]
Altres cossos subnuclears
[modifica]| Nom de l'estructura | Diàmetre de l'estructura |
|---|---|
| Cossos de Cajal | 0,2–2,0 µm[27] |
| PIKA | 5 µm[28] |
| Cossos PML | 0,2–1,0 µm[29] |
| Paraspeckles | 0,2–1,0 µm[30] |
| Speckles | 20–25 nm[28] |
A part del nuclèol, el nucli conté una sèrie d'altres cossos no delimitats per membranes. Aquests cossos inclouen els cossos de Cajal, gèminis de cossos enrotllats, associacions cariosòmiques d'interfase polimòrfica (PIKA), cossos de leucèmia promielocítica (PML), paraspeckles i speckles d'empalmament. Tot i que se sap poca cosa sobre alguns d'aquests dominis, són significants pel fet que demostren que el nucleoplasma no és una mescla uniforme, sinó que conté subdominis funcionals organitzats.[29]
Altres estructures subnuclears apareixen com a part de processos patològics anormals. Per exemple, s'ha observat la presència de petites barres intranuclears en alguns casos de miopatia nemalínica. Aquest trastorn sol ser el resultat de mutacions en l'actina, i les barres en si es componen d'actina mutant, a més d'altres proteïnes citoesquelètiques.[31]
Cossos de Cajal i gemmes
[modifica]Un nucli típicament conté entre una i deu estructures compactes anomenades cossos de Cajal o cossos enrotllats (CB), amb un diàmetre d'entre 0,2 µm i 2,0 µm, segons el tipus de cèl·lula i l'espècie.[27] Quan se'ls observa al microscopi electrònic, semblen cabdells de fils embolicats,[28] i són focus densos de distribució de la proteïna coilina.[32] Els CB tenen una sèrie de papers diferents relacionats amb el processament de l'ARN, específicament l'ARN nucleolar petit (ARNnop) i la maduració de l'ARN nuclear petit (ARN) i la modificació de l'ARNm de les histones.[27]
Similars als cossos de Cajal són els gèminis de cossos enrotllats, o gemmes, el nom dels quals deriva de la constel·lació de Gèminis, en referència a la seva estreta relació de «bessons» amb els CB. Les gemmes tenen una mida i forma similars a les dels CB, i de fet són pràcticament indiferenciables al microscopi.[32] A diferència dels CB, les gemmes no contenen ribonucleoproteïnes nuclears petites (RNPnp), però sí que contenen una proteïna anomenada "supervivent de les neurones motrius" (SNM), la funció de la qual està relacionada amb la biogènesi de les RNPnp. Es creu que les gemmes ajuden els CB en la biogènesi d'RNPnp,[33] tot i que també s'ha suggerit, basant-se en proves de microscòpia, que els CB i les gemmes són manifestacions diferents de la mateixa estructura.[32]
Dominis PIKA i PTF
[modifica]Els dominis PIKA, o associacions cariosòmiques d'interfase polimòrfiques, foren descrites per primer cop en estudis de microscòpia l'any 1991. La seva funció era i roman incerta, tot i que no es creia que estiguessin associades a la replicació o transcripció de l'ADN actives ni al processament de l'ARN.[34] S'ha observat que sovint s'associen a dominis diferenciats definits per una localització densa del factor de transcripció PTF, que promou la transcripció de l'ARNnp.[35]
Cossos PML
[modifica]Els cossos de leucèmia promielocítica (cossos PML) són cossos esfèrics que es troben dispersos arreu del citoplasma, amb una mida de 0,2–1,0 µm. Se'ls coneix amb una sèrie de noms diferents, incloent-hi domini nuclear 10 (ND10), cossos de Kremer i dominis oncogènics PML. Sovint se'ls observa al nucli en associació amb els cossos de Cajal i cossos d'excisió. S'ha suggerit que tenen un paper en la regulació de la transcripció.[29]
Paraspeckles
[modifica]Els paraspeckles, descoberts per Fox et al. el 2002, són compartiments de forma irregular que es troben a l'espai intercromatínic del nucli.[36] Foren observats originalment en cèl·lules HeLa, on generalment n'hi ha 10–30 per nucli,[37] però actualment se sap que els paraspeckles existeixen en totes les cèl·lules primàries humanes, en els llinatges de cèl·lules transformades i les seccions de teixits.[38] El seu nom deriva de la seva distribució al nucli: para és una abreviació de "paral·lel", mentre que speckles ("XXX", en anglès), es refereix als speckles d'excisió, als quals sempre estan pròxims.[37]
Els paraspeckles són estructures dinàmiques que s'alteren en resposta a canvis en l'activitat metabòlica cel·lular. Són dependents de la transcripció[36] i en absència de la transcripció d'ARN Pol II, el paraspeckle desapareix i tots els seus components proteics associats (PSP1, p54nrb, PSP2, CFI(m)68 i PSF) formen un tap en forma de creixent al nuclèol. Aquest fenomen s'observa durant el cicle cel·lular. Al cicle cel·lular, els paraspeckles estan presents durant la interfase i tota la mitosi, tret de la telofase. Durant telofase, quan es formen els dos nuclis fills, no hi ha transcripció d'ARN Pol II, de manera que els components proteics formen un tap perinucleolar.[38]
Speckles d'empalmament
[modifica]També coneguts com a "clústers de grànuls d'intercromatina" o "compartiments de factor d'empalmament", els speckles són rics en PRNnp d'empalmament i altres proteïnes d'empalmament necessàries per al preprocessament de l'ARNm.[39] A causa de les necessitats canviants de la cèl·lula, la composició i situació d'aquests cossos canvia segons la transcripció d'ARNm i la regulació per fosforilació de diverses proteïnes.[40]
Funcions
[modifica]La funció principal del nucli cel·lular és controlar l'expressió gènica i mitjançar la replicació de l'ADN durant el cicle cel·lular. El nucli proporciona un lloc per la transcripció segregat de la ubicació de la traducció al citoplasma, permetent nivells de regulació gènica que no són possibles pels procariotes.
Compartimentalització cel·lular
[modifica]L'embolcall nuclear permet que el nucli controli els seus continguts, i els separa de la resta del citoplasma quan és necessari. Això és important per controlar processos a banda i banda de la membrana cel·lular. En els casos en què cal restringir un procés citoplasmàtic, se'n mou un participant clau al nucli, on interacciona amb els factors de transcripció per subregular la producció de determinats enzims de la ruta. Aquest mecanisme regulador es dona per la glucòlisi, una ruta cel·lular que descompon la glucosa per produir energia. L'hexocinasa és un enzim que s'encarrega del primer pas de la glucòlisi, formant glucosa-6-fosfat a partir de la glucosa. En concentracions elevades de fructosa-6-fosfat, una molècula que es forma posteriorment a partir de glucosa-6-fosfat, una proteïna reguladora s'enduu l'hexocinasa al nucli,[41] on forma un complex repressor transcripcional amb proteïnes nuclears per reduir l'expressió de gens implicats en la glucòlisi.[42]
Per tal de controlar quins gens es transcriuen, la cèl·lula separa algunes proteïnes de factor de transcripció encarregades de regular l'expressió gènica de l'accés físic a l'ADN, fins que siguin activades per altres rutes de senyalització. Això evita fins i tot els nivells baixos d'expressió gènica inapropiada. Per exemple, en els casos dels gens controlats per NF-κB, que estan implicats en la majoria de respostes inflamatòries, la transcripció és induïda en resposta a una ruta de senyalització, com la que inicia la molècula de senyalització TNF-α, s'uneix a un receptor de la membrana cel·lular, provocant el reclutament de proteïnes de senyalització, i finalment activa el factor de transcripció NF-κB. Un senyal de localització nuclear de la proteïna NF-κB permet que sigui transportada pel porus nuclear cap a dins del nucli, on estimula la transcripció dels gens diana.[6]
La compartimentalització permet a la cèl·lula evitar la traducció d'ARNm no empalmat.[43] L'ARN eucariota conté introns que han de ser eliminats abans de ser traduït per generar proteïnes funcionals. L'empalmament es fa dins el nucli, abans que els ribosomes puguin arribar a l'ARNm per traduir-lo. Sense el nucli, els ribosomes traduirien ARNm acabat de produir (no processat), creant proteïnes deformes i no funcionals.
Expressió gènica
[modifica]L'expressió gènica implica en primer lloc la transcripció, en què es fa servir l'ADN com plantilla per produir ARN. En el cas dels gens que codifiquen proteïnes, l'ARN produït a partir d'aquest procés és ARN missatger (ARNm), que després ha de ser traduït pels ribosomes per formar una proteïna. Com que els ribosomes es troben a l'exterior del nucli, cal exportar l'ARNm.[44]
Com que el nucli és el lloc on es produeix la transcripció, també conté una sèrie de proteïnes que o bé mitjancen directament la transripció, o bé tenen un paper en la regulació del procés. Aquestes proteïnes inclouen les helicases que desenrotllen la molècula bicatenària d'ADN per facilitar-hi l'accés, les ARN polimerases que sintetitzen la molècula d'ARN en creixement, les topoisomerases que canvien la quantitat de superenrotllament de l'ADN, ajudant-lo a enrotllar-se i desenrotllar-se, i una gran varietat de factors de transcripció que regulen l'expressió.[45]
Processament de pre-ARNm
[modifica]Les molècules d'ARN acabades de sintetitzar són conegudes com a transcrits primaris o pre-ARNm. Han d'experimentar modificació posttranscripcional al nucli abans de ser exportats al citoplasma; l'ARNm que apareix al nucli sense aquestes modificacions és degradat en lloc de ser utilitzat per la traducció. Les tres modificacions principals són el càping 5', la poliadenilació 3', i l'empalmament d'ARN. Dins el nucli, el pre-ARNm està associat a una sèrie de proteïnes en complexos coneguts com a partícules de ribonucleoproteïnes heterogènies (RNPhn). L'afegit del 5' cap és cotranscripcional i és el primer pas de la modificació postranscripcional. La cua de poliadenina 3' només és afegida quan ja s'ha acabat la transcripció.
L'empalmament de l'ARN, dut a terme per un complex anomenat spliceosoma, és el procés pel qual els introns, regions d'ADN que no codifiquen proteïnes, són eliminats del pre-ARNm i els exons restants són connectats per reformar una única molècula contínua. Aquest procés sol produir-se després del càping 5'i la poliadenilació 3' però en els transcriptes amb molts exons pot començar abans que es completi la síntesi.[5] Molts pre-ARNm, incloent-hi els que codifiquen anticossos, poden ser empalmats de múltiples maneres per produir diferents ARNm madurs que codifiquin seqüències proteiques diferents. Aquest procés rep el nom d'empalmament alternatiu i permet produir una gran varietat de proteïnes a partir d'una quantitat limitada d'ADN.
Dinàmica i regulació
[modifica]Transport nuclear
[modifica]
L'entrada i sortida de molècules grans del nucli estan controlades estrictament pels complexos de porus nuclears. Tot i que les molècules petites poden entrar al nucli sense regulació,[46] les macromolècules com ara l'ARN i les proteïnes requereixen carioferines d'associació anomenades importines (per entrar al nucli) o exportines (per sortir-ne). Les proteïnes "carregades" que han de ser mogudes del citoplasma al nucli contenen seqüències d'aminoàcids curtes conegudes com a senyals de localització cel·lular, que s'uneixen a les importines, mentre que les que han de ser mogudes del nucli al citoplasma porten senyals d'exportació nuclear que s'uneixen a les exportines. La capacitat de les importines i les exportines de transportar la seva càrrega és regulada per les GTPases, uns enzims que hidrolitzen la molècula trifosfat de guanosina per alliberar energia La GTPasa clau del transport nuclear és la Ran, que es pot unir o bé al GTP o el GDP (difosfat de guanosina) depenent de si es troba al nucli o el citoplasma. Les importines depenen de la RanGTP per dissociar-se de la seva càrrega, mentre que les exportines la necessiten per unir-s'hi.[11]
La importació nuclear necessita que de la importina s'uneixi a la seva càrrega al citoplasma i la porti al nuclèol a través del porus nuclear. Dins el nuclèol, la RanGTP separa la càrrega de la importina, permetent que aquesta última surti del nucli i sigui reutilitzada. L'exportació nuclear és similar: l'exportina s'uneix a la càrrega dins el nucli en un procés facilitat per la RanGTP, surt pel porus nuclear i se separa de la càrrega al citoplasma.
Hi ha proteïnes especialitzades en l'exportació d'ARNm i ARNt madur al citoplasma una vegada s'ha completat la modificació posttranscripcional. Aquest mecanisme de control de la qualitat és important a causa del paper central d'aquestes molècules en la traducció proteica; l'expressió errònia d'una proteïna a causa d'una excisió incompleta dels exons o una incorporació incorrectad'aminoàcids tindria conseqüències negatives per la cèl·lula, de manera que l'ARN que arriba al citoplasma sense haver estat modificat completament és degradat, en lloc de ser utilitzat a la traducció.[5]
Assemblatge i desassemblatge
[modifica]
Al llarg de la seva vida, un nucli es pot trencar, o bé durant la divisió cel·lular o bé a conseqüència de l'apoptosi, una forma regulada de mort cel·lular. Durant aquests esdeveniments, els components estructurals del nucli (l'embolcall i la làmina) són degradats sistemàticament.
Durant el cicle cel·lular, la cèl·lula es divideix per formar dues cèl·lules noves. Per tal que aquest procés sigui possible, cal que cada cèl·lula filla tingui un conjunt complet de gens, un procés que requereix la replicació dels cromosomes i la segregació dels dos conjunts. Això es produeix quan els cromosomes replicats, les cromàtides germanes, s'uneixen a microtúbuls, que al seu torn estan units a diferents centrosomes. Aleshores, les cromàtides germanes són tibades a punts separats de la cèl·lula. En moltes cèl·lules, el centrosoma es troba al citoplasma, a l'exterior del nucli, car els microtúbuls serien incapaços d'unir-se a les cromàtides en presència de l'embolcall nuclear.[47] Per tant, a les primeres fases del cicle cel·lular, començant a la profase i finsaproximadament la prometafase, la membrana nuclear és desmantellada.[14] De manera similar, la làmina nuclear també és desassemblada durant el mateix període, un procés regulat per la fosforilació de les làmines.[48] Cap a finals del cicle cel·lular, es restaura la membrana nuclear i, aproximadament al mateix temps, les làmines nuclears són reassemblades per mitjà de la fosforilació de les làmines.[48]
Tanmateix, en els dinoflagel·lats l'embolcall nuclear roman intacte, els centrosomes es troben al citoplasma i els microtúbuls entren en contacte amb els cromosomes, les regions centromèriques dels quals són incorporades a l'embolcall nuclear (l'anomenada mitosi amb fus extranuclear). En molts altres protists (ex., ciliats o esporozous) i fongs, els centrosomes són intranuclears, i l'embolcall nuclear no es desacobla durant la divisió cel·lular.
L'apoptosi és un procés controlat en què els components estructurals de la cèl·lula són destruïts, provocant-ne la mort. Els canvis associats amb l'apoptosi afecten directament el nucli i els seus continguts, com per exemple en la condensació de la cromatina i la desintegració de l'embolcall i la làmina nuclears. La destrucció de les xarxes de làmines és controlada per proteases apoptòtiques especialitzades anomenades caspases, que fragmenten les proteïnes laminars i degraden així la integritat estructural del nucli. A vegades es fa servir la fragmentació de làmines com indicador al laboratori d'activitat de caspases en assajos que busquen activitat apoptòtica prematura.[14] Les cèl·lules que expressen làmines mutants resistents a les caspases són deficients en els canvis nuclears associats a l'apoptosi, cosa que suggereix que les lamines tenen un paper a l'hora d'engegar el procés que condueix a la degradació apoptòtica del nucli.[14] La inhibició de l'assemblatge de làmines indueix l'apoptosi.[49]
L'embolcall nuclear serveix de barrera per impedir que tant els virus ADN com els ADN penetrin al nucli. Alguns virus necessiten accedir a proteïnes de l'interior del nucli per tal de replicar-se i/o assemblar-se. Els virus ADN com ara l'herpesvirus es repliquen i assemblen al nucli cel·lular, i en surten per gemmació a través de la membrana nuclear interna. Aquest procés és acompanyat pel desassemblatge de les làmines de la cara nuclear de la membrana interior.[14]
Cèl·lules anucleades i polinucleades
[modifica]Tot i que la majoria de cèl·lules tenen un únic nucli, alguns tipus cel·lulars eucariotes no en tenen, i d'altres en tenen múltiples. Això pot ser un procés normal, com en la maduració dels glòbuls vermells dels mamífers, o el resultat d'una divisió cel·lular defectuosa.
Les cèl·lules anucleades manquen de nucli i per tant són incapaces de dividir-se per generar cèl·lules filles. La cèl·lula anucleada més coneguda és el glòbul vermell mamífer, o eritròcit, que també manca d'altres orgànuls com ara mitocondris, i serveix principalment de recipient de transport per dur oxigen dels pulmons als teixits del cos. Els eritròcits maduren per eritropoesi a la medul·la òssia, on perden el nucli, els orgànuls i els ribosomes. El nucli és expulsat durant el procés de diferenciació d'eritroblast a reticulòcit, que és el precursor immediat de l'eritròcit madur.[50] La presència de mutàgens pot induir l'alliberament d'alguns eritròcits immadurs "micronucleats" al flux sanguini.[51][52] Les cèl·lules anucleades també poden sorgir d'una divisió cel·lular defectuosa en què una cèl·lula filla manca de nucli i l'altra en té dos.
Les cèl·lules polinucleades contenen més d'un nucli. La majoria d'espècies de protozous acantaris[53] i alguns fongs en micorrizes[54] tenen cèl·lules naturalment polinucleades. Altres exemples són els paràsits intestinals del gènere Giardia, que tenen dos nuclis per cèl·lula.[55] En els humans, les cèl·lules dels músculs esquelètics, anomenades miòcits, esdevenen polinucleades durant el seu desenvolupament: la configuració resultant dels nuclis a prop de la perifèria de les cèl·lules permet maximitzar l'espai intracel·lular per les miofibril·les.[5] Les cèl·lules multinucleades també poden ser anormals en els humans; per exemple, les cèl·lules que sorgeixen de la fusió de monòcits i macròfags, conegudes com a cèl·lules gegants multinucleades, a vegades acompanyen la inflamació[56] i també estan implicades en la formació de tumors.[57]
Evolució
[modifica]Com que és la principal característica definidora de la cèl·lula eucariota, l'origen evolutiu del nucli ha estat objecte de gran especulació. S'han proposat quatre grans teories per explicar l'existència del nucli, però cap d'elles no ha aconseguit encara un ampli suport.[58]
La teoria coneguda com a "model sintròfic" proposa que una relació simbiòtica entre arqueus i bacteris creà la cèl·lula eucariota amb nucli. S'ha hipotetitzat que la simbiosi començà quan arqueus ancestrals, similars als metanògens actuals, envaïren i s'establiren dins d'eubacteris semblants als mixobacteris actuals, i acabaren formant el nucli primitiu. Aquesta teoria és anàloga a la teoria acceptada de l'origen dels mitocondris i cloroplasts dels eucariotes, que es creu que es desenvoluparen a partir d'una relació endosimbiòtica semblant entre protoeucariotes i bacteris aeròbics.[59] L'origen arqueuà del nucli és sostingut per les observacions que els arqueus i els eucariotes tenen gens similars per determinades proteïnes, incloent-hi les histones. Les observacions que els mixobacteris són mòtils, poden formar complexos pluricel·lulars i tenen cinases i proteïnes G semblants a les dels eucariotes també sostenen un origen bacterià de la cèl·lula eucariota.[60]
Un segon model suggereix que les cèl·lules protoeucariotes evolucionaren de bacteris sense cap fase endosimbiòtica. Aquest model es basa en l'existència de bacteris planctomicets moderns que tenen una estructura similar amb porus primitius i altres estructures membranals compartimentalitzades.[61] Una proposta similar postula que una cèl·lula semblant a un eucariota, el cronòcit, evolucionà en primer lloc i fagocità arqueus i bacteris per generar el nucli i la cèl·lula eucariota.[62]
El model més controvertit, conegut com a "eucariogènesi vírica", suggereix que el nucli envoltat per una membrana, així com altres trets eucariotes, s'originà amb la infecció d'un procariota per part d'un virus. Aquest suggeriment es basa en semblances entre els eucariotes i els virus, com ara cadenes lineals d'ADN, càping d'ARNm i una unió estreta amb les proteïnes (de manera anàloga a com ho fan les histones amb els embolcalls vírics). Una versió de la proposta suggereix que el nucli coevolucionà amb la fagocitosi per formar un "predador" cel·lular primitiu-[63] Una altra variant proposa que els eucariotes s'originaren a partir d'arqueus primitius infectats per poxvirus, basant-se en la semblança observada entre les ADN polimerases dels poxvirus i eucariotes actuals.[64][65] S'ha suggerit que la qüestió encara no resolta de l'evolució del sexe podria estar relacionada amb la hipòtesi de l'eucariogènesi vírica.[66]
Finalment, una proposta molt recent suggereix que les variants tradicionals de la teoria endosimbiòtica no són prou convincents per explicar l'origen del nucli eucariota. Aquest model, anomenat "hipòtesi de l'exomembrana", suggereix que el nucli s'originà a partir d'una única cèl·lula ancestral que desenvolupà una segona membrana cel·lular exterior; la membrana interior que envoltava la cèl·lula original esdevingué la membrana nuclear i desenvolupà estructures de porus cada vegada més complexes per permetre el pas de components cel·lulars sintetitzats a l'interior, com ara subunitats ribosòmiques.[67]
Referències
[modifica]- ↑ Leeuwenhoek, A. van: Opera Omnia, seu Arcana Naturae ope exactissimorum Microscopiorum detecta, experimentis variis comprobata, Epistolis ad varios illustres viros. J. Arnold et Delphis, A. Beman, Lugdinum Batavorum 1719–1730. Citat a: Dieter Gerlach, Geschichte der Mikroskopie. Verlag Harry Deutsch, Frankfurt del Main, Alemanya, 2009. ISBN 978-3-8171-1781-9
- ↑ Harris, H. The Birth of the Cell. New Haven: Yale University Press, 1999.
- ↑ Brown, Robert «On the Organs and Mode of Fecundation of Orchidex and Asclepiadea». Miscellaneous Botanical Works, i, 1866, pàg. 511–514.
- ↑ 4,0 4,1 Cremer, Thomas. Von der Zellenlehre zur Chromosomentheorie. Berlín, Heidelberg, Nova York, Tòquio: Springer Verlag, 1985. ISBN 3-540-13987-7. Versió en línia aquí
- ↑ 5,0 5,1 5,2 5,3 5,4 5,5 5,6 5,7 Lodish, H; Berk A, Matsudaira P, Kaiser CA, Krieger M, Scott MP, Zipursky SL, Darnell J.. Molecular Cell Biology. 5a edició. Nova York: WH Freeman, 2004.
- ↑ 6,0 6,1 6,2 6,3 Bruce Alberts, Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, Peter Walter. Molecular Biology of the Cell, Capítol 4, pàgines 191-234. 4a ed.. Garland Science, 2002.
- ↑ Clegg JS «Properties and metabolism of the aqueous cytoplasm and its boundaries». Am. J. Physiol., 246, 2 Pt 2, Febrer 1984, pàg. R133–51. PMID: 6364846.
- ↑ Paine P, Moore L, Horowitz S «Nuclear envelope permeability». Nature, 254, 5496, 1975, pàg. 109-114. DOI: 10.1038/254109a0. PMID: 1117994.
- ↑ «Capítol 3». A: Rodney Rhoades, Richard Pflanzer. Human Physiology. 3a ed.. Saunders College Publishing, 1996.
- ↑ Shulga N, Mosammaparast N, Wozniak R, Goldfarb D «Yeast nucleoporins involved in passive nuclear envelope permeability». J Cell Biol, 149, 5, 2000, pàg. 1027–1038. DOI: 10.1083/jcb.149.5.1027. PMID: 10831607.
- ↑ 11,0 11,1 Pemberton L, Paschal B «Mechanisms of receptor-mediated nuclear import and nuclear export». Traffic, 6, 3, 2005, pàg. 187–198. DOI: 10.1111/j.1600-0854.2005.00270.x. PMID: 15702987.
- ↑ Stuurman N, Heins S, Aebi U «Nuclear lamins: their structure, assembly, and interactions». J Struct Biol, 122, 1–2, 1998, pàg. 42–66. DOI: 10.1006/jsbi.1998.3987. PMID: 9724605.
- ↑ Goldman A, Moir R, Montag-Lowy M, Stewart M, Goldman R «Pathway of incorporation of microinjected lamin A into the nuclear envelope». J Cell Biol, 119, 4, 1992, pàg. 725–735. DOI: 10.1083/jcb.119.4.725. PMID: 1429833.
- ↑ 14,0 14,1 14,2 14,3 14,4 Goldman R, Gruenbaum Y, Moir R, Shumaker D, Spann T «Nuclear lamins: building blocks of nuclear architecture». Genes Dev, 16, 5, 2002, pàg. 533–547. DOI: 10.1101/gad.960502. PMID: 11877373.
- ↑ Moir RD, Yoona M, Khuona S, Goldman RD. «Nuclear Lamins A and B1: Different Pathways of Assembly during Nuclear Envelope Formation in Living Cells». Journal of Cell Biology, 151, 6, 2000, pàg. 1155–1168. DOI: 10.1083/jcb.151.6.1155. PMID: 11121432.
- ↑ Spann TP, Goldman AE, Wang C, Huang S, Goldman RD. «Alteration of nuclear lamin organization inhibits RNA polymerase II–dependent transcription». Journal of Cell Biology, 156, 4, 2002, pàg. 603–608. DOI: 10.1083/jcb.200112047. PMID: 11854306.
- ↑ Mounkes LC, Stewart CL «Aging and nuclear organization: lamins and progeria». Current Opinion in Cell Biology, 16, 2004, pàg. 322–327. DOI: 10.1016/j.ceb.2004.03.009. PMID: 15145358.
- ↑ Ehrenhofer-Murray A «Chromatin dynamics at DNA replication, transcription and repair». Eur J Biochem, 271, 12, 2004, pàg. 2335–2349. DOI: 10.1111/j.1432-1033.2004.04162.x. PMID: 15182349.
- ↑ Grigoryev S, Bulynko Y, Popova E «The end adjusts the means: heterochromatin remodelling during terminal cell differentiation». Chromosome Res, 14, 1, 2006, pàg. 53–69. DOI: 10.1007/s10577-005-1021-6. PMID: 16506096.
- ↑ Schardin, Margit; T. Cremer, H. D. Hager, M. Lang «Specific staining of human chromosomes in Chinese hamster x man hybrid cell lines demonstrates interphase chromosome territories». Human Genetics. Springer Berlin / Heidelberg, 71, 4, Desembre 1985, pàg. 281–287. Arxivat de l'original el 2019-09-13. DOI: 10.1007/BF00388452. PMID: 2416668 [Consulta: 17 octubre 2009].
- ↑ Lamond, Angus I.; William C. Earnshaw «Structure and Function in the Nucleus». Science, 280, 24-04-1998, pàg. 547–553. DOI: 10.1126/science.280.5363.547. PMID: 9554838.
- ↑ Kurz, A; S Lampel, JE Nickolenko, J Bradl, A Benner, RM Zirbel, T Cremer and P Lichter «Active and inactive genes localize preferentially in the periphery of chromosome territories». The Journal of Cell Biology. The Rockefeller University Press, 135, 1996, pàg. 1195–1205. Arxivat de l'original el 2007-09-29. DOI: 10.1083/jcb.135.5.1195. PMID: 8947544 [Consulta: 17 octubre 2009].
- ↑ NF Rothfield, BD Stollar «The Relation of Immunoglobulin Class, Pattern of Antinuclear Antibody, and Complement-Fixing Antibodies to DNA in Sera from Patients with Systemic Lupus Erythematosus». J Clin Invest, 46, 11, 1967, pàg. 1785–1794. PMID: 4168731.
- ↑ S Barned, AD Goodman, DH Mattson «Frequency of anti-nuclear antibodies in multiple sclerosis». Neurology, 45, 2, 1995, pàg. 384–385. PMID: 7854544.
- ↑ Hernandez-Verdun, Daniele «Nucleolus: from structure to dynamics». Histochem. Cell. Biol, 125, 125, 2006, pàg. 127–137. DOI: 10.1007/s00418-005-0046-4.
- ↑ 26,0 26,1 Lamond, Angus I.; Judith E. Sleeman «Nuclear substructure and dynamics». Current Biology, 13, 21, pàg. R825–828. DOI: 10.1016/j.cub.2003.10.012. PMID: 14588256.
- ↑ 27,0 27,1 27,2 Cioce M, Lamond A «Cajal bodies: a long history of discovery». Annu Rev Cell Dev Biol, 21, pàg. 105–131. DOI: 10.1146/annurev.cellbio.20.010403.103738. PMID: 16212489.
- ↑ 28,0 28,1 28,2 Pollard, Thomas D.; William C. Earnshaw. Cell Biology. Filadèlfia: Saunders, 2004. ISBN 0-7216-3360-9.
- ↑ 29,0 29,1 29,2 Dundr, Miroslav; Tom Misteli «Functional architecture in the cell nucleus». Biochem. J., 356, 2001, pàg. 297–310. DOI: 10.1146/annurev.cellbio.20.010403.103738. PMID: 11368755.
- ↑ Entrevista per correu electrònic de R. Sundby a Archa Fox sobre la mida dels paraspeckles. 7 de març del 2007.
- ↑ Goebel, H.H.; I Warlow «Nemaline myopathy with intranuclear rods—intranuclear rod myopathy». Neuromuscular Disorders, 7, 1, Gener 1997, pàg. 13–19. DOI: 10.1016/S0960-8966(96)00404-X. PMID: 9132135.
- ↑ 32,0 32,1 32,2 Matera AG, Frey MA. «Coiled Bodies and Gems: Janus or Gemini?». American Journal of Human Genetics, 63, 2, 1998, pàg. 317–321. DOI: 10.1086/301992. PMID: 9683623.
- ↑ Matera, A. Gregory «Of Coiled Bodies, Gems, and Salmon». Journal of Cellular Biochemistry, 70, 1998, pàg. 181–192. DOI: 10.1086/301992. PMID: 9671224.
- ↑ Saunders WS, Cooke CA, Earnshaw WC «Compartmentalization within the nucleus: discovery of a novel subnuclear region.». Journal of Cellular Biology, 115, 4, 1991, pàg. 919–931. DOI: 10.1083/jcb.115.4.919. PMID: 1955462
- ↑ Pombo A, Cuello P, Schul W, Yoon J, Roeder R, Cook P, Murphy S «Regional and temporal specialization in the nucleus: a transcriptionally active nuclear domain rich in PTF, Oct1 and PIKA antigens associates with specific chromosomes early in the cell cycle». EMBO J, 17, 6, 1998, pàg. 1768–1778. DOI: 10.1093/emboj/17.6.1768. PMID: 9501098.
- ↑ 36,0 36,1 Fox, Archa et al. «Paraspeckles:A Novel Nuclear Domain». Current Biology, 12, 2002, pàg. 13–25. Arxivat de l'original el 2012-12-08. DOI: 10.1016/S0960-9822(01)00632-7 [Consulta: 2 novembre 2009]. «no»
- ↑ 37,0 37,1 Fox, Archa; Wendy Bickmore. «Nuclear Compartments: Paraspeckles». Nuclear Protein Database, 2004. Arxivat de l'original el 2006-05-02. [Consulta: 6 març 2007].
- ↑ 38,0 38,1 Fox, A. et al. «P54nrb Forms a Heterodimer with PSP1 That Localizes to Paraspeckles in an RNA-dependent Manner». Molecular Biology of the Cell, 16, 2005, pàg. 5304–5315. DOI: 10.1091/mbc.E05-06-0587. PMID: 16148043. «no» PMID: 16148043
- ↑ Lamond AI, Spector DL «Nuclear speckles: a model for nuclear organelles». Nat. Rev. Mol. Cell Biol., 4, 8, Agost 2003, pàg. 605–12. DOI: 10.1038/nrm1172. PMID: 12923522.
- ↑ Handwerger, Korie E.; Joseph G. Gall «Subnuclear organelles: new insights into form and function». TRENDS in Cell Biology, 16, 1, Gener 2006, pàg. 19–26. DOI: 10.1016/j.tcb.2005.11.005. PMID: 16325406.
- ↑ Lehninger, Albert L.; David L. Nelson, Michael M. Cox.. Lehninger principles of biochemistry. 3a ed.. Nova York: Worth Publishers, 2000. ISBN 1-57259-931-6.
- ↑ Moreno F, Ahuatzi D, Riera A, Palomino CA, Herrero P. «Glucose sensing through the Hxk2-dependent signalling pathway.». Biochem Soc Trans, 33, 1, 2005, pàg. 265–268. DOI: 10.1042/BST0330265. PMID: 15667322. PMID: 15667322
- ↑ Görlich, Dirk; Ulrike Kutay «Transport between the cell nucleus and the cytoplasm». Ann. Rev. Cell Dev. Biol., 15, 1999, pàg. 607–660. DOI: 10.1042/BST0330265. PMID: 10611974.
- ↑ Nierhaus, Knud H.; Daniel N. Wilson. Protein Synthesis and Ribosome Structure: Translating the Genome. Wiley-VCH, 2004. ISBN 3527306382.
- ↑ Nicolini, Claudio A. Genome Structure and Function: From Chromosomes Characterization to Genes Technology. Springer, 1997. ISBN 0792345657.
- ↑ Watson, JD; Baker TA, Bell SP, Gann A, Levine M, Losick R.. «9–10». A: Molecular Biology of the Gene. 5a edició. Peason Benjamin Cummings; CSHL Press., 2004.
- ↑ Lippincott-Schwartz, Jennifer «Cell biology: Ripping up the nuclear envelope». Nature, 416, 6876, 07-03-2002, pàg. 31–32. DOI: 10.1038/416031a. PMID: 11882878.
- ↑ 48,0 48,1 Boulikas T «Phosphorylation of transcription factors and control of the cell cycle». Crit Rev Eukaryot Gene Expr, 5, 1, 1995, pàg. 1–77. PMID: 7549180.
- ↑ Steen R, Collas P «Mistargeting of B-type lamins at the end of mitosis: implications on cell survival and regulation of lamins A/C expression». J Cell Biol, 153, 3, 2001, pàg. 621–626. DOI: 10.1083/jcb.153.3.621. PMID: 11331311.
- ↑ Skutelsky, E.; Danon D «Comparative study of nuclear expulsion from the late erythroblast and cytokinesis». J Cell Biol, 60(3), Juny 1970, pàg. 625–635. DOI: 10.1083/jcb.153.3.621. PMID: 5422968.
- ↑ Torous, DK; Dertinger SD, Hall NE, Tometsko CR. «Enumeration of micronucleated reticulocytes in rat peripheral blood: a flow cytometric study». Mutat Res, 465(1–2), 2000, pàg. 91–99. DOI: 10.1083/jcb.153.3.621. PMID: 10708974.
- ↑ Hutter, KJ; Stohr M «Rapid detection of mutagen induced micronucleated erythrocytes by flow cytometry». Histochemistry, 75(3), 1982, pàg. 353–362. DOI: 10.1083/jcb.153.3.621. PMID: 7141888.
- ↑ Zettler, LA; Sogin ML, Caron DA «Phylogenetic relationships between the Acantharea and the Polycystinea: A molecular perspective on Haeckel's Radiolaria». Proc Natl Acad Sci USA, 94, 1997, pàg. 11411–11416. DOI: 10.1083/jcb.153.3.621. PMID: 9326623.
- ↑ Horton, TR «The number of nuclei in basidiospores of 63 species of ectomycorrhizal Homobasidiomycetes». Mycologia, 98(2), 2006, pàg. 233–238. DOI: 10.1083/jcb.153.3.621. PMID: 16894968.
- ↑ Adam RD «The biology of Giardia spp». Microbiol. Rev., 55, 4, Desembre 1991, pàg. 706–32. PMC: 372844. PMID: 1779932.
- ↑ McInnes, A; Rennick DM «Interleukin 4 induces cultured monocytes/macrophages to form giant multinucleated cells». J Exp Med, 167, 1988, pàg. 598–611. DOI: 10.1083/jcb.153.3.621. PMID: 3258008.
- ↑ Goldring, SR; Roelke MS, Petrison KK, Bhan AK «Human giant cell tumors of bone identification and characterization of cell types». J Clin Invest, 79(2), 1987, pàg. 483–491. DOI: 10.1083/jcb.153.3.621. PMID: 3027126.
- ↑ Pennisi E. «Evolutionary biology. The birth of the nucleus». Science, 305, 5685, 2004, pàg. 766–768. DOI: 10.1126/science.305.5685.766. PMID: 15297641.
- ↑ Margulis, Lynn. Symbiosis in Cell Evolution. San Francisco: W. H. Freeman and Company, 1981, p. 206–227. ISBN 0-7167-1256-3.
- ↑ Lopez-Garcia P, Moreira D. «Selective forces for the origin of the eukaryotic nucleus». Bioessays, 28, 5, 2006, pàg. 525–533. DOI: 10.1002/bies.20413. PMID: 16615090.
- ↑ Fuerst JA. «Intracellular compartmentation in planctomycetes». Annu Rev Microbiol., 59, 2005, pàg. 299–328. DOI: 10.1146/annurev.micro.59.030804.121258. PMID: 15910279.
- ↑ Hartman H, Fedorov A. «The origin of the eukaryotic cell: a genomic investigation». Proc Natl Acad Sci U S A., 99, 3, 2002, pàg. 1420–1425. DOI: 10.1073/pnas.032658599. PMID: 11805300.
- ↑ Bell PJ. (2001). "Viral eukaryogenesis: was the ancestor of the nucleus a complex DNA virus?" J Mol Biol Sep;53(3):251–256. PMID: 11523012
- ↑ Takemura M. (2001). Poxviruses and the origin of the eukaryotic nucleus. J Mol Evol 52(5):419–425. PMID: 11443345
- ↑ Villarreal L, DeFilippis V «A hypothesis for DNA viruses as the origin of eukaryotic replication proteins». J Virol, 74, 15, 2000, pàg. 7079–7084. DOI: 10.1128/JVI.74.15.7079-7084.2000. PMID: 10888648.
- ↑ Bell PJ. (2006). "Sex and the eukaryotic cell cycle is consistent with a viral ancestry for the eukaryotic nucleus." J Theor Biol 2006 November 7;243(1):54–63. PMID: 16846615
- ↑ de Roos AD «The origin of the eukaryotic cell based on conservation of existing interfaces». Artif Life, 12, 4, 2006, pàg. 513–523.. DOI: 10.1162/artl.2006.12.4.513. PMID: 16953783.
Bibliografia complementària
[modifica]- Goldman, Robert D.; Yosef Gruenbaum, Robert D. Moir, Dale K. Shumaker i Timothy P. Spann «Nuclear lamins: building blocks of nuclear architecture». Genes & Dev., 16, 16, 2002, pàg. 533–547. DOI: 10.1101/gad.960502. PMID: 11877373. Revisió sobre les làmines nuclears, explicant-ne l'estructura i les diverses funcions
- Görlich, Dirk; Ulrike Kutay «Transport between the cell nucleus and the cytoplasm». Ann. Rev. Cell Dev. Biol., 15, 15, 1999, pàg. 607–660. DOI: 10.1146/annurev.cellbio.15.1.607. PMID: 10611974.
- Article sobre el transport nuclear, que explica els principis del mecanisme i les diferents rutes de transport
- Lamond, Angus I.; William C. Earnshaw «Structure and Function in the Nucleus». Science, 280, 24-04-1998, pàg. 547–553. DOI: 10.1126/science.280.5363.547. PMID: 9554838.
- Article sobre el nucli, que explica l'estructura dels cromosomes dins de l'orgànul i descriu el nuclèol i altres cossos subnuclears
- Pennisi E. «Evolutionary biology. The birth of the nucleus». Science, 305, 5685, 2004, pàg. 766–768. DOI: 10.1126/science.305.5685.766. PMID: 15297641.
- Article sobre l'evolució del nucli, que explica una sèrie de teories diferents
- Pollard, Thomas D.; William C. Earnshaw. Cell Biology. Filadèlfia: Saunders, 2004. ISBN 0-7216-3360-9.: Llibre de text de nivell universitari centrat en la biologia cel·lular. Conté informació sobre l'estructura i la funció nuclears, incloent-hi el transport nuclear i els dominis subnuclears
Galeria d'imatges del nucli
[modifica]-
Comparació de cromosomes d'humans i ximpanzés.
-
Territoris cromosòmics de ratolins en diferents tipus cel·lulars.
-
24 territoris cromosòmics en cèl·lules humanes.



Enllaços externs
[modifica]- cellnucleus.com Web sobre l'estructura i funció del nucli del Department d'Oncologia de la Universitat d'Alberta. (anglès)
- The Nuclear Protein Database Informació sobre els components del nucli. (anglès)
- The Nucleus Collection Arxivat 2006-11-12 a Wayback Machine. a la biblioteca d'imatges i vídeos Arxivat 2011-06-10 a Wayback Machine. de The American Society for Cell Biology, amb fotografies i videoclips revisats científicament que il·lustren el nucli.
- Nuclear Envelope and Nuclear Import Section Arxivat 2008-02-16 a Wayback Machine. de Landmark Papers in Cell Biology Arxivat 2009-12-02 a Wayback Machine., Joseph G. Gall, J. Richard McIntosh, eds., conté comentaris digitalitzats i enllaços a articles investigatius seminals sobre el nucli. Publicat en línia a la biblioteca d'imatges i vídeos Arxivat 2011-06-10 a Wayback Machine. de The American Society for Cell Biology (anglès)
| Orgànuls i estructures cel·lulars | Acrosoma · Aparell de Golgi · Centríol · Cili · Citoplasma · Cloroplast · Citoesquelet · Endosoma · Flagel · Glioxisoma · Leucoplast · Lisosoma · Melanosoma · Membrana nuclear · Membrana plasmàtica · Mitocondri · Nuclèol · Nucli cel·lular · Peroxisoma · Reticle endoplasmàtic · Ribosoma · Vacúol · Vesícula · VTC's |
|---|---|
| Processos cel·lulars | |
| Metabolisme de les macromolècules | |