Oligocè
| Període | Època | Estatge | Edat (Ma) | |
|---|---|---|---|---|
| Neogen | Miocè | Aquitanià | (més recent) | |
| Paleogen | Oligocè | Catià | 27,82 | |
| Rupelià | 33,9 | |||
| Eocè | Priabonià | 37,2 | ||
| Bartonià | 40,4 | |||
| Lutecià | 48,6 | |||
| Ipresià | 55,8 | |||
| Paleocè | Thanetià | 58,7 | ||
| Selandià | 61,6 | |||
| Danià | 66,0 | |||
| Cretaci | Superior | Maastrichtià | (més antic) | |
| Comissió Internacional d'Estratigrafia (2023) | ||||
L'Oligocè[nota 1] o Oligocé[nota 2] és l'època geològica que començà fa 33,9 milions d'anys i s'acabà fa 23,03 milions d'anys. Es tracta de la tercera època del període Paleogen i de l'era del Cenozoic, que encara dura. El seu nom deriva de les paraules gregues antigues ὀλίγος (oligos), que significa ‘poc’, i καινός (kenós), que significa ‘recent’, i en el seu conjunt vol dir ‘poc recent’.[1]
La Grande Coupure, un canvi faunístic definit per l'arribada de nombroses espècies de mamífers d'origen asiàtic a Europa i l'extinció de moltes espècies autòctones d'aquest continent, marca el principi de l'Oligocè.[2] En canvi, el límit Oligocè-Miocè no queda marcat per un únic esdeveniment global de fàcil identificació, sinó que es troba en límits regionals entre el Catià, més càlid, i el Miocè inferior, més fred. Com en altres períodes geològics, els estrats que delimiten aquest període estan ben identificats, però no han estat datats amb màxima precisió.
El nom de l'Oligocè, definit per l'alemany Heinrich Ernst von Beyrich, es refereix a l'escassetat de nous grups moderns de mamífers en comparació amb l'explosió evolutiva de l'Eocè.
Subdivisions
[modifica]L'Oligocè se subdivideix en Rupelià i Catià, cadascun dels quals correspon a un dels dos estatges faunístics que componen aquesta època. Aquests estatges faunístics són:
- Rupelià: començà fa 33,9 milions d'anys amb el final de la Grande Coupure, un episodi d'extincions, de canvis faunístics i d'abundant especiació, sobretot entre els mamífers. Estratigràficament, el seu inici queda marcat per foraminífers planctònics i l'extinció del gènere Hantkenina. S'acabà fa 28,4 ± 0,1 milions d'anys i el seu nom, encunyat el 1850 pel geòleg belga André Hubert Dumont, fa referència al Rupel, un riu del nord de Bèlgica, afluent de l'Escalda.[3]
- Catià: començà fa 27,82 milions d'anys. Estratigràficament, el seu inici queda marcat per foraminífers planctònics i l'extinció del gènere Chiloguembelina. Durant aquest període tingué lloc l'erupció volcànica individual més gran coneguda: l'erupció de Fish Canyon a La Garita, amb un IEV de 9,2 (un volum d'ejecció de > 10.000 km³),[4] i que tingué lloc fa 27,51 milions d'anys.[5] L'estatge s'acabà fa 23,03 ± 0,05 milions d'anys i el seu nom, encunyat el 1894 pel geòleg alemany Theodor Fuchs, fa referència als cats, una tribu de l'antiga Germània, coneguts amb el nom llatí de Chatti.[6]
Paleoclima
[modifica]L'Oligocè ha estat descrit com un «període crucial, ja que fa entroncar el món arcaic de l'Eocè tropical amb els ecosistemes d'aspecte més modern del Miocè».[7] En efecte, aquesta època fou un període de transició que provocà grans alteracions en els ecosistemes, i molts grups d'animals foren incapaços d'adaptar-se i s'extingiren. El canvi més destacat fou l'obertura dels boscos que havien cobert la Terra fins a l'Eocè, creant grans planes obertes, matollars i fins i tot zones semidesèrtiques. Tanmateix, l'herba encara no havia experimentat la gran expansió que tindria més tard, de manera que les planes no s'assemblaven gaire a les sabanes d'avui en dia, i estaven poblades per altre tipus de vegetació.[7]

Amb la seva separació del continent australià fa uns 45 milions d'anys, durant l'Eocè, l'Antàrtida havia quedat privada del flux d'aigües equatorials que fins aleshores en suavitzaven el clima. Privada d'aquestes aigües càlides, l'Antàrtida es refredà i l'oceà Antàrtic inicià la seva glaciació, creant un flux d'aigua freda (vegeu Corrent circumpolar antàrtic) i banquises que reforçaren l'efecte refredant. Per això, durant l'Oligocè, el pol Sud quedà glaçat per primera vegada des de principis del Permià, mentre que el pol Nord romangué lliure de glaç.[8] Les regions de l'Antàrtida que encara no havien quedat cobertes de glaç tenien ecosistemes de tundra. A nivell mundial, l'Oligocè fou un període d'augment del volum de glaç,[9] que provocà una baixada de cinquanta-cinc metres en el nivell del mar, i que estigué estretament relacionat amb una caiguda de les temperatures.[10]
Malgrat un breu augment de les temperatures a l'acabament del Rupelià, durant l'Oligocè començà a accelerar-se i prendre força el refredament iniciat per un esdeveniment que tingué lloc fa uns cinquanta milions d'anys: l'esdeveniment d'Azolla. Creixeren grans quantitats de la falguera d'aigua dolça Azolla a l'oceà Àrtic. A mesura que s'enfonsaven al fons marí, les plantes quedaven incorporades als sediments, on no es descomponien a causa del baix nivell d'oxigen de les capes d'aigua profundes. La reducció resultant de la quantitat de carboni a l'atmosfera terrestre contribuí a transformar el planeta d'una «Terra hivernacle», prou càlida perquè prosperessin tortugues i palmeres als pols, a l'estat actual de «Terra glaçada».[11][12][13] Es creu que l'esdeveniment d'Azolla i les seves conseqüències són una de les principals causes, en combinació amb l'aïllament i glaciació de l'Antàrtida, del declivi gradual de la temperatura del planeta des de mitjan Paleogen fins avui en dia.
A part de l'aïllament de l'Antàrtida i els efectes de l'esdeveniment d'Azolla, que modificaren el clima a escala global, també hi hagué altres esdeveniments que influïren en el clima a menor escala. N'és un exemple la fase final de l'orogènesi laramiana, que havia començat al Paleocè i que promogué l'aparició d'un clima continental al que actualment és la meitat occidental dels Estats Units[7] Finalment, sembla que el clima de l'Oligocè permeté l'aparició, com a mínim en algunes ocasions, d'un pont de terra entre els continents eurasiàtic i nord-americà, unint les ecozones del Paleàrtic i el Neàrtic.
Paleogeografia
[modifica]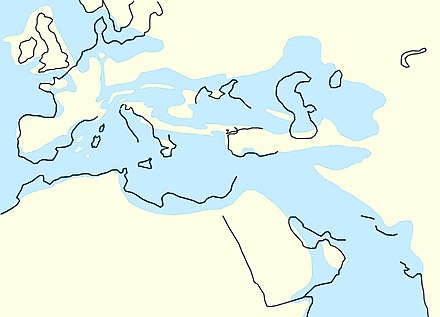
La tercera i última gran fase de la fragmentació del supercontinent Pangea tingué lloc a principis del Cenozoic, entre el Paleocè i l'Oligocè. Durant l'Oligocè, els continents no es trobaven gaire lluny de les seves posicions actuals, amb algunes excepcions. Sud-amèrica acabà de separar-se de l'Antàrtida i començà a moure's vers el nord; tanmateix, encara faltaven desenes de milions d'anys perquè es formés l'istme que més tard uniria el continent sud-americà amb el nord-americà.
Com que el casquet polar del Pol Nord encara no existia i el del Pol Sud encara no havia assolit la seva màxima extensió, el nivell del mar era més alt que el d'avui en dia. Per això, nombroses regions es trobaven submergides. A Europa, gran part del que actualment són Països Catalans es trobava sota l'aigua, així com Bèlgica, els Països Baixos, Dinamarca i Itàlia. Més a l'est, la zona caucàsica també es trobava sota el mar.
Orogènesi
[modifica]
El tancament gradual de l'oceà de Tetis, entre Euràsia al nord i Àfrica i l'Índia al sud, causà la formació de serralades muntanyoses com ara els Alps, els Pirineus i l'Himàlaia, en un gran procés orogènic anomenat orogènesi alpina. L'aixecament de les muntanyes formades per aquesta orogènesi separaren una bona part de l'oceà de Tetis de l'oceà Índic, creant un nou mar tancat que rep el nom de Paratetis. A l'acabament de l'oligocè, el subcontinent indi s'acoblà amb Àsia, accelerant la formació de l'Himàlaia. Aquest moviment convergent de les plaques encara continua avui en dia, i fa que l'Himàlaia sigui uns cinc mil·límetres més alt cada any.[15]
Durant l'Oligocè, l'escorça que havia quedat subduïda milions d'anys abans a la regió alpina començà a pujar de nou. Això provocà un ascens de la gruixuda escorça continental que al Miocè causaria una extensió. En el cas dels Alps, aquesta extensió només podia tenir lloc en una direcció oest-est, car la placa adriàtica encara convergia al sud. Es desenvolupà una zona d'impuls enorme que més tard esdevindria la línia periadriàtica. La zona també incloïa cisallament dextral resultant de l'extensió oest-est. Amb l'excepció del material austroalpí al·lòcton, aquest impuls evolucionà al límit de les plaques adriàtica i europea. Les regions centrals dels Alps s'alçaren i foren posteriorment erodides. Finestres i cúpules tectòniques com ara la finestra de Hohe Tauern es formaren d'aquesta manera.
Fauna
[modifica]Aus
[modifica]
L'Oligocè fou una època en què els ocells aquàtics i marins conegueren una gran radiació evolutiva. A l'hemisferi sud, els pingüins, que havien aparegut durant el Paleocè, continuaren diversificant-se i creixent a grans mides. N'és un exemple Archaeospheniscus, que presentava una barreja de característiques dels ocells primitius i dels pingüins moderns: el seu húmer encara és molt esvelt, a mig camí entre la forma típica de les ales d'ocells normals i la versió més gruixuda dels pingüins moderns. D'altra banda, el tarsometatars presenta una barreja peculiar de caràcters moderns i primitius. No se sap si això implica que aquest gènere és l'avantpassat de grups moderns o si representa un cas d'evolució paral·lela.[16][17]
Mentrestant, els anseriformes (els membres actuals dels quals inclouen els ànecs i les oques) continuaren el vertiginós procés evolutiu que havien començat a l'Eocè. A Austràlia, lliures de la competència dels mamífers euteris que patien les aus en altres indrets del món, els anseriformes creixeren fins a mides mai vistes abans.[18] Els dromornítids (una família d'anseriformes) més petits ja eren igual de grans que els casuaris d'avui en dia. Malgrat la seva aparença terrorífica, no és clar fins a quin punt eren carnívors. Els becs massius i potents d'algunes espècies suggereixen que almenys alguns membres de la família eren o bé caçadors i carronyaires o bé omnívors. Altres característiques, com ara les seves potes similars a peülles, l'estructura de l'estómac, i l'estructura ocular, que permetia un gran abast de visió però que creava un punt cec d'uns 40° (cosa que hauria representat un gran problema per la caça), suggereixen un estil de vida més herbívor i migratori. Tanmateix, els dromornítids de l'Oligocè encara eren relativament petits en comparació amb el que vindria després.[19]
Mentrestant, un dels llinatges més antics d'anseriformes, el dels presbiornítids, s'extingí al final de l'oligocè. L'estudi de la família Presbyornithidae és important per entendre l'evolució de les aus; les espècies que vivien en zones costaneres i que estaven menys especialitzades tendien a extingir-se menys que les espècies que estaven molt adaptades a un ecosistema en concret. Tanmateix, un cop les condicions del planeta tornaren a afavorir les formes especialitzades, els presbiornítids no pogueren suportar la seva competència i desaparegueren.
Mamífers
[modifica]Amb l'obertura dels espessos boscos que havien cobert el planeta durant l'Eocè, els mamífers pogueren augmentar dràsticament la seva mida.[7] Aquest augment en les dimensions dels animals es pot observar en gairebé tots els grups de mamífers pels quals es disposa d'un registre fòssil prou gran per fer comparacions.

Durant la primera part del Cenozoic, els perissodàctils (rinoceronts, èquids i altres) havien estat els ungulats dominants al planeta Terra. Tanmateix, els nous ecosistemes que oferia l'Oligocè donaren als artiodàctils (remugants, camells, porcs i altres) la possibilitat de destronar els perissodàctils, gràcies al seu particular aparell digestiu, més adaptat a la vegetació de l'Oligocè.[20] Tanmateix, algunes espècies de perissodàctils es convertiren en espècies de les més reeixides d'aquest període, incloent-hi el paracerateri, que amb una alçada de cinc metres i mig a l'espatlla fou el mamífer terrestre més gran de tots els temps;[21] o el calicoteri, un estrany animal brostejador d'Euràsia i Àfrica que només s'alimentava dels brots més joves.[7]
Després dels primers passos en l'evolució dels cetacis durant l'Eocè, una segona onada de mamífers s'adaptaren a la vida marina. Les proves moleculars recents suggereixen que els pinnípedes evolucionaren d'un avantpassat semblant a un os fa aproximadament 23 milions d'anys durant el Catià.[22] Els fòssils de pinnípede més primitius descoberts són els del gènere Enaliarctos, que visqué entre fa 24 i 22 milions d'anys. Es creu que era un bon nedador, però era capaç de moure's igual de bé a la terra que a l'aigua, més similar a una llúdria que als pinnípedes moderns. Fa temps que dura el debat de si les morses divergiren d'un avantpassat comú otàrid-fòcid, o si els fòcids divergiren d'un avantpassat comú otàrid-odobènid. Les proves més recents suggereixen que aquesta última hipòtesi és més probable.[23] A diferència de les foques actuals, els pinnípedes de l'Oligocè disposaven de dents carnisseres per tallar carn.[24]
El fòssil més antic conegut de diprotodont (l'ordre més gran de marsupials avui en dia) data del Catià d'Austràlia i pertany a Litokoala kanunkaensis.[25] Tanmateix, és probable que aquests animals siguin molt anteriors, tenint en compte la gran diversitat dels diprotodonts coneguts de principis del Miocè.[26] Els grans buits en el registre fòssil d'Austràlia i de Nova Guinea explicarien per què no s'han trobat fòssils anteriors de diprotodonts. Els dasiüromorfs, que inclouen gairebé tots els marsupials carnívors, també aparegueren a l'Oligocè.
A l'Oligocè tingueren lloc diversos episodis importants per l'evolució humana. Fa aproximadament trenta milions d'anys, el subordre dels haplorrins es dividí en dos subordres, els platirrins i els catarrins. Cinc milions d'anys més tard, els catarrins donaren origen a la superfamília dels hominoïdeus. Algunes innovacions del gènere Proconsul que es reflecteixen en Homo sapiens són la seva manca de cua,[27] l'estructura dels seus colzes, i la mida del cervell relativament gros en relació a la mida corporal.[28]
Peixos
[modifica]
La formació de Danata, al Turkmenistan, ha ofert fòssils de diversos tipus diferents de peixos. El grup més representatiu és el dels turkmènids, relacionat amb els lamprídids actuals. Aquesta família contenia tres gèneres: Danatinia (D. casca) i Turkmene (T. finitimus). Analectis (A. pala) també és un peix de l'Oligocè del Turkmenistan, però prové d'un jaciment diferent al de les altres dues espècies. Els tres gèneres eren relativament petits en comparació amb els lamprídids d'avui en dia, però a l'altre extrem es trobava Megalampris keyesi, un lamprídid de l'Oligocè superior de Nova Zelanda que visqué fa aproximadament vint-i-sis milions d'anys i que mesurava uns quatre metres de llargada (és a dir, el doble que l'espècie de lamprídid més gran vivent, Lampris guttatus).[29] Amb la desaparició d'Analectis s'extingiren els turkmènids.
Rèptils
[modifica]L'Oligocè fou el testimoni de l'aparició de dos dels gèneres de cocodrils encara vivents avui en dia, Osteolaemus i Crocodylus; possiblement, aquest últim gènere deriva del primer, més primitiu.[30] Les restes d'Osteolaemus descobertes al Zaire són considerablement més grans que les de l'espècie vivent, el cocodril nan, que és l'espècie de cocodril més petita del món.[31]

Meiolania platyceps.
També es troben cocodrils a Oceania. Mekosuchus whitehunterensis visqué durant l'Oligocè superior a Queensland. Es caracteritza per la seva petitesa i per formar part de l'últim grup de cocodrils terrestres, que s'extingí a l'Holocè, probablement a causa de l'arribada dels humans a la regió del món on habitava. Un altre cocodril relacionat amb Mekosuchus era Australosuchus clarkae, que visqué al sud d'Austràlia entre l'Oligocè superior i el Miocè inferior. Els gaviàlids també prosperaren durant aquest període: Gavialosuchus aparegué a l'est de Nord-amèrica durant l'Oligocè superior i s'estengué a Europa durant el Miocè inferior. Era un cocodril d'estuari que vivia en aigües marines somes. A diferència dels mekosuquins, tenia un musell llarg i era molt gran: G. carolinensis mesurava com a mínim 5,37 metres de llarg.[32]
D'altra banda, també es coneixen restes fòssils destacables de tortugues. A més d'Anosteira, Chelydropsis i Psephophorus, és d'especial interès Meiolania, que amb una llargada de dos metres i mig fou la tortuga terrestre més gran de tots els temps.[33] Tanmateix, la seva mida no era l'únic tret característic d'aquesta tortuga. Meiolania tenia un crani inusual amb moltes protrustions semblants a bonys o banyes. Tenia dues grans banyes que apuntaven als costats, donant al crani una amplada total de 60 cm. Aquestes banyes li haurien impedit amagar el cap dins la closca. La cua estava protegida per «anells» blindats i presentava punxes a l'extrem. D'una certa manera, dona la impressió que la morfologia de Meiolania convergia amb la dels dinosaures ancilosàurids i els xenartres gliptodonts.[34]
Mol·luscs
[modifica]Els nàutils són animals cefalòpodes considerats fòssils vivents, a causa del fet que la seva aparença gairebé no ha canviat en els últims 500 milions d'anys. Nautilus praepompilius és l'espècie fòssil de nàutil que s'extingí més recentment; se n'han trobat fòssils a la formació de Txegan al Kazakhstan. El seu parent vivent més proper és N. pompilius.
Els gastròpodes també tenen un abundant registre fòssil a l'Oligocè; entre els gastròpodes d'aquesta època es troben Clavilithes (Europa, Nord-amèrica i Sud-amèrica), Ficopsis (tots els continents menys l'Antàrtida) o Rimella (Europa).
Flora
[modifica]
Els primers fòssils que es poden definir amb tota certesa com a herbes daten del Paleocè superior d'Amèrica del Nord, fa un xic més de cinquanta-cinc milions d'anys.[7] Tanmateix, les herbes foren amb gran probabilitat rares durant la primera part del Cenozoic, quan el calorós i humit clima regnant afavoria un altre tipus de plantes. Molts científics creuen que el clima més fresc i sec de l'Oligocè donà a les herbes una primera possibilitat d'expandir-se; a mesura que els boscos tropicals retrocedien vers l'equador, les herbes haurien colonitzat els boscos temperats que omplien el seu lloc.[7]
Hi ha indicis que els herbassars (ecosistemes on les herbes són la vegetació dominant) aparegueren abans a Àsia que la resta del món, segons es pot deduir de les adaptacions observades en els rosegadors i lagomorfs fòssils d'aquest continent. Aquests animals tenien unes dents i un aparell digestiu especialment adaptats per processar les bastes fulles i tiges de les herbes.[7]
Durant l'Oligocè, les plantes suculentes i els cactus desenvoluparen una nova ruta de fixació del carboni que augmenta l'eficiència de la fotosíntesi; es tracta del metabolisme àcid de les crassulàcies.[35] Tanmateix, aquestes plantes no esdevindrien ecològicament significatives fins al final del Miocè.[36] D'altra banda cal remarcar que alguns fòssils carbonitzats conserven teixits organitzats en l'anatomia de Kranz, amb cèl·lules fasciculars intactes,[37] permetent identificar sense cap mena de dubte la presència del metabolisme vegetal C₄ en gramínies d'aquell temps.
Submersió de l'Oligocè
[modifica]
Nova Zelanda cavalca el límit entre dues plaques tectòniques. La subducció de la placa pacífica sota la placa australiana resulta en vulcanisme, especialment a la zona volcànica de Taupo (Illa del Nord). L'energia geotèrmica produïda és utilitzada en nombroses plantes d'energia hidrotèrmiques,[38] Alguns punts volcànics també són cèlebres destinacions turístiques, com ara els guèisers de Rotorua.
La particular situació geològica de Nova Zelanda es troba a l'origen de l'anomenada submersió de l'Oligocè. A principis de l'Oligocè, la massa terrestre de Nova Zelanda començà a enfonsar-se davant l'impuls de la placa australiana vers el Pacífic, fins que fa 30-25 milions d'anys, només una cinquena part de la Nova Zelanda actual es trobava per sobre el nivell del mar.[39] Aquesta submersió tingué un doble efecte. D'una banda, és probable que moltes espècies s'extingissin completament, causant una gran pèrdua en la biodiversitat. De l'altra banda, les parts més elevades de la Nova Zelanda actual quedaren aïllades en forma d'una cadena d'illes que romangueren separades durant milions d'anys. Això permeté que les poblacions que havien quedat aïllades a cada illa seguissin camins evolutius diferents i provocant una gran especiació: el resultat foren onze o més moes, quatre o més kiwis, nombrosos lacèrtids, molts invertebrats i múltiples plantes.[39]
Aquesta submersió arribà a la seva fi quan la massa terrestre de Nova Zelanda quedà encallada a la zona de subducció entre la placa australiana i la pacífica. Com que era massa gran com per ser subduïda, quedà a cavall entre les dues plaques, cosa que ha provocat una intensa activitat volcànica i orogènica al llarg dels últims milions d'anys.
Jaciments
[modifica]Aquests són alguns jaciments destacats de l'Eocè.
Hsanda Gol
[modifica]
La formació de Hsanda Gol és un jaciment paleontològic situat a la part mongol del desert del Gobi, especialment conegut per la seva riquesa en fòssils de mamífers de l'època de l'Oligocè, com ara hienodonts, entelodonts o els fòssils més complets coneguts de paracerateris. Entre aquests últims es troba un esquelet únic de paracerateri que havia mort dempeus. Probablement degué morir atrapat en sorra movent, i la posició de les potes indica que estava lluitant per escapar-se'n.
L'exploració paleontològica del segle xix i principis del segle xx s'havia concentrar principalment en jaciments europeus i nord-americans, com les fosforites del Carcí o la formació de Hell Creek. Tanmateix, Roy Chapman Andrews, del Museu Americà d'Història Natural, aconseguí reunir fons per fer una expedició al desert del Gobi, fins aleshores ignorat pels perills que representava. Andrews esperava trobar al Gobi l'anella perduda entre els simis i els humans. El que descobrí fou uns estrats que permeteren ampliar molt la informació coneguda sobre la fauna asiàtica de l'Oligocè. Tanmateix, la recerca científica a Hsanda Gol es veié interrompuda el 1930 a causa de la inquietud política de la zona, i el jaciment no tornà a quedar obert als científics estrangers fins als anys noranta.
Riversleigh
[modifica]
Riversleigh, situat al nord-oest de Queensland, és un dels jaciments paleontològics més cèlebres d'Austràlia. Aquesta zona de 100 km² té restes fòssils de mamífers, ocells i rèptils de l'Oligocè i el Miocè. El jaciment fou declarat Patrimoni de la Humanitat el 1994 i és una extensió del Parc Nacional de Lawn Hill.
Els fòssils de Riversleigh es troben en roques calcàries, al costat de basses d'aigua dolça rica en calç i en coves, i data de quan l'ecosistema evolucionava d'una rica jungla a una comunitat d'herbassar semiàrida. S'han identificat trenta-cinc fòssils de ratpenats, fent que sigui el jaciment més ric del món en aquest sentit. Entre les troballes de l'Oligocè de Riversleigh són especialment destacables Nimbacinus, un parent prehistòric del llop marsupial; i Baru, un cocodril mekosuquí amb el cap en forma de mitja lluna.
Formació del John Day
[modifica]La formació del John Day (Oregon, Estats Units d'Amèrica) representa una sèrie d'ecosistemes de boscos caducifolis datant des de l'Eocè superior (37 Ma) fins al Miocè inferior (20 Ma). S'hi han descobert més de cent grups de mamífers, incloent-hi representants de gossos, gats, porcs, cavalls, camells, rinoceronts, rosegadors, i gèneres com ara Temnocyon[40] o Merycoidodon. Durant aquest temps, múltiples esdeveniments volcànics deixaren la seva «empremta» en capes primes de cendra volcànica que s'enduriren en tova. Aquestes capes han estat datades amb precisió per mitjans radiomètrics i per comparació amb les mateixes capes en altres localitats, cosa que permet utilitzar-les com a marcadores temporals en aquesta formació.
Notes
[modifica]- ↑ En els parlars orientals.
- ↑ En els parlars occidentals.
Referències
[modifica]- ↑ Riba i Arderiu i Reguant i Serra, 1986, «Oligocè».
- ↑ Gosàlbez, 1987, «Els mamífers fòssils» (J. Agustí).
- ↑ Rohde, Robert A. «Rupelian ICS Stage». GeoWhen Database. Comissió Internacional d'Estratigrafia, 2005. Arxivat de l'original el 22 nov 2008. [Consulta: 13 setembre 2008].
- ↑ Mason, Ben G.; Pyle, David M., Oppenheimer, Clive «The size and frequency of the largest explosive eruptions on Earth». Bulletin of Volcanology, 66, 8, 2004, pàg. 735–748. DOI: 10.1007/s00445-004-0355-9.
- ↑ Lanphere, Marvin A.; Baadsgaard, H. «Precise K–Ar, 40Ar/39Ar, Rb–Sr and U/Pb mineral ages from the 27.5 Ma Fish Canyon Tuff reference standard». Chemical Geology, 175, 3–4, 2001, pàg. 653–671. DOI: 10.1016/S0009-2541(00)00291-6.
- ↑ Rohde, Robert A. «Chattian ICS Stage». GeoWhen Database. Comissió Internacional d'Estratigrafia, 2005. Arxivat de l'original el 18 nov 2008. [Consulta: 13 setembre 2008].
- ↑ 7,0 7,1 7,2 7,3 7,4 7,5 7,6 7,7 Haines, Tim. «Land of the Giants». A: Walking with Beasts. Londres: BBC Books, 2001 [Consulta: 13 setembre 2008].
- ↑ Christopher Scotese. «Oligocene Climate». Paleomap Project, 2000. [Consulta: 13 setembre 2008].
- ↑ Lorraine E. Lisiecki Nov 2004; A Pliocene-Pleistocene stack of 57 globally distributed benthic δ18O records Brown University, PALEOCEANOGRAPHY, VOL. 20
- ↑ Kenneth G. Miller Jan-Feb 2006; Eocene–Oligocene global climate and sea-level changes St. Stephens Quarry, Alabama GSA Bulletin, Rutgers University, NJ «Enllaç».
- ↑ Tim Appenzeller «Great green north». National Geographic, 5-2005. Arxivat de l'original el 2007-06-01 [Consulta: 13 setembre 2008].
- ↑ Doug MacDougall. Frozen Earth: The Once and Future Story of Ice Ages. University of California Press, 2004. ISBN 0-520-24824-4.
- ↑ Kieran Mulvaney. At the Ends of the Earth: A History of the Polar Regions. Washington, D.C.: Island Press, 2001. ISBN 1-55963-908-3.
- ↑ «Geologic Provinces of the United States: Rocky Mountains» (HTML). USGS Geology in the Parks. Arxivat de l'original el 2004-12-04. [Consulta: 10 desembre 2006]. (public domain source)
- ↑ Husson, Laurent; Jean-Louis Mugnier, Pascal Leturmy i Gérard Vidal «Kinetics and Sedimentary Balance of the Subhimalayan Zone, West Nepal». Thrust Tectonics and Hydrocarbon Systems, 82, 2004, pàg. 115-130. Arxivat de l'original el 2007-04-18 [Consulta: 20 setembre 2008].
- ↑ Marples, Brian J. (1952): Early Tertiary penguins of New Zealand. New Zealand Geol. Surv., Paleont. Bull. 20: 1-66.
- ↑ Simpson, George Gaylord (1971): A review of the pre-Pleistocene penguins of New Zealand. Bulletin of the American Museum of Natural History 144: 319–378. Document PDF
- ↑ Murray, P. F. & Megirian, D. (1998): The skull of dromornithid birds: anatomical evidence for their relationship to Anseriformes (Dromornithidae, Anseriformes). Records of the South Australian Museum 31: 51-97.
- ↑ Rich, P. (1979): The Dromornithidae, an extinct family of large ground birds endemic to Australia. Bureau of National Resources, Geology and Geophysics Bulletin 184: 1-196.
- ↑ Janis, Christine & Jarman, Peter. Macdonald, D. The Encyclopedia of Mammals. Nova York: Facts on File, 1984, p. 498-499. ISBN 0-87196-871-1.
- ↑ Microsoft Encarta. «Mammal». Entrada de l'Encarta. Microsoft Corpotation, 2007. Arxivat de l'original el 2008-04-15. [Consulta: 30 abril 2008].
- ↑ John J. Flynn et al. «Molecular Phylogeny of the Carnivora» (Scholar search). Systematic Biology, 54, 2005, pàg. 317 – 337. Arxivat de l'original el 2007-06-30 [Consulta: 12 novembre 2008].
- ↑ Ulfur Arnason, Anette Gullberg, Axel Janke, et al. «Pinniped phylogeny and a new hypothesis for their origin and dispersal». Molecular Phylogenetics and Evolution, 41, 2006, pàg. 345–354. DOI: 10.1016/j.ympev.2006.05.022.1
- ↑ Berta, A., C.E. Ray and A.R. Wyss. 1989. Skeleton of the oldest known pinniped, Enaliarctos mealsi. Science 244:60-62.
- ↑ Entrada «Diprotodontia» de la Paleobiology Database (en anglès). [Consulta: 20 desembre 2022].
- ↑ Robert William Mederith, Phylogeny and evolution of Diprotodontia marsupials (2008). Thesis (Ph. D.)--University of California, Riverside
- ↑ Fleagle, J.G. 1988. Primate Adaptation and Evolution. Academic Press: Nova York.
- ↑ Conroy, G.C. 1990. Primate Evolution. W.W. Norton and Co.: Nova York.
- ↑ Michael D. Gottfried; R. Ewan Fordyce & Seabourne Rust «Megalampris keyesi, a giant moonfish (Teleostei, Lampridiformes) from the late Oligocene of New Zealand». Journal of Vertebrate Paleontology, 26, 3, 2006, pàg. 544-551. DOI: 10.1671/0272-4634(2006)26[544:MKAGMT]2.0.CO;2.
- ↑ Aoki, R. (1976). On the generic status of Mecistops (Crocodylidae) and the origin of Tomistoma and Gavialis. Bulletin of Atagawa Institute, 6,7; 23-30
- ↑ Aoki, R. (1992). Fossil crocodilians from the late Tertiary strata in the Sinda Basin, Eastern Zaire. African Study Monographs, Suppl. 17; 67-85
- ↑ Erickson, Bruce R.; & Sawyer, Glen T.. The estuarine crocodile Gavialosuchus carolinensis n. sp. (Crocodylia: Eusuchia) from the late Oligocene of South Carolina, North America. St. Paul: The Science Museum of Minnesota, 1996, p. 1-47 (The Science Museum of Minnesota St. Paul, Minnesota Monograph 3, Paleontology).
- ↑ Barry Cox, Colin Harrison, R.J.G. Savage, and Brian Gardiner. (1999): The Simon & Schuster Encyclopedia of Dinosaurs and Prehistoric Creatures: A Visual Who's Who of Prehistoric Life. Simon & Schuster
- ↑ Vegeu aquesta imatge per una il·lustració de com hauria semblat Meiolania en vida.
- ↑ Osborne, C.P.; Beerling, D.J. «Review. Nature's green revolution: the remarkable evolutionary rise of C₄ plants». Philosophical Transactions: Biological Sciences, 361, 1465, 2006, pàg. 173-194. DOI: 10.1098/rstb.2005.1737 [Consulta: 11 febrer 2008].[Enllaç no actiu]
- ↑ Retallack, G.J. «Neogene Expansion of the North American Prairie». PALAIOS, 12, 4, 1997, pàg. 380–390. DOI: 10.2307/3515337 [Consulta: 11 febrer 2008].
- ↑ Thomasson, J.R.; Nelson, M.E., Zakrzewski, R.J. «A Fossil Grass (Gramineae: Chloridoideae) from the Miocene with Kranz Anatomy». Science, 233, 4766, 1986, pàg. 876–878. DOI: 10.1126/science.233.4766.876.
- ↑ Matthew Hall (2004) Existing and Potential Geothermal Resource for Electricity Generation. Ministry for Economic Development.
- ↑ 39,0 39,1 «The Nature of New Zealand's Land Environment». Ministeri del Medi Ambient de Nova Zelanda. Arxivat de l'original el 2008-10-16. [Consulta: 13 novembre 2008].
- ↑ Hunt, Robert M, Jr. (2004) «Global Climate and the Evolution of Large Mammalian Carnivores during the Later Cenozoic in North America». A: Cenozoic Carnivores and Global Climate de Robert M. Hunt, Jr.
Bibliografia
[modifica]- Gosàlbez, J. Història natural dels Països Catalans. Volum 13: Amfibis, rèptils i mamífers. Enciclopèdia Catalana, 1987. ISBN 84-7739-005-3.
- Riba i Arderiu, O.; Reguant i Serra, S. Una taula dels temps geològics. Institut d'Estudis Catalans, 1986. ISBN 9788472830776.