Ribozim

Ribozimi (ribonukleinske kiseline + enzimi) su RNK molekule koje su sposobne za katalizu specifičnih biohemijskih reakcija, slično djelovanju proteinskih enzima. Otkriće ribozima 1982. pokazalo je da RNK može biti i genetički materijal (kao DNK) i biološki katalizator (kao što je proteinski enzim) i doprinijelo postavci hipoteze RNK svijeta, koja ukazuje da RNK može biti važna u evoluciji prebiotskih sistema koji se sami razmnožavaju. Najčešće aktivnosti prirodnih ili in vitro razvijenih ribozima su cijepanje ili spajanje fragmenata RNK i DNK i formiranje peptidnih veza.[1] U ribosmima, funkcija ribozima kao djela velike podjedinice ribozomake RNK je povezivanje aminokiselina u sintezi proteina. Oni također sudjeluju u raznim reakcijama obrada transktibirane RNK, uključujući i preradu RNK, virusnu replikaciju i RNK biosinteze. Primjeri ribozima uključuju ribozim čekićar, VS ribozim, lidzim i tzv. ukosnica ribozim.[2][3][4][5][6]
Istraživači koji proučavaju porijekla života su proizveli ribozime u laboratorijama, a koji su sposobni za katalizu sopstvene sinteze od skoro monomera pod vrlo specifičnim uvjetima, kao što je RNK polimerazni ribozim.[7] Mutageneza i selekcija su rezultiraje dobijanjem i izolacijom potvrđenih varijanti "round-18" polimeraznog ribozibozima od 2001. "B6.61" može za 24 sata dodati do 20 nukleotida na primarnu matricu, dok se ne razlaže do cijepanja svojih fosfodiesterskih veza.[8] Ribozim "tC19Z" može dodati do 95 nukleotida sa tačnošću od 0,0083 mutacija/nukleotid.[9] Pokušaji su rađeni u cilju da razviju ribozime kao terapeutska sredstava, kao enzime koji ciljaju cijepanje definirane RNK sekvence , kao biosenzora te za aplikacije u finkcijskoj genomici i otkriću gena. .[10]
Otkriće
[uredi | uredi izvor]
Prije otkrića ribozima, enzimi, koji su definirani kao katalitski proteini,[11] bili su jedini poznati biološki katalizatori. U 1967., Carl Woese, Francis Crick i Leslie Orgel prvi su sugerirali da RNK može djelovati kao katalizator. Ova ideja je bila zasnovana na otkriću da RNK može formirati kompleksne sekundarne strukture.[12] Ribozime je, ranih 1980ih, otkrio Thomas R. Cech, koji je proučavao preradu RNK kod trepljarske protozoe Tetrahymena thermophila, sa Sidneyom Altmanom and Normanom Paceom, kji su proučavali kompleks bakterijske RNkaze P. Ovi ribozimi su nađeni intronu RNK transkripta, koji se od njega odstranio, kao i u komponenti RNK kompleksa RNkaze P, koji je uključen u sazrijevanje pre-tRNK. U 1989., Thomas R. Cech i Sidney Altman su podijelili Nobelovu nagradu za hemiju za njigovo "otkriće katalitskih svojstvenosti RNK."[13] Termin ribozim su prvi predstavili Kelly Kruger et al. 1982 u radu koji je publiciran u časopisu Cell.[14]
Do tada je bio čvrsto uspostavljeno vjerovanje u biologiji da je kataliza rezervirana za proteine. Međutim, ideja RNK katalize je motivirana dijelimično i od starog pitanja o poreklu života: šta se pojavilo prvo, enzim koji obavljaju funkcije ćelije ili nukleinske kiseline koje nose informacije koje su potrebne za proizvodnju enzima? Koncept "ribonukleinske kiseline kao katalizatori" zaobilazi ovaj problem. RNK, u suštini, može biti „i pile i jaje“ .
U 1980. Thomas Cech, na University of Colorado u Boulderu, je proučavao eksciziju introna u ribozomnom RNK genu u Tetrahymena thermophila. Dok je pokušavao da očisti enzim odgovoran za reakciju prerade RNK, otkrio je da je introni mogu biti spojeni u odsustvu dodatka ekstrakta ćelija. Koliko god su pokušavali, Cech i njegove kolege nisu mogli identificirati bilo koji protein povezan s reakcijom prerade. Nakon mnogo posla, Cech je predložio da intronska sekvenca dio RNK može razložiti i reformirati fosfodiesterske veze. Otprilike u isto vrijeme, Sidney Altman, profesor na Yale University, je proučavao način kako se tRNK molekule se obrađuju u ćeliji, kada su on i njegove kolege izolovali enzim RNkaza-P, koji je odgovoran za konverziju prekursorne tRNK u aktivnu tRNK. Na njihovo veliko iznenađenje, otkrili su da je u RNK postoji RNkaza-P, osim proteina i da je RNK bitna komponenta aktivnog enzima. To je bio tako strana ideja da su imali poteškoća da objave svoje nalaze. Naredne godine, Altman je pokazao da RNK može djelovati kao katalizator, dokazujući da je RNkaza-P individualne RNK mogla katalizirati cijepanje prekursora tRNK u aktivnu tRNK, i u odsustvu proteinske komponente.
Od otkrića Cecha i Altmana, drugi istražitelji su otkrili druge primjere samoobnavljajuće RNK ili katalizirajućih molekula RNK. Mnogi ribozimi imaju ili tzv. ukosnički ili čekićasti oblik aktivnog centrfa i jedinstvenu sekundarnu strukturu koja im omogućava da RNK razlaže druge molekule na specifične sekvence. To je sada moguće napraviti ribozimima da će se posebno razložiti bilo koja molekula RNK. Ovi RNK katalizatori mogu imati farmaceutske aplikacije. Naprimjer, dizajniran je ribozim koji zazlaže RNK HIV-a. Ako bi takve ribozime pravile same ćelije, sve dolazne virusne čestice će doći u situaciju da njihov RNK genom razlože ribozimi, što bi spriječilo infekciju.
Struktura i mehanizam
[uredi | uredi izvor]Unatoč tome, što postoje samo četiri izbora za svaku monomernu jedinicu (nukleotidi), u odnosu na 20 aminokiselina koje se nalaze u bočnim lancima proteina, ribozimi imaju različite strukture i mehanizme. U mnogim slučajevima oni su u mogućnosti da oponašaju mehanizam koji s koriste njihovi proteinski oblici. Naprimjer, u samokatalizi ribozima RNK, SN2 reakcija se provodi uz pomoć 2 'hidroksilnske grupe kao nukleofil koji napada fosfatne veze i pretvaraju 5' kisik u N + 1 bazu i ponašaju se kao napušta grupu. U odnosu na RNkazu A, protein koji katalizira istu reakciju, koristi koordinaciju histidina i lizina da djeluje kao baza za napad na fosfatnu okosnicu. Kao i kod mnogi drugih proteinskih enzima. Često ove interakcije koristiti i fosfatna kičma i baze nukleotida, uzrokujući drastične konformacijske promjene.[15][16]
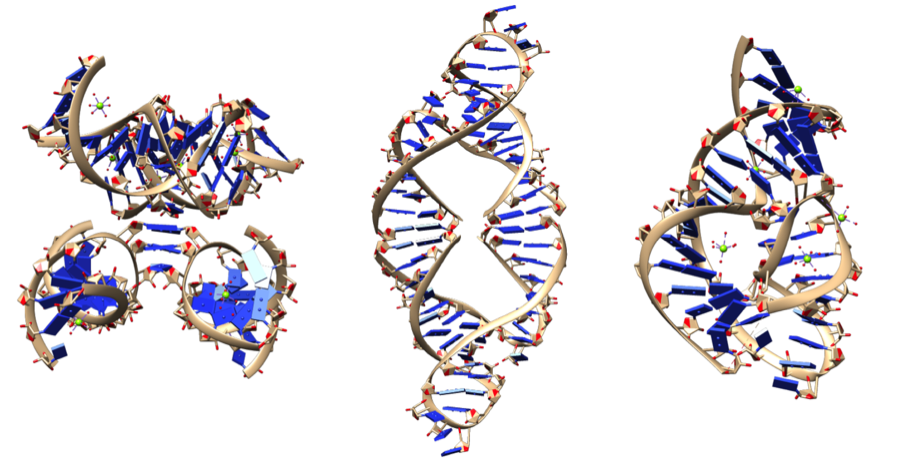
S lijeva na desno: lidzim, čekićasti ribozim i uvijeni („ukosnica“) ribozim
Aktivnost
[uredi | uredi izvor]Iako je većina ribozima sasvim rijetka u ćeliji, njihova uloga je ponekad esencijalna za život. Naprimjer, funkcijski dio ribosom, molekulske mašine koja prevodi iRNK u proteine, fundamentalno je ribozim, sastavljen od tercijarnih strukturnih motiva RNK koji su često koordinirani metalnim ionima, kao što je Mg2+, kao kofatorima. U model sistemima nema prihvatanja divalentnih kationa u peto-nukleotidnu RNK katalizirajući trans-fenilalanaciju četvero-nukleotidnog supstrata sa 3 komplementarna bazna para sa katalizatorom, gdje je katalizator/supstrat osmislio skraćivanje C3 ribozima.[17] RNK može katalizirati savijanje proteina patološke proteinske konformacije priona, na način koji mje sličan haperoninu,[18] i može biti uključen u virusnu konkatamerno cijepanje koji prethodi pakovanju virusnog genetičkog materijala u nekim virionima.
RNK također može djelovati kao nasljedne molekula, što je ohrabrilo Waltera Gilberta da predloži da se u dalekoj prošlosti, ćelije koristile RNK kao i genetički materijal i strukturne i katalitske molekule radije nego odvijanje ove funkcije između DNK i proteina, kao što je danas; ova hipoteza je poznat kao "hipoteza RNK svijeta" u tumačenju porijekla života. Dokaz da su ribozimi bili prve molekulske mašine ranog života ukazuje na to da su oni u stvari "molekulski fosili".[19]
Vještački ribozimi
[uredi | uredi izvor]Od otkrića ribozima koj postoje u živim organizmima, došlo je interesovanja za istraživanje novih sintetičkih ribozima je u laboratorijama. Naprimjer, umjetno je proizvedena samo-obnavljajuća RNK koja ima dobru enzimsku aktivnost. Tang i Breaker su izolirali samo-replikacijsku RNK in vitro iz iz slučajnih sekvenci RNK. Neki od sintetičkih ribozima, koji su proizvedeni imli su duge strukture, a neki su bili slični prirodnim čekićastim ribozimima. U 2015. godini, istraživači u Northwestern University i University of Illinois, u Chicagu su projektovali ograničeni ribosom, koji djeluje gotovo kao i autentične ćelijske komponente koje proizvode sve proteine i enzime unutar ćelije. Nazvan je ribosom-T ili ribo-T, umjetni ribosom, a kreirali su ga jMichael Jewett i Alexander Mankin. Prvi umjetni ribosom – dizajnirani ribosom, koji bi mogao dovesti do novih lijekova i nove generacije biomaterijala, objavljen je 31. jula 2015. na Northwestern University Tehnike koje se koriste za stvaranje veštačkih ribozima uključuju usmjerenu evoluciju. Ovaj pristup koristi prednosti dualne prirodi RNK, i kao katalizator i informacijskog polimera, što ga čini lahkim za istraživanje proizvodnje ogromne populacije RNK katalizatora, pomoću polimeraznih enzima. Ribozimi su mutirali po obrnutoj transkripciji reverzhnom transkriptazom u različite cDNK i pojačani PCRom. Izbor parametara u ovim eksperimentima često se razlikuju. Jedan od pristupa za izbor ligaznog ribozima koji uključuje tragove biotina, koji su kovalentno povezani sa podlogom. Ako molekula posjeduje željenu ligaznu aktivnost, za oporavak aktivne molekule može se koristiti streptavidinska matrica.
Lincoln i Joyce su razvili RNK enzimski sistem sposoban za samoreplikaciju za oko sat vremena. Koristeći molekulsku konkurenciju ( in vitro evolucija) kandidata RNK smjese, pojavio se par ribozima, u kojem se svaki spaja s druge strane spajanja sintetičkih oligonukleotida, bez prisustva proteina.
Iako nisu istinski katalizatori, stvaranje umjetnih ribozima je nazvano riboupaljačima (riboswichers) samoreplikacije, što je također bilo aktivno područje istraživanja.Riboupaljači su regulatorni RNK motivi koji mijenjaju svoju strukturu kao odgovor na ligande malih molekula za reguliranje translacije. Iako postoje mnogi poznati prirodni riboupaljači, koji vežu široku lepezu metabolita i drugih malih organskih molekula, opisan je samo jedan ribozim kao riboupaljač, glmS . ]].[20] Rani rad u karakterišu samo-paranje riboswitches je fokusirana na pomoću teofilin kao ligand. U ovim studijama formirana je „RNK ukosnica“ koji blokira ezanje ribosomskog mjesta, čime se translacija inhibira . U prisustvu liganda, u ovim slučajevima teofilina, region cijepanja regulacijske RNK , omogućava ribosomima da se vežu i prevedu ciljni gen. Mnogo toga u RNK inženjerstvu je bazirano na racionalnom dizajnu i ranije utvrđenoj strukturi RNK, a ne usmjerenom evolucijom, kao u gornjim primjerima. Noviji rad je proširio liganada koji se koriste u ribozimskom riboupaljkačima, uključujući timin pirofosfat. U inženjerstvu aptozimama koristi se također i fluorescentna activacija sortiranja ćelija (protočna citometrija).[21][22]
Primjena
[uredi | uredi izvor]Ribozimi su predloženi i razvijeni za liječenje bolesti putem genske terapije. Jedan od glavnih izazova korištenja RNK enzima u terapiji je kratak poluživot katalitske molekule RNK u tijelu. Za borbu protiv ovog, stav 2' na ribozi je modificiran da se poboljša stabilnost RNK. Jedno područje ribozima za genske terapije bilo je inhibicija RNK-baziranih virusa. Neki sintetski ribozimi su usmjereni protiv HIV-a. Razvijena je RNK nazvna „genske škare“ i ušla je kliničko testiranje na infekciju HIV-om.
Slično tome, dizajniran je ribozim da cilja na RNK virusa hepatitisa C . Ribozim je u stanju da cijepa konzervirani regiona virusnog genoma, za koji je pokazalo da reducira virus u sisarskoj kulturi ćelija. Uprkos ovim naporima istraživača, ovi projekti su ostali u pretkliničkoj fazi.
Poznati ribozimi
[uredi | uredi izvor]Potvrđene klase prirodnih ribozima:
- GIR1 granati ribozim[23]
- GlmS ribozim
- Grupa I samoprerađeni introni
- Grupa II. Samoprerađujući intron - splajsosom je vjerovatno izveden iz grupe II samoprerađujućih ribozima.
- Ukosnica ribozim
- Čekićasti ribozim
- HDV ribozim
- rRNK* – nalazi se u svim živim ćelijama i vežu aminokiseline da formiraju proteine.
- RNkaza P
- Twister ribozim
- VS ribozim.[24]
Također pogledajte
[uredi | uredi izvor]Reference
[uredi | uredi izvor]- ^ Fedor M. J., Williamson J. R. (2005). "The catalytic diversity of RNAs". Nat. Rev. Mol. Cell Biol. 6: 399–412. doi:10.1038/nrm1647.
- ^ Hadžiselimović R., Pojskić N. (2005): Uvod u humanu imunogenetiku. Institut za genetičko inženjerstvo i biotehnologiju (INGEB), Sarajevo, ISBN 9958-9344-3-4.
- ^ Alberts B. (2002)ː Molecular biology of the cell. Garland Science, New York, ISBN 0-8153-3218-1.
- ^ Bajrović K, Jevrić-Čaušević A., Hadžiselimović R., Ed. (2005): Uvod u genetičko inženjerstvo i biotehnologiju. Institut za genetičko inženjerstvo i biotehnologiju (INGEB), Sarajevo, ISBN 9958-9344-1-8.
- ^ Kapur Pojskić L., Ed. (2014): Uvod u genetičko inženjerstvo i biotehnologiju, 2. izdanje. Institut za genetičko inženjerstvo i biotehnologiju (INGEB), Sarajevo, ISBN 978-9958-9344-8-3.
- ^ Hall J. E., Guyton A. C. (2006): Textbook of medical physiology, 11th edition. Elsevier Saunders, St. Louis, Mo, ISBN 0-7216-0240-1.
- ^ Johnston WK, Unrau PJ, Lawrence MS, Glasner ME, Bartel DP (maj 2001). "RNA-catalyzed RNA polymerization: accurate and general RNA-templated primer extension". Science. 292 (5520): 1319–1325. Bibcode:2001Sci...292.1319J. doi:10.1126/science.1060786. PMID 11358999.CS1 održavanje: više imena: authors list (link)
- ^ Zaher HS, Unrau PJ (juli 2007). "Selection of an improved RNA polymerase ribozyme with superior extension and fidelity". RNA. 13 (7): 1017–1026. doi:10.1261/rna.548807. PMC 1894930. PMID 17586759.
- ^ Wochner A, Attwater J, Coulson A, Holliger P (april 2011). "Ribozyme-catalyzed transcription of an active ribozyme". Science. 332 (6026): 209–212. Bibcode:2011Sci...332..209W. doi:10.1126/science.1200752. PMID 21474753.CS1 održavanje: više imena: authors list (link)
- ^ Hean J, Weinberg MS (2008). "The Hammerhead Ribozyme Revisited: New Biological Insights for the Development of Therapeutic Agents and for Reverse Genomics Applications". u Morris KL (ured.). RNA and the Regulation of Gene Expression: A Hidden Layer of Complexity. Norfolk, England: Caister Academic Press. ISBN 1-904455-25-5. Upotreblja se zastarjeli parametar
|chapterurl=(pomoć)CS1 održavanje: nepreporučeni parametar (link) - ^ Enzyme definition Dictionary.com Accessed 6 April 2007
- ^ Carl Woese, The Genetic Code (New York: Harper and Row, 1967).
- ^ [1]
- ^ Kruger K, Grabowski PJ, Zaug AJ, Sands J, Gottschling DE, Cech TR (novembar 1982). "Self-splicing RNA: autoexcision and autocyclization of the ribosomal RNA intervening sequence of Tetrahymena". Cell. 31 (1): 147–57. doi:10.1016/0092-8674(82)90414-7. PMID 6297745.CS1 održavanje: više imena: authors list (link)
- ^ Pyle A. M. (1993). "Ribozymes: a distinct class of metalloenzymes". Sci. 261: 709–714. doi:10.1126/science.7688142. PMID 7688142.
- ^ Freisinger E., Sigel R. K. O. (2007). "From nucleotides to ribozymes—A comparison of their metal ion binding properties". Coord. Chem. Rev. 251: 1834–1851. doi:10.1016/j.ccr.2007.03.008.
- ^ Turk RM, Chumachenko NV, Yarus M (mart 2010). "Multiple translational products from a five-nucleotide ribozyme". Proc. Natl. Acad. Sci. U.S.A. 107 (10): 4585–4589. Bibcode:2010PNAS..107.4585T. doi:10.1073/pnas.0912895107. PMC 2826339. PMID 20176971.CS1 održavanje: više imena: authors list (link)
- ^ Supattapone S (juni 2004). "Prion protein conversion in vitro". J. Mol. Med. 82 (6): 348–356. doi:10.1007/s00109-004-0534-3. PMID 15014886.
- ^ Gilbert W (1986). "Origin of life: The RNA world". Nature. 319 (6055): 618. doi:10.1038/319618a0.
- ^ Winkler, W. C., Nahvi, A., Roth, A., Collins, J. A. & Breaker, R. R. Control of gene expression by a natural metabolite-responsive ribozyme. 4–9
- ^ Lincoln TA, Joyce GF (februar 2009). "Self-sustained replication of an RNA enzyme". Science. 323 (5918): 1229–1232. Bibcode:2009Sci...323.1229L. doi:10.1126/science.1167856. PMC 2652413. PMID 19131595.
- ^ Lynch , Gallivan J. P. (2009). "A flow cytometry-based screen for synthetic riboswitches". Nucleic Acids Res. 37: 184–92. doi:10.1093/nar/gkn924.
- ^ Nielsen H., Westhof E., Johansen S. (2005): An mRNA is capped by a 2', 5' lariat catalyzed by a group I-like ribozyme. Science, 309, 5740: 1584–1587.pmid=16141078 |doi=10.1126/science.1113645.
- ^ Astrid Sigel, Helmut Sigel and Roland K. O. Sigel, ured. (2011). Structural and catalytic roles of metal ions in RNA. Metal Ion in Life Sciences. 9. Cambridge, U.K.: RSC Publishing. doi:10.1039/9781849732512. ISBN 978-1-84973-251-2.
Vanjski linkovi
[uredi | uredi izvor]- http://www.ibiology.org/ibiomagazine/issue-1/tom-cech-discovering-ribozymes.html Arhivirano 4. 11. 2013. na Wayback Machine Tom Cech kratki govor: "Otkrivanje Ribozymes"