Usmjerenost (genetika)

Usmjerenost, u molekulskoj biologiji i biohemiji je hemijska orijentacija od kraja do kraja jednog lanca nukleinskih kiselina. U jednom lancu DNK ili RNK, hemijska konvencija imenovanja atoma ugljika u nukleotidnom šećernom prstenu znači da će postojati 5′ -kraj (obično se izgovara "pet prim kraj"), koji često sadrži fosfatnu grupu vezanu za 5' ugljik prstena riboze i 3'-end (obično se izgovara "tri prim kraj"), koji je tipski nemodificiran od supstituenta riboznog –OH. U dvostrukoj spirali DNK, niti se pružaju u suprotnim smjerovima, kako bi se omogućilo uparivanje baza između njih, što je neophodno za replikaciju ili transkripciju kodiranih informacija.[1]
Nukleinske kiseline mogu se sintetizirati in vivo, u smjeru 5′-do-3′, jer se polimeraze koje okupljaju različite vrste novih lanaca uglavnom oslanjaju na energiju koja nastaje razlaganjem nukleozid-trifosfatnih veze za vezanje novih nukleozidnih monofosfata na 3′-hidroksil (–OH) grupu, putem fosfodiesterske veze. Relativni položaj struktura duž lanca nukleinske kiseline, uključujući I gen i razne proteine veznog mjesta, obično se opisuje ili kao uzvodno (prema 5'-kraju) ili nizvodno (prema kraju 3′). (Vidi takođe uzvodno i nizvodno.)
Usmjerenost je povezana, ali neovisna o smislenosti. Transkripcija jednolančane RNK iz dvolančane matrica DNK zahtijeva odabir jednog predloška (polu)lanca DNK kao lanca matrice koji izravno djeluje s novonastalom RNK zbog komplementarnosti sekvenci. Drugi lanac se ne kopira direktno, ali nužno će njegova sekvenca biti slična onoj u RNAK. Mjesta pokretanja transkripcije obično se javljaju na oba lanca DNK organizma i određuju mjesto, smjer i okolnosti pod kojima će se transkripcija dogoditi. Ako transkript kodira jedan ili (rijetko) više protein, prevođenje svakog proteina u ribosomima nastavit će se u smjeru od 5′ do 3′ i protezat će se od njegovog N-kraja do C-kraja. Naprimjer, u tipiskm genu, početni kodon (5′-ATG-3′) je sekvenca DNK unutar smislenog lanca. Transkripcija započinje na uzvodnom mjestu (u odnosu na smisleni lanac) i dok prolazi kroz regiju kopira 3′-TAC-5′ iz lanca predloška da bi proizvela 5′-AUG-3′ unutar informacijske RNK (iRNK). iRNK se skenira u riboomima s 5′ kraja, gdje startni kodon usmjerava ugradnju metionina (umjesto toga, bakterije, mitohondrije i plastidi koriste N-formilmetionin) na N-kraju proteina. Prema dogovoru, pojedinačni lanci sekvenci DNK i RNK zapisuju se u pravcu od 5′ do 3′, osim ako je potrebno za ilustraciju obrasca uparivanja baza.
5′ – kraj
[uredi | uredi izvor]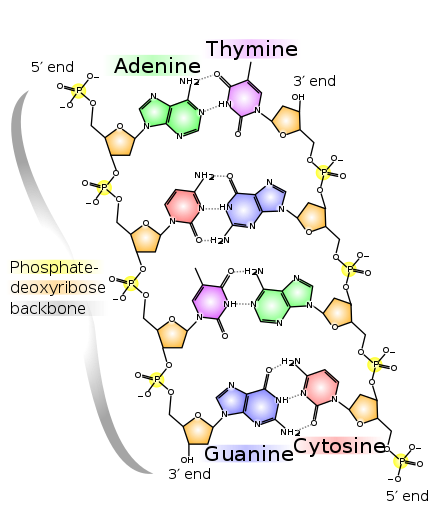
5′-kraj (izgovara se kao „pet prim kraj“) označava kraj lanca DNK ili RNK, koji ima peti ugljik u šećernom prstenu dezoksiribote ili riboze na svom kraju. Fosfatna grupa vezana za 5′-kraj dozvoljava ligaciju dva nukleotida, tj. kovalentno vezanje 5′-fosfata za 3′-hidroksilnu grupu drugog nukleotida, da bi se formirala fosfodiesterska veza. Uklanjanje 5′-fosfata sprečava podvezivanje. Da bi se sprečilo neželjeno vezivanje nukleinske kiseline (npr. samoligacija plazmidnog vektora u kloniranju molekula DNK), molekulski biolozi obično uklanjaju 5′-fosfat sa fosfatazazama.
5'-kraj novonastajuće iRNK je mjesto na kojem se post-transkripcijsko kapiranje, proces koji je od vitalnog značaja za stvaranje zrele informacijske RNK. Ograničenje povećava njenu stabilnost, dok se podvrgava translaciji, pružajući otpor degradacijskim efektima egzonukleaza. Sastoji se od metiliranog nukleotida (metilgvanozin) vezanog za transportnu RNK u rijetkoj 5′-do 5′-trifosfatnoj vezi.
5'bočni regioni gena često označavaju region DNK koji se ne transkribuje u RNK. 5-bočno područje sadrži promotorski gen, a može sadržavati i pojačivače ili druga mesta vezanja za proteine.
5′- neprevedenih regija (5′-UTR) je područje gena koji se transkribira u iRNK i nalazi se na njenom 5′-kraju. Ovo područje iRNK može ali ne mora biti prevedeno, ali je obično uključeno u regulaciju prevođenja. 5'-neprevedeno područje je dio DNK koji započinje od mjesta kape i proteže se do baze neposredno prije kodona iniciranja translacije AUG kodona glavne kodirajuće sekvence. Ova regija može imati sekvence, kao što su mjesto vezanja ribosoma i Kozakova sekvenca, koje određuju efikasnost translacije iRNK ili koje mogu uticati na njenu stabilnost.
3′–kraj
[uredi | uredi izvor]
3′-kraj (tri prim kraj) lanca je nazvan tako što završava na hidroksilnoj grupi trećeg ugljika u prstenu šećera, a poznat je kao repni kraj. 3′-hidroksil je neophodan u sintezi novih molekula nukleinske kiseline jer je podvezana (spojena) sa 5′-fosfatom odvojenog nukleotida, omogućavajući stvaranje lanaca povezanih nukleotida.
U molekulskoj biologiji mogu se koristiti nukleotidi kojima nedostaje 3′-hidroksil (didezoksiribonukleotidi) da prekine replikaciju DNK. Ova tehnika je poznata kao metod prekida lanca didezoksi ili Sangerov metod i koristi se za određivanje redoslijeda nukleotida u DNK.
3′-kraj novonastajuće iRNK je mjesto post-transkripcijske poliadenilacije, koje veže lanac od 50 do 250 adenozinskih ostataka za proizvodnju zrele informacijske RNK. Ovaj lanac pomaže u određivanju koliko dugo iRNK traje u ćeliji, utičući na to koliko se proteina iz nje proizvodi.
Regija 3′-bočna je regija DNK koja se ne kopira u zrelu iRNK, ali je prisutna uz 3'-kraj gena. Prvobitno se smatralo da 3'-bočna DNK uopće nije transkribirana, ali je otkriveno da je transkribirana u RNK i brzo uklonjena, tokom prerade primarnog transkripta, da bi stvorila zrela iRNK. 3'-bočno područje često sadrži sekvence koje utiču na formiranje poruke 3'-kraja. Može sadržavati pojačivače ili druga mjesta na koja se proteini mogu vezati.
3′-neprevedeno područje (3′-UTR) je područje DNK koje se transkribuira u iRNK i postaje 3'-kraj poruke , ali koji ne sadrži sekvencu za kodiranje proteina. Sve između stop kodona i poli-A repa smatra se 3′-neprevedenim. 3′-neprevedeno područje može uticati na efikasnost translacije iRNK ili na njenu stabilnost. Također ima sekvence potrebne za dodavanje poli (A) repa poruci, uključujući heksanukleotid AAUAAA.
Također pogledajte
[uredi | uredi izvor]Reference
[uredi | uredi izvor]- ^ Harvey Lodish; Arnold Berk; Paul Matsudaira; Chris A. Kaiser (2004). Molecular Cell Biology (5th izd.). New York City: W.H. Freeman and Company. ISBN 978-0-7167-4366-8.