TMEM268
| TMEM268 | |||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Identifikatori | |||||||||||||||||||||||||
| Aliasi | TMEM268 | ||||||||||||||||||||||||
| Vanjski ID-jevi | MGI: 1913920 HomoloGene: 17005 GeneCards: TMEM268 | ||||||||||||||||||||||||
| |||||||||||||||||||||||||
| |||||||||||||||||||||||||
| |||||||||||||||||||||||||
| Ortolozi | |||||||||||||||||||||||||
| Vrste | Čovjek | Miš | |||||||||||||||||||||||
| Entrez | |||||||||||||||||||||||||
| Ensembl | |||||||||||||||||||||||||
| UniProt | |||||||||||||||||||||||||
| RefSeq (mRNK) | |||||||||||||||||||||||||
| RefSeq (bjelančevina) | |||||||||||||||||||||||||
| Lokacija (UCSC) | Chr 9: 114.61 – 114.65 Mb | Chr 4: 63.48 – 63.5 Mb | |||||||||||||||||||||||
| PubMed pretraga | [3] | [4] | |||||||||||||||||||||||
| Wikipodaci | |||||||||||||||||||||||||
| |||||||||||||||||||||||||





Transmembranski protein 268 je protein koji je kod ljudi kodiran genom TMEM268. Ovaj transmembranski protein sadrži 342 aminokiselina, sa osam alternativno prerađenih varijanti. Identificiran je u organizmima od obične vinske mušice do primata. Do danas, njegova ekspresija nije nađena kod jednostavnijih organizama od insekata.
| C9orf91 | |||||||
|---|---|---|---|---|---|---|---|
| Identifikatori | |||||||
| Simbol | C9orf91 | ||||||
| RefSeq | NP_694590.2 | ||||||
| UniProt | Q5VZI3 | ||||||
| Ostali podaci | |||||||
| Lokus | Hrom. 9 q32 | ||||||
| |||||||
Gen
[uredi | uredi izvor]Lokacija
[uredi | uredi izvor]Gen TMEM268 mapiran je na hromosomu 9, sekvenca 9q32.[6] U susjedstvu su mu doljeprikazani geni . Područje zatvoreno crvenim krugom je TMEM268', područje kodiranja koje je okruženo susjedima DFNB31 i ATP6V1G1.

iRNK
[uredi | uredi izvor]Pronađeno je osam različitih [[alternativna prerada RNK|alternativnih varijanti prerade TMEM268.[7] Egzonske sekvence za prikazane kombinacije označava kombinirana područja. Dužina aminokiselina razlikuje se zbog varijacija u dodavanju ili deletiranju nukleotida unutar egzonskih regija, dajući različite varijante prerade sa sličnim kombinacijama sekvenci.


Protein
[uredi | uredi izvor]Opća svojstva
[uredi | uredi izvor]TMEM268 ima sedam varijanti. NP_694590.2 je varijanta koja se najviše proučava. ExPasy rezultat ukazuje da TMEM268 ima izoelektričnu tačku na 5,19 i molekulsku težinu oko 37,6 kDa.[8] TMEM268 is a member of a domain of unknown function, DUF4481.[9] Regija se nalazi unutar aminokiselinskih sekvenci 37 i 328. Rezultati BLAST-a indiciraju da postoje paralozi kod ljudi, zebrica i vinskih mušica.
Aminokiselinska sekvenca
[uredi | uredi izvor]Dužina polipeptidnog lanca je 342 aminokiseline, а molekulska težina 37.569 Da.[11]
| 10 | 20 | 30 | 40 | 50 | ||||
|---|---|---|---|---|---|---|---|---|
| MACEPQVDPG | ATGPLPPSSP | GWSALPGGSP | PGWGQELHNG | QVLTVLRIDN | ||||
| TCAPISFDLG | AAEEQLQTWG | IQVPADQYRS | LAESALLEPQ | VRRYIIYNSR | ||||
| PMRLAFAVVF | YVVVWANIYS | TSQMFALGNH | WAGMLLVTLA | AVSLTLTLVL | ||||
| VFERHQKKAN | TNTDLRLAAA | NGALLRHRVL | LGVTDTVEGC | QSVIQLWFVY | ||||
| FDLENCVQFL | SDHVQEMKTS | QESLLRSRLS | QLCVVMETGV | SPATAEGPEN | ||||
| LEDAPLLPGN | SCPNERPLMQ | TELHQLVPEA | EPEEMARQLL | AVFGGYYIRL | ||||
| LVTSQLPQAM | GTRHTNSPRI | PCPCQLIEAY | ILGTGCCPFL | AR |
C: Cistein
D: Aspartat
E: Glutamat
F: Fenilalanin
G: Glicin
H: Histidin
I: Izoleucin
K: Lizin
L: Leucin
M: Metionin
N: Asparagin
P: Prolin
Q: Glutamin
R: Arginin
S: Serin
T: Treonin
V: Valin
- W: Triptofan
Y: Tirozin
Transmembranska svojstva
[uredi | uredi izvor]Dvije predviđene transmembranske regije nalaze se u polipeptidu, smještene na aminokiselinama 104-125 i 130-152 .[12] Alat SAPS na San Diego Super Computing Biology Workbench prikupio je strukturne karakteristike proteina.[13]
Hipoteza za transmembranski smjer je da N– i C-terminal ostanu unutar ćelije, a petlja koja strši iz membrane u citosol, predstavljena na susjednoj slici. Dokazi o smjeru transmembrane proizlaze iz predviđanja mjesta fosforilacije. Rezultati Net Phos 2.0 za TMEM268 ukazuju na područja fosforilacije koja su prisutna oko transmembranske regije. To znači da će N– i C kraj biti okrenuti prema ćeliji, pružajući podršku smjeru transmembrane.

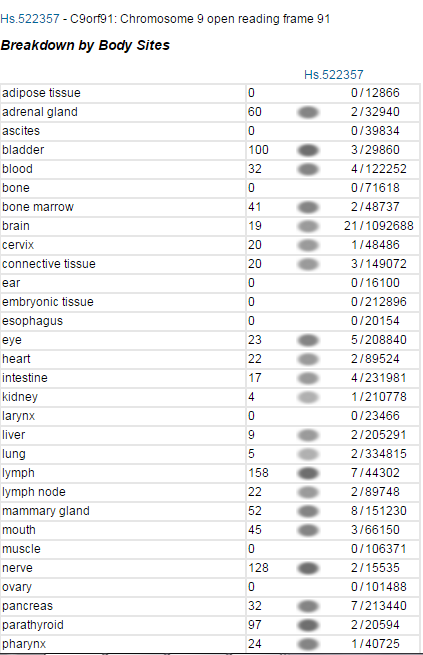
Utvrđeno je da se protein eksprimira u brojnim različitim tkivima. ETS profil za TMEM268 pokazuje da se gen eksprimira sveprisutno u tijelu.[14] U prilog tome, postoje eksperimentalni dokazi koji koriste bojenje antitijelima za prikaz različitih tipova tkiva koje eksprimiraju TMEM268.[15]
Nedavno istraživanje povezalo je TMEM268 kao lizosomskog transportera.[16] Tačna funkcija unutar lizosoma i interakcije s drugim molekulama, međutim, nije poznata.
Povezana bolest
[uredi | uredi izvor]Zbog ograničene karakterizacije, veza sa povezanim bolestima nije dobro poznata. Tehnike pregleda jednonukleotidnih polimorfizama korištene su u brojnim bolestima za određivanje utjecaja datih lokusa. Kod spondilokoštane disostoze i spondiloartritisa, za oba je utvrđeno da imaju SNP-ove koji su naveli TMEM268 kao mogući faktor bolesti [17][18]
Interaktivni proteini
[uredi | uredi izvor]Interakcija proteina pronađena je sa c-REL (putem afinitetne hromatografije i dvohibridnog kvasca) i ELAVL1, putem dvohibrida kvasaca.[19][20] REL je povezan s proto-onkogenom iz porodice transkripcijskih faktora. Povezan je s proliferacijom B-ćelija. ELAV1 je povezan sa vezivanjem za iRNK, kako bi se povećala stabilnost transkripata.
Homologija
[uredi | uredi izvor]Identitet i sličnost ortologa
[uredi | uredi izvor]Identitet ortolognih domena unutar TMEM268 kod drugih vrsta sa 40% identiteta i iznad prikazan je u nastavku. Početak DUFF4481 prikazan na slici zadržava konzerviranost tokom vremena. Zanimljivo je da područja transmembranske regije nisu toliko konzervirana kao što se mislilo, što ukazuje da određene aminokiseline možda nisu vitalna komponenta u funkciji ovog proteina.

Konzervacija unutar vrste
[uredi | uredi izvor]TMEM268 je konzerviran u mnogim organizmima. Ortolozi su pronađeni kod sisara, ptica, gmizavaca, vodozemaca, riba, uključujući morske pse i nekoliko insekata. Nije pronađeno u biljkama, bakterijama ili arhejama. Dužina aminokiselinskog lanca ima tendenciju povećanja unutar ortologa jer se divergiranje od ljudi povećava na temelju rezultata pretraživanja BLAST-a za slične proteine.
Reference
[uredi | uredi izvor]- ^ a b c GRCh38: Ensembl release 89: ENSG00000157693 - Ensembl, maj 2017
- ^ a b c GRCm38: Ensembl release 89: ENSMUSG00000045917 - Ensembl, maj 2017
- ^ "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- ^ "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- ^ "I-TASSER server for protein structure and function prediction". zhanglab.ccmb.med.umich.edu. Pristupljeno 8. 5. 2016.
- ^ mieg@ncbi.nlm.nih.gov, Danielle Thierry-Mieg and Jean Thierry-Mieg, NCBI/NLM/NIH. "AceView: Gene:C9orf91, a comprehensive annotation of human, mouse and worm genes with mRNAs or ESTsAceView". www.ncbi.nlm.nih.gov. Pristupljeno 28. 4. 2016.
- ^ "NCBI Gene".
- ^ "ExPASy isoelectric point".
- ^ "transmembrane protein C9orf91 [Homo sapiens] - Protein - NCBI". www.ncbi.nlm.nih.gov. Pristupljeno 28. 4. 2016.
- ^ a b "GlobPlot 2.3 - Predictor of intrinsic protein disorder & globularity". globplot.embl.de. Pristupljeno 28. 4. 2016.
- ^ "UniProt, Q5VZI3". Pristupljeno 12. 9. 2021.
- ^ "C9orf91 - Transmembrane protein C9orf91 - Homo sapiens (Human) - C9orf91 gene & protein". www.uniprot.org. Pristupljeno 28. 4. 2016.
- ^ "SDSC Biology Workbench". workbench.sdsc.edu. Pristupljeno 28. 4. 2016.
- ^ "EST Profile - Hs.522357". www.ncbi.nlm.nih.gov. Pristupljeno 28. 4. 2016.
- ^ "Tissue expression of C9orf91 - Summary - The Human Protein Atlas". www.proteinatlas.org. Pristupljeno 28. 4. 2016.
- ^ Chapel A, Kieffer-Jaquinod S, Sagné C, Verdon Q, Ivaldi C, Mellal M, et al. (juni 2013). "An extended proteome map of the lysosomal membrane reveals novel potential transporters". Molecular & Cellular Proteomics. 12 (6): 1572–88. doi:10.1074/mcp.M112.021980. PMC 3675815. PMID 23436907.
- ^ Zinovieva E, Kadi A, Letourneur F, Cagnard N, Izac B, Vigier A, et al. (juli 2011). "Systematic candidate gene investigations in the SPA2 locus (9q32) show an association between TNFSF8 and susceptibility to spondylarthritis". Arthritis and Rheumatism. 63 (7): 1853–9. doi:10.1002/art.30377. PMID 21480186.
- ^ Sparrow DB, McInerney-Leo A, Gucev ZS, Gardiner B, Marshall M, Leo PJ, et al. (april 2013). "Autosomal dominant spondylocostal dysostosis is caused by mutation in TBX6". Human Molecular Genetics. 22 (8): 1625–31. doi:10.1093/hmg/ddt012. PMID 23335591.
- ^ Abdelmohsen K, Srikantan S, Yang X, Lal A, Kim HH, Kuwano Y, et al. (maj 2009). "Ubiquitin-mediated proteolysis of HuR by heat shock". The EMBO Journal. 28 (9): 1271–82. doi:10.1038/emboj.2009.67. PMC 2683047. PMID 19322201.
- ^ Wang J, Huo K, Ma L, Tang L, Li D, Huang X, et al. (oktobar 2011). "Toward an understanding of the protein interaction network of the human liver". Molecular Systems Biology. 7: 536. doi:10.1038/msb.2011.67. PMC 3261708. PMID 21988832.