Ubikvitin
| Ubikvitinska familija | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Dijagram ubikvitina. Sedam lizinskih bočnih lanaca je obojeno narandžasto. | |||||||||
| Identifikatori | |||||||||
| Simbol | ubiquitin | ||||||||
| Pfam | PF00240 | ||||||||
| InterPro | IPR000626 | ||||||||
| PROSITE | PDOC00271 | ||||||||
| SCOP | 1aar | ||||||||
| SUPERFAMILY | 1aar | ||||||||
| |||||||||
Ubikvitin je mali regulatorni protein, koji je prisutan u skoro svim tkivima eukariota. Ubikvitinacija je proces posttranslacione modifikacije nekog proteina, koja nastaje njegovim vezivanjem za jedan ili više ubikvitinskih monomera kovalentnim vezama.[1] Najčešća svrha ubikvitinacije je označavanje proteina za razgradnju. Ubikvitinska etiketa usmerava proteine ka proteasomu, koji je veliki proteinski kompleks u ćelijama koji razlaže i reciklira nepotrebne proteine. Osim toga, ubikvitinacija kontroliše stabilnost, funkciju i unutrašnji ćelijski položaj velikog broja proteina. Proces ubikvitinacije započinje enzim E1.[2]
Identifikacija
[уреди | уреди извор]
Ubikvitin (prvotno sveprisutni imunopotentni polipeptid) je bio identifikovan 1975. godine kao sveprisutni protein mase 8.5 kDa. Osnovne uloge ubikvitina otkrivene su u ranim 1980-im u istraživanjima Foks Čejs instituta za rak, a otkrila ga je ekipa naučnika koju su sačinjavali Aron Ciechanover, Avram Hershko i Irvin Rose, koji su za svoje otkriće dobili Nobelovu nagradu za hemiju 2004. godine.[3][4][5]
Ubikvitinski sistem je prvotno okarakterisan kao ATP-zavisni proteolitički sistem prisutan u ćelijskim ekstraktima. Za termostabilani polipeptid prisutan u ovim ekstraktima, ATP-zavisni proteolitički faktor 1 (APF-1), je utvrđeo da je povezan kovalentnom vezom za model supstrata lizozima, u ATP i Mg2+ zavisnim procesima. Višestruki APF-1 molekuli su bili povezani s jednim molekulom supstrata izopeptidnim vezama. Ustanovljeno je da se njihovi konjugovani proteini brzo razlažu uz oslobađanje slobodnih APF-1. Ubrzo nakon toga, razjašnjena je konjugacija APF-1 proteina, koji je nazvan ubikvitinom. Karboksilna grupa aminokiseline glicina (Gly76) je otkrivena kao mjesto spajanja sa lizinom supstrata.
Protein
[уреди | уреди извор]| Broj aminokiselina | 76 |
| Molekulska masa | 8564.47 Da |
| Izoelektrično polje (pI) | 6.79 |
| Nazivi gena | RPS27A (UBA80, UBCEP1), UBA52 (UBCEP2), UBB, UBC |
Ubikvitin je protein koji se sastoji od 76 aminokiselina i ima molekulsku masu od 8.5 kDa. Ključne karakteristike su C-terminalni rep i 7 lizina. Visoko je konzerviran među eukariotskim vrstama: ljudi i kvasci dele 96% sekvence. Sekvenca ljudskog ubikvitina, pri čemu svako slovo označava aminokiselinu, je:
| MQIFVKTLTGKTITLEVEPSDTIENVKAKIQDKEGIPPDQQRLIFAGKQLEDGRTLSDYNIQKESTLHLVLRLRGG |
Ubikvitinacija
[уреди | уреди извор]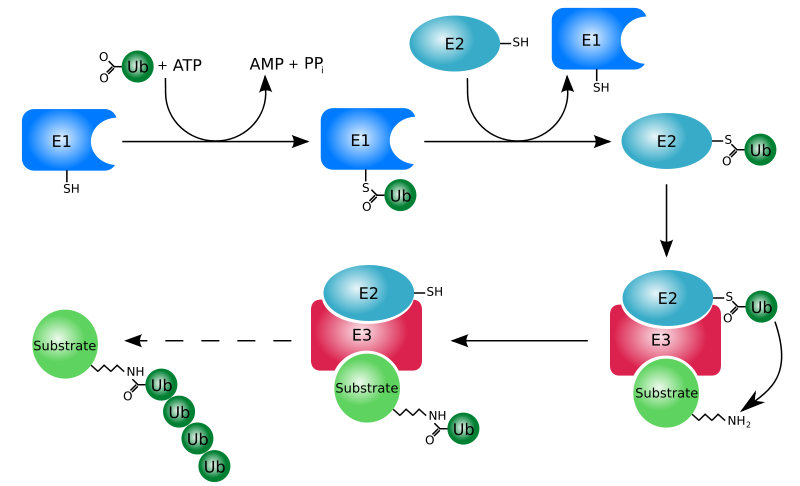
Proces označavanja nekog proteina ubikvitinom (ubikvitinacija) se sastoji od serije koraka:
- Aktivacija ubikvitina: Ubikvitin aktivira E1 ubikvitin-aktivirajući enzim u procesu koji zahtijeva ATP kao izvor energije. Prvi korak uključuje proizvodnju ubikvitin-adenilata. Druga faza je prenos ubikvitina do aktivnog mjesta, cisteina na E1 enzimu, uz otpuštanje AMP-a. Ovaj korak rezultuje u tioesterskoj vezi između C-terminalne karboksilne grupe ubikvitina i sulfihidrilnom grupom cisteina na enzimu E1.
- Prenos ubikvitina s E1 do aktivnog mesta cisteina ubikvitin-konjugujućeg enzima E2 putem tran(tio)esterifikacijske reakcije. Genomi sisara imaju 30 - 40 ubikvitin-konjugujućih enzima.
- Završni korak reakcije ubikvitinacije je stvaranje izopeptidne veze između lizina ciljanog proteina i C-terminalnog glicina ubikvitina. Ovaj korak u reakciji zahteva aktiviranje jednog od stotina E3 ubikvitin-protein ligaza (ili skraćeno, ubikvitin ligaza). E3 enzimi deluju kao supstrat za prepoznavanje i imaju sposobnost reagovanja sa E2 i sa supstratom.
U reakciji ubikvitinacije E1 se može povezati sa desetinama E2, koji se mogu povezati sa stotinama E3, hijerarhijski. Ostali ubikvitinu-slični proteini takođe reaguju preko E1-E2-E3 kaskade.
E3
[уреди | уреди извор]E3 enzimi poseduju jedan od dva moguća domena:
- HECT (engl. Homologous to the E6-AP Carboxyl Terminus) domen
- RING (engl. Really Interesting New Gene) domen
Prenos je moguć na dva načina:
- Direktno s E2, preko katalize RING domena E3 enzima
- Preko E3 enzima, katalizom HECT domena E3. U ovom slučaju, intermedijar kovalentnog E3-ubikvitina se stvara pre prebacivanja na supstrat.
Funkcije i načini vezivanja
[уреди | уреди извор]Osim da se samo jedan ubikvitin veže na supstrat, moguće je vezivanje niza molekula ubikvitina, pa tako nastaje poliubikvitinski lanac. Osim toga, neki supstrati su modifikovaani adicijom ubikvitinskih modula s lizinskim ostatkom, u procesu koji se naziva multiubikvitinacija. Kao što je već rečeno, ubikvitin ima 7 lizinskih ostataka. Istorijski, prvobitni tip prepoznatih ubikvitinskih lanaca su bili oni povezani preko lizina 48. Novija istraživanja su otkrila varijetet veza koji uključuje sve moguće lizinske nastavke, a osim toga i linearne lance povezane s N-terminalom ubikvitinskog molekula.[6][7] Radovi izdani 2007. godine pokazuju stvaranje razgranatih lanaca ubikvitina putem različitih veza.[8][9] "Atipični" (nelizinski vezani) lanci ubikvitina se spominju u radu naučnik Ikede i Đikića.[10]
Ubikvitinacijski sistem učestvuje u brojnim ćelijskim procesima, kao što su[11]:
- Razgradnja antigena
- Apoptoza
- Stvaranje organela
- Ćelijski ciklus i deoba
- Transkripciju DNK i njen popravak
- Diferencijacija i razvoj
- Imunski odgovor i upale
- Živčana i mišićna degeneracija
- Morfogeneza živčanih ćelija
- Modulacija receptora na površini ćelija i jonskih kanala
- Odgovor na stres i spoljašnje činioce
- Stvaranje ribozoma
- Virusne infekcije
Lanci povezani preko 48-og lizina
[уреди | уреди извор]

Najistraživaniji poliubikvitinski lanci, oni povezani preko 48-og molekule lizina, označavaju proteine za razgradnju u procesu koji se zove proteoliza. Barem četiri ubikvitinska molekula moraju biti povezana za lizinske nastavke ciljanog proteina kako bi ga prepoznao 26S-proteasom.[12]. Proteasom je kompleksna, bačvasta struktura s dve komore, u kojima se odvija proteoliza. Proteini se brzo razgrađaju u manje peptide (obično duge od 3 - 24 aminokiseline). Ubikvitinski molekuli se odvajaju od proteina pre same razgradnje i recikliraju za dalju upotrebu. Iako je većina proteosomskih supstrata ubikvitinirana, postoje primeri neubikvitiniranih proteina za razgradnju u proteasomu.
Monoubikvitinacija
[уреди | уреди извор]Ubikvitin takođe može da označava membranske proteine (na primer, receptore) koje treba odstraniti s membrane, i vrši nekoliko signalnih uloga u ćeliji. Molekuli na ćelijskoj membrani označeni s ubikvitinom su često monoubikvitinizirani, i ove modifikacije često označavaju protein za razgradnju u lizosomu.
Povezanost s bolestima i poremećajima
[уреди | уреди извор]Genetički poremećaji
[уреди | уреди извор]Neki genetički poremećaji često povezani s ubikvitinom su:
- Gen čija disfunkcija uzrokuje Angelmanov sindrom, UBE3A, nosi na sebi kod za ubikvitin ligazu (E3) enzmin pod nazivom E6-AP.
- Gen poremećen Von Hipel-Lindauvim sindromom kodira ubikvitin E3 ligazu pod nazivom VHL tumorski supresor ili VHL gen.
- Gen poremećen Lidlovim sindromom dovodi do disregulacije epitelnog Na+ kanala (ENaC) i uzrokuje hipertenziju.
- Osam od trinaest identifikovanih gena čija disfunkcija uzrokuje Fanconijevu anemiju kodira proteina koji formira veliki ubikvitin ligaza (E3) kompleks.
- Mutacije Cullin7 E3 ubikvitin ligaze su povezane s 3-M sindromom, recesivnim poremećajem koji rezultuje u retardaciji.[13]
Imunohistohemija
[уреди | уреди извор]Antitijela za ubikvitin se koriste u histologiji kako bi se identifikovale abnormalne nakupine proteina unutar ćelije, koje su markeri bolesti. Ove nakupine se nazivaju inkluzijska telešca. Primeri za takve nenormalne inkluzije su:
- Neufobrilarna klupka u Alzheimerovoj bolesti
- Levijeva telešca u Parkinskonovoj bolesti
- Pickova telešca u Pickovoj bolesti
- Inkluzije u bolesti motornih neurona i Huntingtonovoj bolesti
- Malorijeva telešca u alkoholnoj bolesti jetre
- Rosenthalska vlakna u astrocitima
Literatura
[уреди | уреди извор]- ^ Kerscher O, Felberbaum R, Hochstrasser M (2006). „Modification of proteins by ubiquitin and ubiquitin-like proteins”. Annual Review of Cell and Developmental Biology. 22: 159—80. PMID 16753028. doi:10.1146/annurev.cellbio.22.010605.093503.
- ^ Pickart CM (2001). „Mechanisms underlying ubiquitination”. Annual Review of Biochemistry. 70: 503—33. PMID 11395416. doi:10.1146/annurev.biochem.70.1.503. Приступљено 07. 04. 2011.
- ^ „The Nobel Prize in Chemistry 2004”. Nobelprize.org. Приступљено 16. 10. 2010.
- ^ „The Nobel Prize in Chemistry 2004: Popular Information”. Nobelprize.org. Архивирано из оригинала 15. 10. 2010. г. Приступљено 16. 10. 2010.
- ^ „Official website of Nobel Prize Commitee, list of 2004 winners”. Приступљено 30. 04. 2008.
- ^ Ping, Xu; Peng, Junmin (01. 05. 2008). „Characterization of polyubiquitin chain structure by middle-down mass spectrometry”. Analytical Chemistry. 80 (9): 3438—44. doi:10.1021/ac800016w.
- ^ Junmin, Peng; Schwartz, Daniel; Joshua E Elias; Carson C Thoreen; Cheng, Dongmei; Marsischky, Gerald; Roelofs, Jeroen; Finley, Daniel; Steven P Gygi (2003). „A proteomics approach to understanding protein ubiquitination”. Nature Biotechnology. 21 (8): 921—6. doi:10.1038/nbt849.
- ^ Takayoshi, Kirisako; Kamei, Kiyoko; Murata, Shigeo; Kato, Michiko; Fukumoto, Hiromi; Kanie, Masato; Sano, Soichi; Tokunaga, Fuminori; Tanaka, Keiji; Kazuhiro Iwai (18. 10. 2006). „A ubiquitin ligase complex assembles linear polyubiquitin chains”. The EMBO Journal. 25 (20): 4877—87. doi:10.1038/sj.emboj.7601360.
- ^ Kim Hyoung Tae; Kwang Pyo Kim; Lledias, Fernando; Alexei F Kisselev; K Matthew Scaglione; Skowyra, Dorota; Steven P Gygi; Alfred L Goldberg (15. 06. 2007). „Certain pairs of ubiquitin-conjugating enzymes (E2s) and ubiquitin-protein ligases (E3s) synthesize nondegradable forked ubiquitin chains containing all possible isopeptide linkages”. The Journal of Biological Chemistry. 282 (24): 17375—86. doi:10.1074/jbc.M609659200.
- ^ Fumiyo, Ikeda; Dikic, Ivan (2008). „Atypical ubiquitin chains: new molecular signals. 'Protein Modifications: Beyond the Usual Suspects' review series”. EMBO Reports. 9 (6): 536—42. doi:10.1038/embor.2008.93.
- ^ „Ubiquitin Proteasome Pathway Overview”. Архивирано из оригинала 30. 03. 2008. г. Приступљено 30. 04. 2008.
- ^ JS Thrower, L Hoffman, M Rechsteiner, C M Pickart (04. 01. 2000). „Recognition of the polyubiquitin roteolytic signal”. The EMBO Journal. 19 (1): 94—102. doi:10.1093/emboj/19.1.94.
- ^ Huber Céline, Dora Dias-Santagata, Anna Glaser, James O'Sullivan, Raja Brauner, Kenneth Wu, Xinsong Xu, Kerra Pearce, Rong Wang, Maria Luisa Giovannucci Uzielli, Nathalie Dagoneau, Wassim Chemaitilly, Andrea Superti-Furga, Heloisa Dos Santos, André Mégarbané, Gilles Morin, Gabriele Gillessen-Kaesbach, Raoul Hennekam, Ineke Van der Burgt, Graeme C M Black, Peter E Clayton, Andrew Read, Martine Le Merrer, Peter J Scambler, Arnold Munnich, Zhen-Qiang Pan, Robin Winter, Valérie Cormier-Daire (2005). „Identification of mutations in CUL7 in 3-M syndrome”. Nature Genetics. 37 (10): 1119—24. doi:10.1038/ng1628.
Spoljašnje veze
[уреди | уреди извор]- Ubiquitin на US National Library of Medicine Medical Subject Headings (MeSH)
- Angelmanov sindrom
- OMIM
- „UniProt”. Архивирано на сајту Wayback Machine (24. март 2006)
- „UbiPred”.
- „Ubikvitin”.
- 7.340 Ubikvitinacija
| Државне | |
|---|---|
| Остале | |