Imunski sistem

Imúnski sistém je organski sistem, sestavljen iz specializiranih celic, organov in procesov, ki nadzorujejo organizem in ga varujejo pred patogeni. Ključna značilnost imunskega sistema je sposobnost razločevanja med telesu lastnimi in tujimi molekulami, ki so izražene na površini celic ali plavajo v medceličnini, pri čemer v normalnih okoliščinah sprožijo imunski odziv samo tuje. Tistim molekulam, ki sprožijo imunski odziv tako, da se vežejo na specifične imunske receptorje, pravimo antigeni.[1] Pogosti so na bakterijah, virusih, zajedavcih in glivah, antigen pa je lahko tudi zdravilna učinkovina v zdravilu, pelod, različne kemikalije, v določenih primerih pa tudi človeku lastne molekule.
Pri mnogih vrstah organizmov lahko imunski sistem razdelimo na podzvrsti, in sicer na prirojeni in pridobljeni imunski sistem. Imunost, ki jo zagotavlja imunski sistem, lahko razdelimo tudi na celično in humoralno; prvo posredujejo neposredno določene imunske celice, drugo pa protitelesa v telesnih tekočinah.[2]
V človeku predstavljajo krvno-možganska pregrada, krvno-likvorska pregrada in podobne pregrade na meji med telesnimi tekočinami in možgani ločnico med perifernim imunskim sistemom in nevroimunskim sistemom, ki varuje možgane.
Patogeni so sposobni hitrega razvoja in prilagoditev, zaradi česar se lahko izognejo odkritju in nevtralizaciji, ki jo sproži imunski sistem, a se je vseeno razvilo mnogo obrambnih mehanizmov, ki patogene prepoznajo in onesposobijo. Tudi enostavni enocelični organizmi posedujejo osnovna orodja v obliki encimov, ki služijo kot zaščita pred okužbo z bakteriofagi. Drugi osnovni mehanizmi so se razvili v starodavnih evkariontih in se ohranili v njihovih potomcih, današnjih rastlinah in nevretenčarjih. Te mehanizmi so fagocitoza, protimikrobni peptidi (defenzini) in sistem komplementa. Vretenčarji s čeljustjo (Gnathostomata), med katere spada tudi človek, imajo še bolj zapleten mehanizem obrambe, saj se lahko imunski sistem skozi čas prilagaja, da učinkoviteje prepozna patogene. Pridobljeni (specifični) imunski sistem ima sposobnost ohranjanja imunskega spomina po prvem odzivu na specifičen patogen. Ob vsakem naslednjem srečanju z istim patogenom je odziv okrepljen. Ta proces pridobljene imunosti je osnova cepljenja.
Okvare imunskega sistema lahko vodijo v avtoimunske bolezni, vnetja in rakave spremembe. O imunski oslabljenosti govorimo, ko je aktivnost imunskega sistema zmanjšana, kar vodi v ponavljajoče se in življenju nevarne okužbe. Pri človeku je posledica genetske bolezni, kot je huda kombinirana imunska pomanjkljivost (SCID), pridobljenih stanj, kot je HIV/aids, ali uporabe imunosupresivnih zdravil. Avtoimunost je po drugi strani posledica prevelike aktivnosti imunskega sistema, ki napada normalno tkivo, ki ga prepoznava kot tujek. Med pogostejša takšna stanja spada Hašimotov tiroiditis, revmatoidni artritis, sladkorna bolezen tipa 1 in lupus.
Z imunskim sistemom se znanstveno ukvarja imunologija, ki je tradicionalno panoga medicine; zaradi pomena za zdravje je najbolj raziskan človeški imunski sistem, vendar imajo pri odkrivanju njegovega delovanja ključno vlogo tudi poskusi na modelnih živalskih vrstah.
Zgodovina raziskav
[uredi | uredi kodo]Imunologija je veda, ki preučuje sestavo in naloge imunskega sistema. Izvira iz medicine in zgodnjih študij vzrokov za imunost pred boleznimi. Najstarejša omemba imunosti je iz časa atenske kuge leta 430 pr. n. št. Tukidid je zapisal, da so preboleli iz prejšnjih izbruhov lahko skrbeli za obolele brez tveganja ponovitve bolezni.[3] V 18. stoletju je Pierre-Louis Moreau de Maupertuis eksperimentiral s strupom škorpijonov in opazil, da so nekateri psi in miši nanj odporni.[4] To in podobna opažanja je kasneje izrabil Louis Pasteur pri razvoju cepiva in teorije, da so povzročitelji mnogih bolezni mikroorganizmi,[5] kar je nasprotovalo do tedaj razširjenemu prepričanju, da je za bolezni odgovorna miazma (starogrško »onesnaženje«, »madež«), strupena oblika zraka, tudi »slab/nočni zrak«. Šele ko je Robert Koch leta 1891 izvedel svoje znamenite poskuse, so bili mikroorganizmi splošno prepoznani kot vzrok nalezljivih bolezni.[6] Z Reedovim odkritjem virusa rumene mrzlice leta 1901 so spoznali, da lahko bolezni povzročajo tudi virusi.[7]
Proti koncu 19. stoletja je pričela imunologija bliskovito napredovati z odkritji na področjih humoralne in celične imunosti.[8] Še posebej pomembno je delo Paula Ehrlicha, ki je s teorijo stranskih verig pojasnil specifičnosti reakcije med antigenom in protitelesom, ter Ilje Mečnikova, ki velja za očeta celične imunologije. Za svoje delo sta leta 1908 prejela Nobelovo nagrado za fiziologijo ali medicino.[9] Za prelomna odkritja na področju imunologije je bilo skupno podeljenih že 11 Nobelovih nagrad za fiziologijo ali medicino, vključno s prvo, ki jo je prejel Emil von Behring za odkritje antiseruma proti toksinom tetanusa v krvi živali, okuženih s to boleznijo.[10]
Slojevita organizacija
[uredi | uredi kodo]Imunski sistem je organiziran v sloje z naraščajočo stopnjo specifičnosti proti patogenom. Najosnovnejša obramba organizma je njegova zunanja plast (integument), ki predstavlja za bakterije in viruse mehansko in kemično prepreko. Če jo patogen uspe prečkati, sproži prirojeni imunski odziv, ki je hiter, a nespecifičen. Sistem prirojene imunosti imajo vse živali in rastline.[11] Najbolj specifičen je sistem pridobljene imunosti, ki ga sproži prirojeni imunski sistem, če se patogenom uspe izogniti prirojenem odzivu. Sistema obrambe aktivirajo receptorji za nespecifične molekulske vzorce (pri prirojenemu imunskemu sistemu) in specifične tarčne molekule (antigene), ki jih na svoji površini izražajo patogeni organizmi.
Tako prirojena in pridobljena imunost temeljita na sposobnost imunskega sistema, da razločuje med telesom lastnimi in tujimi molekulami. Pri imunologiji so lastne molekule tiste komponente organizma, ki jih imunski sistem razločuje od tujih snovi.[12] Iz tega sledi, da so tuje molekule tiste, ki jih imunski sistem prepozna za tuje. Ena vrsta takšnih molekul so antigeni (iz angleškega izraza antibody generators, »tvorci protiteles«), ki so definirani kot snovi, ki po vezavi na specifične imunske receptorje pridobljenega imunskega sistema sprožijo imunski odgovor.[1] Imunski sistem proizvaja ogromno različnih receptorjev te vrste, ki se izražajo na površini posebnih celic, imenovanih limfociti (natančneje na limfocitih B in T). Ko se na enega od teh receptorjev veže antigen, je to signal imunskemu sistemu za boj proti točno temu patogenu. Pride do močnega, a lokaliziranega odziva na mestu, kjer je bil patogen zaznan, hkrati pa se manjši del limfocitov pretvori v t. i. spominske celice. Te potujejo do limfatičnih organov, kjer lahko preživijo več let, in stimulirajo proizvodnjo limfocitov z enakimi imunskimi receptorji, kar ob morebitnem kasnejšem vdoru istega patogena omogoči bistveno hitrejši ter učinkovitejši odziv. Pojav, ki mu pravimo imunski spomin, predstavlja temelj imunizacije z vnašanjem umetno inaktiviranih patogenov v telo – cepljenja.
| Prirojeni imunski sistem | Pridobljeni imunski sistem |
|---|---|
| Nespecifični odziv | Odziv je odvisen od vrste patogena in antigena |
| Izpostavljenost vodi v takojšnji največji odziv | Med izpostavljenjem in največjim odzivom je zamik |
| Celične in humoralne komponenete | Celične in humoralne komponente |
| Brez imunskega spomina | Izpostavljenost vodi do imunskega spomina |
| V neki obliki v vseh oblikah življenja na Zemlji | Samo v vretenčarjih s čeljustjo |
Imunskega sistema ni vedno smiselno razločevati na prirojeno in pridobljeno komponento, saj sta medsebojno marsikje povezani in soodvisni. Sesalčji imunski sistem sestavljajo:
- zunanje ovire (koža, sluznice, želodčna kislina, črevesna flora ipd.)
- limfatični organi (priželjc, vranica, bezgavke in limfne kapilare ter žile)
- imunske celice (makrofagi, granulociti, antigen predstavitvene celice, limfociti)
- sistem komplementa
- protitelesa (imunoglobulini – Ig)
- citokini in kemokini
Prirojeni imunski sistem
[uredi | uredi kodo]Mikroorganizmi in toksini, ki uspešno vstopijo v organizem, se srečajo s celicami in mehanizmi prirojenega imunskega odziva. Le-tega navadno sproži vezava mikroba na receptorje za prepoznavo molekularnih vzorcev PRR (pattern recognition receptors), ki prepoznavajo komponente, prisotne pri širokem spektru mikroorganizmov.[13] Sprožijo ga tudi poškodovane celice in celice pod stresom, katerih (nekatere) signale prepoznajo isti receptorji kot za patogene.[14] Prirojena obramba ni specifična, kar pomeni, da se na patogene odziva nespecifično.[1] Ne zagotavlja dolgotrajne imunosti proti patogenu. V večini organizmov je prirojeni imunski sistem poglavitni sistem zaščite.[11]
Površinske ovire
[uredi | uredi kodo]Organizme pred okužbami ščitijo številne različne zunanje bariere – mehanske, kemične in biološke. Voskasta kutikula na večini rastlinskih listov, zunanji skelet pri členonožcih, lupina in membrane na izvaljenem jajcu in koža so primeri mehanskih ovir, ki predstavljajo prvi sloj obrambe pred okužbami.[15] Ker pa organizmi niso povsem ločeni od svojega okolja, so se v okolici telesnih odprtin (pljuča, črevesje, sečila itn.) razvili tudi drugi sistemi obrambe. Kašljanje in kihanje predstavlja mehansko izmetavanje patogenov in drugih dražljivih snovi iz dihalnega trakta. Solze in seč izplavljajo patogene, medtem ko jih sluz v dihalnem in prebavnem traktu ujame.[16]
Tudi kemične ovire ščitijo pred okužbami. Koža in dihalne poti izločajo protimikrobne peptide, kot so β-defenzini.[17] Encimi, kot so lizocim in fosfolipaza A2 v solzah, slini in materinem mleku, so prav tako protibakterijske učinkovine.[18][19] Vaginalni izločki služijo po menarhi kot kemična ovira, saj postanejo rahlo kisli. Semenska tekočina vsebuje defenzine in cink za ubijanje patogenov.[20][21] V želodcu delujejo želodčna kislina in proteaze kot močna kemična obramba pred zaužitimi patogeni.
V sečilih in prebavnem traktu se nahaja komenzalna bakterijska flora, ki služi kot biološka ovira, saj poteka tekma za hrano in prostor med njimi in patogenimi organizmi, včasih tudi s spreminjanjem pH in količine razpoložljivega železa.[22] Zaradi tega simbiotskega razmerja med komenzali in imunskim sistemom je verjetnost, da bodo patogeni dosegli zadostno številčnost za povzročitev bolezni, zmanjšana. Ker pa večina antibiotikov precej nespecifično cilja na bakterije in hkrati ne učinkuje na glive, lahko peroralni antibiotiki vodijo do glivnih okužb, kot so vaginalne kandidoze (okužba s kvasovko iz rodu kandid).[23] Obstajajo zadostni dokazi, da lahko s probiotiki, kot so čiste kulture rodu Lactobacillus iz nepasteriziranega jogurta, dosežemo povrnitev zdravega ravnovesja v mikrobni populaciji po črevesni okužbi, podobne učinke pa lahko dosežemo tudi pri bakterijskem gastroenteritisu, kronični vnetni črevesni bolezni, okužbah sečil in bolnišničnih okužbah prebavil.[24][25][26]
Vnetje
[uredi | uredi kodo]Vnetje je eden prvih odzivov imunskega sistema na okužbo.[27] Simptomi vnetja so rdečina, otekanje, toplota na otip in bolečine, ki so posledica povečanega krvnega pretoka skozi tkivo. Vnetje spodbudijo eikozanoidi in citokini, ki se sproščajo iz poškodovanih in okuženih celic. Med eikozanoide štejemo med drugim prostaglandine, ki povzročajo dvig telesne temperature in dilatacijo krvnih žil, povezano z vnetjem, in levkotriene, ki pritegnejo nekatere bele krvne celice (levkocite).[28][29] Pogosti citokini so interlevkini, odgovorni za komunikacijo med levkociti, kemokini, ki povzročajo kemotakso (gibanje, ki ga izzove kemični stimulant), in interferoni s protivirusnim delovanjem, kot je zaviranje sinteze beljakovin v gostiteljski celici.[30] Včasih se sproščajo tudi rastni dejavniki in citotoksični faktorji. Citokini in druge molekule privlačijo imunske celice na mesto okužbe in spodbujajo zdravljenje poškodovanega tkiva po odstranitvi patogenov.[31]
Sistem komplementa
[uredi | uredi kodo]Sistem komplementa je biokemijska kaskada reakcij, usmerjenih v napad na površino tuje celice. Takšno ime je dobil, ker komplementira/dopolnjuje druge mehanizme ubijanja patogenov. Vključuje preko 20 beljakovin in je pomembna humoralna komponenta prirojenega imunskega sistema.[32][33] Veliko vrst organizmov poseduje svojevrstne sisteme komplementa, med njimi tudi nesesalci, kot so rastline, ribe in nekateri nevretenčarji.[34]
Pri ljudeh ga sproži vezava komplementa na protitelesa, ki so vezana na mikrobe, ali vezava beljakovin sistema komplementa na ogljikove hidrate na površini mikroba. Takšen signal prepoznave sproži hiter ubijalski odziv.[35] Hitrost odziva je posledica ojačitve, ki se zgodi zaradi sosledne proteolitske aktivacije molekul komplementa, iz katerih nastanejo nove proteaze. Po tem, ko se proteini komplementa sprva vežejo na mikrob, aktivirajo svojo proteazno aktivnost, ki posledično aktivira druge proteaze komplementa itn. To privede do katalitske kaskade, ki ojača prvotni signal kot pri nadzorovani pozitivni povratni zanki,[36] nastanejo pa tudi peptidi, ki privlačijo imunske celice, povečajo prepustnost kapilarne stene in opsonizirajo (obdajo) površino patogena, s čimer ga označijo za uničenje. Takšno oblaganje komplementa lahko neposredno ubije celico zaradi motenja plazemske membrane.[32]
Celične ovire
[uredi | uredi kodo]
Levkociti (bele krvne celice) delujejo kot samostojni enocelični organizmi in predstavljajo drugo vejo prirojenega imunskega sistema.[1] Med levkocite prirojenega imunskega sistema spadajo fagociti (nevtrofilci, makrofagi in dendritične celice), tkivni bazofilci (mastociti), bazofilci, eozinofilci in naravne celice ubijalke. Te celice so sposobne patogene prepoznati in odstraniti, kar dosežejo tako, da večje patogene napadejo preko stika ali pa mikroorganizem obdajo in ga ubijejo.[34] Levkociti prirojenega imunskega sistema so pomembni posredniki informacij pri razvoju limfoidnih organov in aktivaciji pridobljenega imunskega sistema.[37]
Fagocitoza je pomemben del celične prirojene imunosti. Celice, imenovane fagociti, obdajo oz. »pojejo« patogene in druge večje delce. Fagociti navadno krožijo po telesu in iščejo patogene, citokini pa jih lahko privabijo na bolj specifično mesto.[1] Ko je patogen ujet v znotrajcelični vezikel (fagosom), pride do združitve z drugim veziklom, imenovanim lizosom, v fagolizosom. Patogen umre zaradi aktivnih prebavnih encimov ali sproščanja prostih radikalov v fagolizosom.[38][39] Fagocitoza se je razvila kot način za pridobivanje hranil, kasneje v evoluciji pa je dobila tudi obrambno vlogo pred patogeni.[40] Je najverjetneje najstarejša oblika gostiteljeve obrambe, saj se fagociti nahajajo tako v vretenčarjih kot nevretenčarjih.[41]
Nevtrofilci in makrofagi so fagociti, ki krožijo po telesu in iščejo patogene.[42] Nevtrofilce po navadi srečamo v krvnem obtoku in predstavljajo največji delež fagocitov (50–60 % vseh levkocitov).[43] V akutni fazi vnetja (zlasti kot posledica bakterijske okužbe) nevtrofilci v procesu kemotakse migrirajo proti mestu vnetja. Navadno tja prispejo prvi. Makrofagi so vsestranske celice, ki se nahajajo v tkivih in proizvajajo široko paleto snovi, med drugim encime, proteine komplementa in citokinov. Delujejo tudi kot lovilci, ki izločajo izčrpane celice in druge ostanke iz telesa. Poleg tega so tudi antigen prestavitvene celice, ki aktivirajo pridobljen imunski sistem.[44]
Dendritične celice (DC) so fagociti v tkivih, ki so v stiku z zunanjim okoljem – v koži, nosu, pljučih, želodcu in črevesju.[45] Ime so dobile po obliki, ki spominja na dendrite živčnega sistema z mnogimi koničastimi izrastki, a z živčevjem nikakor niso povezane. Služijo kot povezava med telesnimi tkivi ter prirojenim in pridobljenim imunskim sistem, saj predstavljajo antigene T-celicam, ki tvorijo enega ključnih celičnih tipov pridobljenega imunskega sistema.[45]
Tkivni bazofilci (mastociti) se nahajajo v vezivnem tkivu in v mukoznih membranah, kjer posredujejo ob vnetnem odzivu.[46] Najpogosteje jih povezujemo z alergijami in anafilaksijo.[43] Bazofilci in eozinofilci so sorodni nevtrofilcem. Sproščajo kemične mediatorje, ki sodelujejo pri obrambi proti zajedavcem, in igrajo ključno vlogo pri alergijskih reakcijah, kot je astma.[47] Naravne celice ubijalke (NK-celice) so levkociti, ki napadajo in uničujejo tumorske celice ter celice, okužene z virusi.[48]
Naravne celice ubijalke
[uredi | uredi kodo]Naravne celice ubijalke (tudi NK-celice) so limfociti in sestavni del prirojenega imunskega sistema. Mikrobov ne napadajo neposredno.[49] NK-celice uničujejo kompromitirane gostiteljske celice, kot so tumorske celice in celice, okužene z virusi. Prepoznajo jih po stanju, imenovanem »missing self« (»manjkajoče sebstvo«). Ta izraz opisuje celice z nizkim nivojem označevalca PHK I na celičnem površju, kar se lahko zgodi ob virusni okužbi gostiteljeve celice.[34] Ime »naravne ubijalke« so dobile zaradi prvotnega prepričanja, da za ubijanje celic preko principa missing self ne potrebujejo aktivacije. Že mnoga leta ostaja uganka, kako NK-celice prepoznajo tumorske celice in okužene celice. Vemo samo, da jih aktivira pomanjkanje PHK I. Normalne celice v telesu se izognejo napadu NK-celic, saj izražajo nespremenjene lastne antigene PHK, ki jih prepoznajo recceptorji, ki zavirajo napad.[50]
Pridobljeni imunski sistem
[uredi | uredi kodo]Pridobljeni imunski sistem se je razvil v zgodnjih vretenčarjih in omogoča močnejši imunski odziv, pojavi pa se tudi imunski spomin, pri kateremu si imunski sistem "zapomni" značilne antigene posameznega patogenega organizma.[51] Pridobljeni imunski odziv sproži specifičen antigen, kar zahteva prepoznavanje tujih molekul med procesom predstavitve antigena. Specifičnost antigenov omogoča odziv, ki je ukrojen za specifičen patogen ali celico, okuženo s patogenom. To sposobnost priprave odgovora ohranjajo "spominske celice". Če patogen ponovno okuži telo, ga spominske celice hitro odstranijo.
Limfociti
[uredi | uredi kodo]Celice pridobljenega imunskega sistema so posebna vrsta levkocitov, imenovane limfociti. Limfociti B in limfociti T sta veliki podskupini limfocitov, ki izhajata iz matičnih krvotvornih celic kostnega mozga.[34] B-celice so vključene v humoral imunski odziv, T-celice pa v celični imunski odziv.
Tako B-celice kot T-celice prenašajo receptorske molekule, ki prepoznajo specifične tarče. T-celice prepoznajo tuje tarče, kot so patogeni, šele ko so bili antigeni (majhni deli patogena) obdelani in predstavljeni na površini celice skupaj s človeku lastnim receptorjem, imenovanim poglavitni histokompatibilnostni kompleks (PHK, angleško tudi MHC – major histocompatibility complex). Obstajata dva glavna podtipa T-celic: celice ubijalke in celice pomagalke. Poleg njiju poznamo še regulatorne T-celice, ki igrajo vlogo pri uravnavanju imunskega odziva. Celice ubijalke prepoznajo samo antigene, sparjene z molekulami PHK tipa I, pomagalke in regulatorne celice pa antigene, sparjene z molekulami PHK tipa II. Dva različna mehanizma kažeta na različni vlogi celic ubijalk in pomagalk. Tretji in manjši podtip so γδ T-celice, ki prepoznavajo nedotaknjene antigene, ki niso povezani z receptorjem PHK.[52] Dvojno pozitivne T-celice so v priželjcu izpostavljene raznolikim »lastnim« antigenom. Za normalno delovanje priželjca je potrebna zadostna količina joda.[53]
Po drugi strani so za antigen specifični receptorji na površini B-celic molekule protiteles, ki prepoznajo celotne patogene brez potrebe po procesiranju antigenov. Vsaka linija B-celic izraža drugo protitelo, zato predstavlja celota vseh B-celičnih antigenskih receptorjev vsa protitelesa, ki jih telo lahko porizvede.[34]
Celice ubijalke
[uredi | uredi kodo]Celice ubijalke so podskupina T-celic. Ubijajo celice, okužene z virusi (in drugimi patogeni), poškodovane celice in druge nefunkcionalne celice.[54] Kot pri B-celicah tudi posamezni tip T-celic prepoznava specifičen antigen. Celice ubijalke aktivira vezava tega specifičnega antigena v kompleksu z PHK I druge celice na T-celični receptor (TRC). Pri prepoznavanju kompleksa PHK-antigen pomaga tudi koreceptor CD8 na T-celici. Le-ta nato potuje po telesu in išče celice, kjer PHK I nosi ta antigen. Ob stiku s takšno celico sprosti citotoksične snovi, kot so perforini, ki tvorijo pore v plazemsko membrano tarčne celice, kar omogoča vdor ionov, vode in toksinov v celico. Vdor toksina granulizina (proteaza) sproži apoptozo v tarčni celici.[55] Ubijanje celic preko celic ubijalk je posebej pomembno pri preprečevanju razmnoževanja virusov. Aktivacija T-celic se začne šele ob zelo močnem aktivacijskem signalu po nastanku kompleksa PHK-antigen ali ob dodatnih aktivacijskih signalih celic pomagalk.[55]
Celice pomagalke
[uredi | uredi kodo]
Celice pomagalke uravnavajo delovanje prirojenega in pridobljenega imunskega sistema in določajo, kateri odziv na določeni patogen se sproži.[56][57] Same po sebi niso citotoksične (ne ubijajo okuženih celic in ne odstranjujejo patogenov). S tem, ko usmerjajo druge celice, nadzorujejo imunski odziv .
Celice pomagalke izražajo T-celične receptorje (TCR), ki prepoznavajo antigene, vezane na molekule PHK II. Kompleks antigen-PHK prepoznajo tudi koreceptorji CD4, ki novačijo molekule iz T-celice (npr. Lck tirozinsko kinazo), ki so odgovorne za aktivacijo celice. Pomagalke so šibkeje povezane z mehanizmom aktivacije preko kompleksa PHK-antigen kot celice ubijalke, saj je pri slednjih za aktivacijo potrebna zgolj ena povezava kompleksa s T-celičnim receptorjem, pri pomagalkah pa okoli 200–300. Pomagalke potrebujejo tudi daljši stik z antigen predstavitveno celico za aktivacijo.[58] Aktivacija mirujoče celice pomagalke sproži sproščanje citokinov, ki vplivajo na delovanje mnogih tipov celic. Citokinski signali celic pomagalk povečajo mikrobicidno delovanje makrofagov in aktivnost celic ubijalk.[1] Poleg tega sproži aktivacija pomagalk zviševanje ravni molekul, izraženih na površini T-celic, npr. CD40, kar priskrbi dodaten stimulatorni signal za aktivacijo B-celične proizvodnje protiteles.[59]
Gama delta T-celice
[uredi | uredi kodo]Gama delta T-celice (γδ T-celice) imajo alternativni T-celični receptor (TCR) v nasprotju s CD4+ in CD8+ (αβ) T-celicami. Delijo si značilnosti celic pomagalk, citotoksičnih T-celic in naravnih celic ubijalk. Pogoji, ki povzročijo odziv γδ T-celic, niso v celoti raziskani. Kot druge »nekonvencionalne« podskupine T-celic z nespremenljivimi receptorji TCR, kot so na CD1d omejene naravne T-celice ubijalke, γδ T-celice prehajajo mejo med prirojeno in pridobljeno imunostjo.[60] Po eni strani so komponenta pridobljene imunosti, saj rekombinirajo gene za TCR za ustvarjanje raznolikih receptorjev in so zmožne tudi tvorbe spominskih fenotipov, po drugi strani pa lahko sodelujejo tudi pri prirojenem imunskem odzivu, saj izražajo nespremenljive TCR in NK receptorje za prepoznavo manj specifičnih molekularnih vzorcev PRR (na primer Vγ9/Vδ2 T-celice se odzovejo na pogoste mikrobne molekule, močno omejene Vδ1+ T-celice epitelija pa na epitelijske celice pod stresom).[52]

Limfociti B in protitelesa
[uredi | uredi kodo]Limfociti B prepoznajo patogen ob vezavi specifičnega tujega antigena na njihovo površinsko protitelo.[62] Ta kompleks antigen-protitelo nato prevzamejo in proteolitsko obdelajo do peptidov, ki jih nato predstavijo na površinskih molekulah PHK II. Ta kombinacija antigena in PHK privlači ujemajočo se T-celico pomagalko, ki izloča limfokine in B-celico aktivira.[63] Ko se le-ta nato začne deliti, njene potomke (plazmatke) izločajo milijone kopij protitelesa, ki prepoznava isti antigen. Protitelesa krožijo po krvi in limfi, vežejo se na patogene s tem antigenom in jih označijo za uničenje z aktivacijo komplementa ali s privzemom in uničenjem v fagocitu. Protitelesa lahko tudi direktno nevtralizirajo bakterijske toksine ali se vežejo na virusne in bakterijske receptorje, ki jih le-ti uporabljajo za okužbo celice.[64]
Alternativni pridobljeni imunski sistem
[uredi | uredi kodo]Evolucijski razvoj pridobljenega imunskega sistema se je zgodil v predniku vretenčarjev s čeljustjo. Mnoge klasične molekule pridobljenega imunskega sistema (npr. protitelesa in TCR) obstajajo samo v njih. Vseeno pa pri nekaterih primitivnih vretenčarjih brez čeljusti, kot so piškurji in glenavice, obstajajo molekule, ki izvirajo iz limfocitov. Te organizmi vsebujejo široko paleto molekul VLR (variabilni limfocitni receptor), ki izvirajo iz enega ali dveh genov in ki naj bi vezale patogenske antigene podobno kot protitelesa z določeno stopnjo specifičnosti.[65]
Imunski spomin
[uredi | uredi kodo]
Ko se začnejo aktivirani limfociti B in T deliti, postanejo nekateri njihovi potomci dolgožive spominske celice. Tekom življenja osebka si te celice zapomnijo vse specifične patogene, s katerimi so bile v stiku, in lahko sprožijo močan odziv, če pridejo z njimi v ponoven stik. Takšen odziv je »prilagojen«, ker se zgodi tekom posameznikovega življenja kot prilagoditev na okužbo z nekim določenim patogenom in kot priprava imunskega sistema na prihodnje izzive. Imunski spomin delimo na pasivni kratkotrajni in aktivni dolgotrajni spomin.
Pasivni spomin
[uredi | uredi kodo]Novorojenci še niso bili izpostavljeni mikrobom in so posebej občutljivi za okužbe, zato jih mati oskrbuje s pasivno zaščito. Med nosečnostjo se protitelesa družine IgG preko posteljice prenašajo iz matere na otroka, zato imajo novorojenci v krvi visoko koncentracijo protiteles proti istim specifičnim antigenom kot njihove matere.[66] Tudi materino mleko vsebuje protitelesa, ki se prenašajo v dojenčkovo črevesje, kjer ga varujejo pred bakterijskimi okužbami, dokler jih ni sposoben izdelovati sam.[67] Takšni zaščiti pravimo pasivna zaščita, saj spominske celice in protitelesa v fetusu ter dojenčku še ne nastajajo, ampak vanj samo pasivno prehajajo. Kmalu po rojstvu pa prevzame dojenčkov lasten imunski sistem nalogo, ki jo bo opravljal vse življenje: prepoznavanje patogenov, kot so gliste, virusi, bakterije in drugi zajedavci.
V medicini se lahko posameznika pasivno zaščiti z vbrizganjem seruma, bogatega s protitelesi.[68]
Aktivni spomin in cepljenja
[uredi | uredi kodo]Dolgotrajni aktivni spomin se tvori po okužbi, ki aktivira limfocite B in T. Ustvarimo ga lahko tudi umetno s cepljenjem. Princip v ozadju cepljenja (imenovanim tudi imunizacija) je predstavitev antigena iz patogena za stimulacijo imunskega sistema in razvoja specifične imunosti proti tem določenem patogenu brez povzročitve bolezni, povezane s tem organizmom.[1] Takšna namerna povzročitev imunskega odgovora je uspešna, ker izrablja specifičnost imunskega sistema. Ker ostajajo pri človeku nalezljive bolezni eden od glavnih razlogov smrti, predstavlja cepljenje najbolj učinkovito manipulacijo imunskega sistema, ki jo je človeštvo razvilo.[34][69]
Večina virusnih celiv temeplji na oslabljenih (atenuiranih) virusih, bakterijska pa vsebujejo necelične komponente mikroorganizmov (npr. neškodljive komponente toksina).[1] Ker mnogi antigeni v neceličnih cepivih ne sprožijo močnega pridobljenega odgovora, je treba v bakterijska cepiva dodati adjuvanse, ki aktivirajo antigen predstavitvene celice prirojenega imunskega sistema in povečajo imunogenost.[70]
Okvare
[uredi | uredi kodo]Imunski sistem je izredno učinkovita struktura, ki vključuje specifičnost, sprožanje po stimulu (inducibilnost) in prilagajanje. Motnje v delovanju imunskega sistema se lahko odražajo v šibkejšem, močnejšem ali manj specifičnem delovanju. Imunsko pomanjkljivost pri človeku lahko povzročijo genetske okvare, pridobljena stanja, kot je HIV/AIDS, ali uporaba imunosupresivov, posledica česar je manjša odpornost proti nalezljivim boleznim. Nasprotno motnje v regulaciji imunskega odziva povzročajo vnetne bolezni. Poleg tega se lahko imunski sistem okvari tako, da prepoznava telesu lastne molekule kot tuje, zaradi česar napada lastna tkiva - posledica so avtoimunske bolezni, npr. revmatoidni artritis, lupus, idr. (odvisno od prizadetega tkiva). Stanja nezmožnosti gostiteljevega imunskega sistema, da brani, lahko razdelimo v tri kategorije: imunska pomanjkljivost, avtoimunske bolezni in preobčutljivostne reakcije.
Imunska pomanjkljivost
[uredi | uredi kodo]Do imunskih pomanjkljivosti pride, ko je ena ali več komponent imunskega sistema neaktivna. Sposobnost imunskega sistema, da se odzove na patogene, je lahko oslabljena pri starejših in mlajših, pri čemer začne pešati okoli 50 leta zaradi imunosenescence.[71][72] V razvitih državah so pogosti razlogi za oslabljen imunski sistem debelost, zloraba alkohola in drog. V državah v razvoju je glavni razlog nedohranjenost. Prehrana brez zadostnih beljakovin lahko vodi v oslabljeno celično imunost, zmanjšano aktivnost sistema komplementa, funkcijo fagocitov, koncentracije protiteles razreda IgA in proizvodnje citokinov. Izguba priželjca v zgodnjih letih življenja zaradi genskih mutacij ali kirurške odstranitve privede do hude imunske pomanjkljivosti in visoke dovzetnosti za okužbe.[73]
Imunske pomanjkljivosti so lahko dedne ali »pridobljene«. Kronična granulomatozna bolezen, kjer je zmanjšana sposobnost fagocitov, da uničijo patogene, je primer dedne imunske pomanjkljivosti. AIDS in nekateri tipi raka povzročajo pridobljeno pomanjkljivost.[74][75]
Avtoimunske bolezni
[uredi | uredi kodo]Drugo stran motenj delovanja imunskega sistema predstavljajo avtoimunske okvare. V tem primeru je imunski sistem preveč aktiven in ne loči med telesu lastnimi in tujimi molekulami, zato napada dele telesa. T-celice in protitelesa tudi pod normalnimi pogoji reagirajo na telesu lastne peptide.[76] Ena izmed funkcij specializiranih celic, ki se nahajajo v priželjcu in kostnem mozgu, je predstavljanje lastnih antigenov iz vsega telesa mladim limfocitom in izločevanje tistih celic, ki prepoznavajo telesu lastne antigene, s čimer v normalnih okoliščinah preprečujejo avtoimunost.
Preobčutljivostne reakcije
[uredi | uredi kodo]Preobčutljivostne reakcije so imunski odzivi, ki poškodujejo telesu lastno tkivo. Delimo jih na štiri tipe (tip I – IV) glede na vključene mehanizme in časovni potek reakcije. Preobčutljivost tipa I je takojšnja oz. anafilaktična reakcija, pogosto povezana z alergijami. Simptomi segajo od blagega nelagodja, vse do hujših zapletov, ki vodijo do smrti. Mediatorji tega tipa preobčutljivosti so protitelesa tipa IgE, ki sprožijo degranulacijo bazofilcev in mastocitov, ko se povežejo z antigeni.[77] Do preobčutljivosti tipa II pride, ko se protitelesa vežejo na antigene osebkovih lastnih celic in jih označijo za uničenje. To imenujemo tudi od protiteles odvisna (tudi citotoksična) preobčutljivost. Mediirajo jo protitelesa razred IgG in IgM. Imunski kompleksi (agregati antigenov, proteini sistema komplementa ter protitelesa IgM in IgG), naloženi v razna tkiva, sprožijo preobčutljivostne reakcije tipa III. Preobčutljivost tipa IV (celično posredovana oz. »zakasnjena« probčutljivost) se razvija od dva do tri dni. Reakcije tipa IV so del mnogih avtoimunskih in nalezljivih bolezni, vključujejo pa lahko tudi alergijski kontaktni dermatitis. Te reakcije mediirajo T-celice, monociti in makrofagi.
Idiopatsko vnetje
[uredi | uredi kodo]Vnetje je eden prvih odzivov imunskega sistema na okužbo, pojavi pa se lahko tudi brez zunanjega vzroka.
Vnetje povzročijo eikozanoidi in citokini, ki jih izločajo poškodovane in okužene celice. Družino eikozanoidov sestavljajo med drugim prostaglandini, ki povzročajo vročino, dilatacijo krvnih žil (povezano z vnetjem) in levkotrieni, ki privlačijo določene bele krvničke (levkocite). Med pogoste citokine spadajo interlevkini, ki so odgovorni za komunikacijo med belimi krvničkami, kemokini, ki spodbujajo kemotakso, in interferoni s protivirusnim delovanjem, kot je zaustavitev sinteze proteinov v gostiteljskih celicah. Pride tudi do sproščanja rastnih in citotoksičnih dejavnikov. Citokini in druge snovi spodbujajo delovanje imunskih celic na mestu okužbe in tudi celjenje poškodovanega tkiva po odstranitvi patogenov.
Drugi mehanizmi in evolucija
[uredi | uredi kodo]Zelo verjetno je, da so vretenčarji prvi razvili večkomponenten, prilagodljiv imunski sistem, saj nevretenčarji ne tvorijo limfocitov ali humoralnega odziva na osnovi protiteles. Veliko vrst organizmov pa vseeno uporablja mehanizme, ki delujejo kot predhodniki teh vidikov vretenčarske imunosti. Imunski sistem se pojavi celo v strukturno najpreprostejših oblikah življenja. Bakterije uporabljajo encime in ti. restrikcijsko modifikacijske sisteme za zaščito pred virusnimi patogeni – bakteriofagi.[78] Prokarionti posedujejo tudi pridobljeno imunost. Preko sistema, ki uporablja sekvence CRISPR za zadržanje delov fagovega genoma, s katerim so prišli v preteklosti že v stik, lahko kasneje z obliko interference RNA zavirajo virusno podvojevanje.[77][79] Prokarionti imajo tudi druge mehanizme.[80][81] Napadalni elementi imunskega sistema so prisotni tudi pri enoceličnih evkariontih, a je študij o le-teh malo.[82]
Vzorčno prepoznavni receptorji so proteini, ki jih uporabljajo skoraj vsi organizmi za prepoznavanje molekul, povezanih s patogeni. Protimikrobni peptidi defenzini so preko evolucije ohranjene komponente prirojenega imunskega sistema v vseh rastlinah in živalih. Predstavljajo glavno obliko nevretenčarske sistemske imunosti. Večina nevretenčarjev uporablja tudi sistem komplementa in fagocitne celice. Interferenca RNA in ribonukleaze so verjetno obstajale v neki obliki pri predniku vseh evkariontov in se ohranile pri mnogih. Igrale naj bi vlogo pri imunskem odgovoru na virusno okužbo.[83][84] Imunski sistem sesalcev je nadgradnja teh sistemov, ki omogoča pridobljeno imunost oz. večjo učinkovitost proti že znanim patogenom, a sistemi prirojene imunosti tudi pri njih ohranjajo pomembno vlogo. Skupaj sestavljajo kompleksen in še ne popolnoma razumljen splet molekul in celic, udeleženih pri imunskem odzivu.
Za razliko od živali rastline nimajo fagocitnih celic. Mnoge rastline se odzovejo z lokalnim in sistemskim kemičnim signaliziranjem, ki potuje po rastlini.[85] Posamezne rastlinske celice se odzovejo na molekule, povezane s patogeni – PAMP (angleško Pathogen-Associated Molecular Patterns).[86] Ko je del rastline okužen, začne rastlina ustvarjati lokalizirano preobčutljivostno reakcijo, pri kateri je v okoliških celicah hitro sprožena apoptoza za preprečitev širjenja bolezni v druge dele rastline.[87] Sistemska pridobljena odpornost je vrsta obrambnega odziva v rastlini, ki nudi odpornost proti določenem agensu za celo rastlino. Interferenca RNA je zelo pomembna v sistemskem odzivu, saj lahko blokira virusno replikacijo.[88]
Imunologija tumorjev
[uredi | uredi kodo]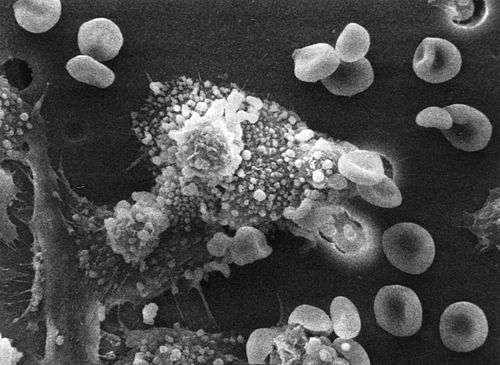
Druga pomembna vloga imunskega sistema je prepoznavanje in odpravljanje tumorjev. Imenuje se tudi imunski nadzor. Tumorsko spremenjene celice izražajo drugačne antigene na svoji površini kot zdrave celice. Imunskem sistemu delujejo takšni antigeni tuji in njihova prisotnost sproži odziv za odstranitev celic. Izražani antigeni izvirajo iz različnih virov;[90] nekateri so del onkogenih virusov, kot je HPV, ki povzroča raka materničnega vratu[91], drugi pa so organizmu lastni proteini, ki se na normalnih celicah pojavljajo v nizkih koncentracijah, na tumorskih celicah pa v visokih. Primer tega je encim tirozinaza, ki pri višjih koncentracijah pretvarja določene kožne celice (na primer melanocite) v tumorje – melanome.[92][93] Tretji možni vir tumorskih antigenov so proteini, ki navadno prisotni pri regulaciji celične rasti in preživetja, ki pogosto mutirajo v molekule, ki inducirajo raka – onkogene.[94][95]
Glavni odziv imunskega sistema proti tumorjem je uničevanje nenormalnih celic s T-celicami ubijalkami, včasih s pomočjo T-celic pomagalk.[96] Tumorski antigeni so predstavljeni na molekulah PHK I podobno kot virusni antigeni. To omogoča T-celično prepoznavo tumorskih celic kot nenormalnih.[97] Tudi naravne celice ubijalke ubijajo na podoben način, sploh če je molekul PHK I na površju celic manj kot navadno.[98] Včasih nastanejo proti tumorjem tudi protitelesa, kar omogoča uničenje preko sistema komplementa.
Nekateri tumorji se izognejo imunskem sistemu in postanejo rakava obolenja.[99][100] Tumorske celice imajo pogosto manj molekul PHK I na površini, s čimer se izognejo nadzoru celic ubijalk. Nekateri tumorji tudi izločajo produkte, ki inhibirajo imunski odziv, npr. citokin TGF-β, ki zavira aktivnost makrofagov in limfocitov.[101] Razvije se lahko tudi imunska toleranca, ko imunski sistem preneha napadati tumorske celice.
Makrofagi lahko paradoksalno promovirajo tumorsko rast,[102] ko tumorske celice izločajo citokine za privlak makrofagov, ki potem tvorijo citokine in rastne dejavnike kot je TNF-α, ki spodbuja razvoj tumorjev in promovira plastičnost teh celic kot pri matičnih celicah. Kombinacija hipoksije v tumorju in citokin iz makrofagov lahko dodatno pripomore k širjenju rakavih celic, saj povzroči zmanjšano proizvodnjo proteina, ki bi blokiral metastazo teh celic.
Fiziološka regulacija
[uredi | uredi kodo]Imunski sistem je vključen v mnoge vidike fiziološke regulacije v telesu. Je tesno prepleten z drugimi sistemi, kot sta endokrini[103][104] in živčni sistem.[105][106][107] Imunski sistem igra tudi poglavitno vlogo pri razvoju zarodka ter popravljanju in regeneraciji tkiva.
Hormoni
[uredi | uredi kodo]Hormoni lahko deluje kot imunomodulatorji, saj spreminjajo občutljivost imunskega sistema. Ženski spolni hormoni so primer imunostimulatorjev tako pridobljenih[108] kot prirojenih imunskih odgovorov.[109] Nekatere bolezni, kot je lupus, pogosteje prizadenejo ženske s pričetkom sprememb v puberteti.[110] Po drugi strani pa so moški spolni hormoni, kot je testosteron, imunosupresivni. Tudi drugi hormoni delujejo na imunski sistem, predvsem prolaktin, rastni hormon in vitamin D.[111][112]
Vitamin D
[uredi | uredi kodo]Ko celica T sreča tuji patogen, iztegne receptor za vitamin D. Ta deluje kot signalna naprava, ki omogoča vezavo celic T z aktivno obliko vitamina D, steroidnim hormonom kalcitriolom. Celice T imajo simbiotsko razmerje z vitaminom D, ne samo zaradi tega receptorja, ampak tudi ker izražajo gen CYP27B1, ki je odgovoren za presnavljanje predhodne oblike vitamina D – kalcidiola v aktivno obliko kalcitriol. Šele po tej vezavi lahko celice T opravljajo svoje naloge. Druge imunske celice, ki izražajo gen CYP27B1, so dendritične celice, keratinociti in makrofagi.[113][114]
Domnevajo, da je nižanje koncentracije hormonov s starostjo delno odgovorno za oslabljen imunski odziv v starajočem se telesu.[115] Nasprotno je pri nekaterih hormonih, katerih koncentracijo uravnava prav imunski sistem (predvsem pri ščitničnih hormonih).[116] Slabljenje imunske funkcije, povezano s starostjo, je povezano z zmanjšanjem koncentracije vitamina D pri starostnikih. Dogajata se dve stvari, ki negativno vplivata na količino vitamina: starostniki se manj gibajo na prostem (tu je pomembno sevanje UVB), zaradi česar proizvedejo tudi manj holekalciferola, starostniška koža pa tudi sama po sebi izgublja sposobnost proizvodnje vitamina D.[117]
Spanje in počitek
[uredi | uredi kodo]Spanje in počitek vplivata na imunski sistem.[118] Pomanjkanje spanja je lahko pogubno za imunske funkcije.[119] Kompleksna povratna zanka, ki vključuje citokine, kot sta interlevkin-1 in dejavnik tumorske nekroze α, udeležene pri imunskem odgovoru na okužbo, vpliva tudi na regulacijo spanja REM in NREM.[120] Imunski odgovor na okužbo lahko tako vpliva na spalni cikel, predvsem se poveča razmerje med spanjem počasnih valov in spanjem REM.[121]
Pri ljudeh s pomanjkanjem spanja ima aktivna imunizacija manjši učinek, zmanjšana je proizvodnja protiteles v primerjavi s spočitim posameznikom. Poleg tega lahko neprespanost in delo v izmenah zaradi motenj v izpostavljenosti naravni svetlobi in temi vpliva na proteine, kot je npr. NFIL3, ki je povezan z diferenciacijo celic T in cirkadianim ritmom. Takšne motnje lahko vodijo v kronična obolenja, kot so bolezni srca, kronično bolečino in astmo.[122]
Spanje in z njim povezani cirkadiani ritem močno uravnavajo imunološke funkcije teko prirojenega kot pridobljenega imunskega sistema. V zgodnji fazi spanja počasnih valov pride do padca krvnih koncentracij kortizola, adrenalina in noradrenalina, ki povzroči povišanje koncentracij hormonov leptina, rastnega hormona iz hipofize in prolaktina. Te signali vzpodbujajo pro-vnetno okolje preko proizvodnje citokinov IL-1, IL-12, TNF-α in INFγ. Te citokini spodbujajo aktivacijo, proliferacijo in diferenciacijo imunskih celic. V tem času dosežejo nediferenciirane in manj diferenciirane imunske celice (naivne in centralne spominske celice T) svoj vrhunec – v času počasi razvijajočega se pridobljenega imunskega odziva. Poleg teh učinkov hormoni leptin, rastni hormon in prolaktin podpirajo interakcijo med antigen predstavljajočimi celicami in celicami T, premik ravnovesja med pomagalkami Th1/Th2 na stran Th1, proliferacijo pomagalk Th, migracijo naivnih celic T v bezgavke, verjetno pa podpirajo tudi tvorbo dolgotrajnega imunskega spomina preko sprožitve odziva celic Th1.[123]
Po drugi strani dosežejo v času budnosti vrhunec diferenciirane efektorske celice, kot so citotoksične naravne celice ubijalke in citotoksični limfociti T (CTL). Zaradi tega lahko učinkovito odgovorijo na vdor patogena. Pride tudi do povečane koncentracije protivnetnih molekul, kot so kortizol in kateholamini. Obstajata dve teoriji, zakaj pride do pro-vnetnega stanja med spanjem. Kot prvo bi vnetje med budnostjo vodilo v resno oslabelost fizičnih in miselnih sposobnosti. Poleg tega pa je med spanjem prisotnega več melatonina, ki lajša oksidativen stres (deluje proti tvorbi prostih radikalov) zaradi prisotnosti vnetja.[123][124]
Prehrana
[uredi | uredi kodo]Prevelik vnos hranil lahko vodi do sladkorne bolezni in debelosti, ki vplivata na imunske funkcije. Zmerna podhranjenost, pomanjkanje določenih mineralov in hranil lahko oslabijo imunski odziv.[125]
Živila, bogata z nekaterimi maščobnimi kislinami, lahko spodbujajo zdrav imunski sistem.[126] Podhranjenost zarodka lahko vodi v vseživljenjsko oslabelost imunskega sistema.[127]
Popravila in regeneracije
[uredi | uredi kodo]Imunski sistem, predvsem prirojeni del, igra ključno vlogo v popravljanju tkiva po poškodbi.[128][129][130][131][132] Ključni akterji so makrofagi in nevtrofilci, druge celične komponente, vključno z γδ celicami T, prirojene limfoidne celice (ILC) in regulatorne celice T (Treg), pa so prav tako pomembne. Plastičnost imunskih celic in ravnovesje med pro-vnetnimi ter protivnetnimi signali so ključni vidiki učinkovitega popravila tkiva.[132] Imunske komponente in poti so vključene v regeneracijo tkiv tudi pri npr. dvoživkah. Glede na eno hipotezo so organizmi, ki se lahko regenerirajo, morda manj imunsko kompetentni kot organizmi, ki se ne morejo.[133][134]
Manipulacija v medicini
[uredi | uredi kodo]
Imunski sistem lahko manipuliramo v smeri zatiranja neželenih odzivov (zaradi npr. avtoimunosti, alergij in zavrnjenih vsadkov) ali v smeri spodbujanja varovalnih odzivov proti patogenom, ki se izogibajo imunskem odzivu, ali raku.
Imunosupresija
[uredi | uredi kodo]Imunosupresivi so razred zdravil, s katerimi uravnavamo potek avtoimunskih bolezni ali vnetij, ko pride do prevelikih poškodb tkiva, lahko pa tudi preprečimo zavrnitev vsadka po presaditvi organa.[135]
Protivnetna zdravila uporabljamo za nadzorovanje učinkov vnetja. Glukokortikoidi so najmočnejša izmed teh zdravil, a imajo mnogo neželenih stranskih učinkov, kot so centralna debelost, hiperglikemija in osteoporoza, zato jih je potrebno ustrezno regulirati.[136] Nižje odmerke protivnetnih zdravil se pogosto uporablja v kombinaciji s citotoksičnimi ali imunosupresivnimi zdravili, kot sta metotreksat in azatioprin. Citotoksična zdravila zavirajo imunski odziv z ubijanjem celic (npr. aktiviranih celic T) med delitvijo. To ubijanje pa ne loči med celicami, zato odmrejo vse celice, ki se stalno delijo, kar privede do toksičnih stranskih učinkov.[135] Imunosupresivna zdravila, kot so ciklosporini, preprečujejo celicam T, da bi se pravilno odzvale na signale s tem, ko inhibirajo poti prenosa signalov.[137]
Imunostimulacija
[uredi | uredi kodo]Imunoterapija raka pokriva vse načine za stimulacijo imunskega sistema v smeri napada na rakave celice.
Teoretični pristopi k imunskemu sistemu
[uredi | uredi kodo]Imunologija je zaradi vsakodnevne prakse zelo eksperimentalna veda, a se je vseeno že od nekdaj lotevamo tudi teoretično. Predlagane so bile že mnoge teorije. Na koncu 19. in v začetku 20. stoletja je potekal boj med »celično« in »humoralno« teorijo imunosti. Po prvi Ilje Iljiča Mečnikova so za odziv odgovorne celice, predvsem fagociti, po drugi Roberta Kocha in Emila von Behringa pa so aktivna komponenta topne molekulske komponente, prisotne v telesnem »humorusu« – tekočinah.[138][139][140]
Sredi 50. let dvajsetega stoletja je Frank Burnet po navdihu Nielsa Jerneja[141] oblikoval teorijo klonske selekcije (CST, Clone Selection Theory).[142] Glede na CST se imunski odziv tvori zaradi razlikovanja med telesu lastnimi in tujimi snovmi: lastne komponente (sestavine telesa) ne sprožijo uničevalnega imunskega odziva, medtem ko tuje komponente (patogeni, vsadki drugega organizma iste vrste) sprožijo uničevalen imunski odziv.[143] Teorijo so kasneje prilagodili novoodkritem konceptu tkivne skladnosti oz. kompleksne »dvosignalne« aktivacije celic T.[144] Delitev »lastno—tuje« je deležna mnogih kritik,[145][146] a ostaja zelo vplivna.[147][148]
Danes so bolj priznani novi teoretski okvirji, med njimi »avtopoetsko« stališče,[149] stališče »miselne imunosti«[150], model nevarnosti (»Danger Model/Theory«) in teorija diskontinuitete.[151][152][153] Model nevarnosti, ki ga je predlagala Polly Matzinger s kolegi, je bil zelo vpliven, saj je odprl veliko razprav in izzval veliko komentarjev.[154][155][156][157]
Napovedovanje imunogenosti
[uredi | uredi kodo]Večja zdravila (molekule nad maso 500 Da) lahko izzovejo imunski odziv nevtralizacije, predvsem če jih moramo vnašati večkrat in v večjih odmerkih. To zmanjšuje učinkovitost zdravil, ki temeljijo na večjih peptidih in beljakovinah (ki so po navadi večje od 6.000 Da). V nekaterih primerih zdravilna učinkovina sama po sebi ni imunogena, a se daje skupaj z imunogeno komponento, kar včasih velja pri Paklitakselu. Razvili so računske metode, s katerimi lahko predvidimo imunogenost pepetidov in beljakovin, zato so zelo uporabne pri snovanju terapevtskih protiteles, vrednotenju verjetne virulence mutacij delcev virusnega ovoja in validaciji predlaganih terapij, ki temeljijo na peptidih. Zgodnje tehnike so se opirale zgolj na predpostavki, da so hidrofilne aminokisline prekomerno zastopane v regijah epitopa v primerjavi s hidrofobnimi;[158] nedavnejši razvoj temelji na tehnikah strojnega učenja s podatkovnimi bazami znanih epitopov, običajno z uporabo dobro raziskanih virusnih beljakovin za učenje algoritma.[159] Ustanovili so tudi javno bazo podatkov za katalogiziranje epitopov, za katere vedo, da jih celice B lahko prepoznavajo.[160] Nastajajoče področje študij imunogenosti, ki temeljijo na bioinformatiki, se imenuje tudi imunoinformatika.[161] Imunoproteomiks je raziskovanje velikih skupin proteinov (proteomika), ki so vključeni v imunski odziv.
Patogenska manipulacija
[uredi | uredi kodo]Uspešnost patogena temelji na njegovi sposobnosti izogibanju gostiteljevemu imunskemu sistemu. Patogeni so zato razvili različne metode, s katerimi uspešno okužijo gostitelja, a se izogibajo odkritju in uničenju s strani njegovega imunskega sistema.[162] Bakterije pogosto premagujejo fizične bariere z izločanjem encimov, ki presnavljajo te ovire (sekrecijski sistem tipa II).[163] Druga možnost je, da uporabljajo sekrecijski sistem tipa III, preko katerega vstavijo v gostiteljevo celico votlo cevko, preko nje pa spuščajo v celico beljakovine, ki onesposobijo gostiteljevo obrambo.[164]
Pogosta tehnika izogibanja prirojenem odzivu je skrivanje znotraj gostiteljevih celic (znotrajcelična patogeneza). Pri tem preživi patogen večji del življenjskega cikla v celici, kjer je obvarovan pred stikom z imunskimi celicami, protitelesi in komplementom. Primeri takšnih organizmov so virusi, bakterije Salmonella (povezane z zastrupitvami s hrano) in evkariontski zajedavci, ki povzročajo malarijo (Plasmodium falciparum) in kala-azar (Leishmania spp.). Druge bakterije, kot so Mycobacterium tuberculosis, živijo v zaščitni kapsuli, ki preprečuje lizo sistema komplementa.[165] Številni patogeni izločajo spojine, ki zmanjšujejo ali napačno usmerijo gostiteljev imunski odziv. Nekatere bakterije (uspešne trajne okužbe – kronična Pseudomonas aeruginosa in Burkholderia cenocepacia, značilne za cistično fibrozo) tvorijo biofilm, da bi se zaščitile pred celicami in beljakovinami imunskega sistema.[166] Druge bakterije ustvarjajo površinske proteine, ki se vežejo na protitelesa, zaradi česar ta postanejo neučinkoivta; primeri so Streptococcus (protein G), Staphylococcus aureus (protein A) in Peptostreptococcus magnus (protein L).[167]
Mehanizmi, s katerimi se izogibajo pridobljenem imunskem sistemu so bolj zapleteni. Najpreprostejši je hitro menjanje neesencialnih epitopov (aminokislin in/ali sladkorjev) na površini patogena, pri čemer ostanejo esencialni zakriti. Dobimo lahko množico organizmov iste vrste (z različnimi izraženimi epitopi), ki jih imenujemo tudi antigenske variacije. Primer tega je virus HIV, ki hitro mutira, da se proteini, ki jih potrebuje za vdor v gostiteljsko celico, stalno menjajo. To spreminjanje pojasni, zakaj cepiva proti HIV ne delujejo.[168] Zajedavec Trypanosoma brucei uporablja podobno strategijo, saj nenehno preklaplja med različnimi vrstami površinskih beljakovin, kar mu omogoča, da ostane vedno korak pred odzivom protiteles.[169] Maskiranje antigenov z gostiteljskimi molekulami je še ena pogosta strategija za izogibanje odkritju. Pri HIV je virusna ovojnica del gostiteljeve zunanje membrane, zaradi česar ga imunski sistem ne prepozna kot tujek.[170]
Sklici in opombe
[uredi | uredi kodo]- ↑ 1,0 1,1 1,2 1,3 1,4 1,5 1,6 1,7 Alberts, Bruce; Johnson, Alexander; Lewis, Julian; Raff, Martin; Roberts, Keith; Walters, Peter (2002). Molecular Biology of the Cell (4. izd.). New York: Garland Science. ISBN 0-8153-3218-1.
- ↑ »Imunost«. Slovenski medicinski e-slovar. Univerza v Ljubljani, Medicinska fakulteta. Pridobljeno 2. aprila 2023.
- ↑ Retief, F.P.; Cilliers, L. (1998). »The epidemic of Athens, 430–426 BC«. South African Medical Journal. Zv. 88, št. 1. str. 50–53. PMID 9539938.
- ↑ Ostoya, P. (1954). »Maupertuis et la biologie«. Revue d'histoire des sciences et de leurs applications. Zv. 7, št. 1. str. 60–78. doi:10.3406/rhs.1954.3379.
- ↑ Plotkin, S.A. (2005). »Vaccines: past, present and future«. Nature Medicine. Zv. 11, št. 4 Suppl. str. S5–11. doi:10.1038/nm1209. PMID 15812490.
- ↑ »The Nobel Prize in Physiology or Medicine 1905«. Nobelprize.org. Nobelov sklad. Pridobljeno 26. aprila 2013.
- ↑ »Major Walter Reed, Medical Corps, U.S. Army«. Walter Reed Army Medical Center. Arhivirano iz spletišča dne 23. oktobra 2007. Pridobljeno 26. aprila 2013.
- ↑ Mečnikov, Ilja Iljič (1905). Immunity in Infective Diseases. Prevod: Binnie, F.G. Cambridge University Press. OCLC 21529.
- ↑ »The Nobel Prize in Physiology or Medicine 1908«. Nobelprize.org. Nobelov sklad. Pridobljeno 26. aprila 2013.
- ↑ »Nobel Prizes and the Immune System«. Nobelprize.org. Nobelov sklad. Pridobljeno 4. junija 2013.
- ↑ 11,0 11,1 Litman, G.W.; Cannon, J.P.; Dishaw, L.J. (2005). »Reconstructing immune phylogeny: new perspectives«. Nature Reviews. Immunology. Zv. 5, št. 11. str. 866–79. doi:10.1038/nri1712. PMID 16261174.
- ↑ Smith A.D. (Ed) Oxford dictionary of biochemistry and molecular biology. (1997) Oxford University Press. ISBN 0-19-854768-4
- ↑ Medzhitov R (Oktober 2007). »Recognition of microorganisms and activation of the immune response«. Nature. 449 (7164): 819–26. Bibcode:2007Natur.449..819M. doi:10.1038/nature06246. PMID 17943118.
- ↑ Matzinger P (april 2002). »The danger model: a renewed sense of self« (PDF). Science. 296 (5566): 301–5. Bibcode:2002Sci...296..301M. doi:10.1126/science.1071059. PMID 11951032.
{{navedi časopis}}: Vzdrževanje CS1: samodejni prevod datuma (povezava) - ↑ Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walters P (2002). Molecular Biology of the Cell (Fourth izd.). New York and London: Garland Science. ISBN 978-0-8153-3218-3.
- ↑ Boyton RJ; Openshaw PJ (2002). »Pulmonary defences to acute respiratory infection«. British Medical Bulletin. 61 (1): 1–12. doi:10.1093/bmb/61.1.1. PMID 11997295.
- ↑ Agerberth B; Gudmundsson GH (2006). »Host antimicrobial defence peptides in human disease«. Current Topics in Microbiology and Immunology. Current Topics in Microbiology and Immunology. 306: 67–90. doi:10.1007/3-540-29916-5_3. ISBN 978-3-540-29915-8. PMID 16909918.
- ↑ Moreau JM; Girgis DO; Hume EB; Dajcs JJ; Austin MS; O'Callaghan RJ (september 2001). »Phospholipase A(2) in rabbit tears: a host defense against Staphylococcus aureus«. Investigative Ophthalmology & Visual Science. 42 (10): 2347–54. PMID 11527949.
{{navedi časopis}}: Vzdrževanje CS1: samodejni prevod datuma (povezava) - ↑ Hankiewicz J; Swierczek E (december 1974). »Lysozyme in human body fluids«. Clinica Chimica Acta; International Journal of Clinical Chemistry. 57 (3): 205–9. doi:10.1016/0009-8981(74)90398-2. PMID 4434640.
{{navedi časopis}}: Vzdrževanje CS1: samodejni prevod datuma (povezava) - ↑ Fair WR; Couch J; Wehner N (Februar 1976). »Prostatic antibacterial factor. Identity and significance«. Urology. 7 (2): 169–77. doi:10.1016/0090-4295(76)90305-8. PMID 54972.
- ↑ Yenugu S; Hamil KG; Birse CE; Ruben SM; French FS; Hall SH (Junij 2003). »Antibacterial properties of the sperm-binding proteins and peptides of human epididymis 2 (HE2) family; salt sensitivity, structural dependence and their interaction with outer and cytoplasmic membranes of Escherichia coli«. The Biochemical Journal. 372 (Pt 2): 473–83. doi:10.1042/BJ20030225. PMC 1223422. PMID 12628001.
- ↑ Gorbach SL (Februar 1990). »Lactic acid bacteria and human health«. Annals of Medicine. 22 (1): 37–41. doi:10.3109/07853899009147239. PMID 2109988.
- ↑ Hill LV; Embil JA (Februar 1986). »Vaginitis: current microbiologic and clinical concepts«. CMAJ. 134 (4): 321–31. PMC 1490817. PMID 3510698.
- ↑ Reid G; Bruce AW (Avgust 2003). »Urogenital infections in women: can probiotics help?«. Postgraduate Medical Journal. 79 (934): 428–32. doi:10.1136/pmj.79.934.428. PMC 1742800. PMID 12954951.
- ↑ Salminen SJ; Gueimonde M; Isolauri E (Maj 2005). »Probiotics that modify disease risk«. The Journal of Nutrition. 135 (5): 1294–8. PMID 15867327.
- ↑ Reid G; Jass J; Sebulsky MT; McCormick JK (Oktober 2003). »Potential uses of probiotics in clinical practice«. Clinical Microbiology Reviews. 16 (4): 658–72. doi:10.1128/CMR.16.4.658-672.2003. PMC 207122. PMID 14557292.
- ↑ Kawai T; Akira S (Februar 2006). »Innate immune recognition of viral infection«. Nature Immunology. 7 (2): 131–7. doi:10.1038/ni1303. PMID 16424890.
- ↑ Miller SB (Avgust 2006). »Prostaglandins in health and disease: an overview«. Seminars in Arthritis and Rheumatism. 36 (1): 37–49. doi:10.1016/j.semarthrit.2006.03.005. PMID 16887467.
- ↑ Ogawa Y; Calhoun WJ (Oktober 2006). »The role of leukotrienes in airway inflammation«. The Journal of Allergy and Clinical Immunology. 118 (4): 789–98, quiz 799–800. doi:10.1016/j.jaci.2006.08.009. PMID 17030228.
- ↑ Le Y; Zhou Y; Iribarren P; Wang J (april 2004). »Chemokines and chemokine receptors: their manifold roles in homeostasis and disease« (PDF). Cellular & Molecular Immunology. 1 (2): 95–104. PMID 16212895.
{{navedi časopis}}: Vzdrževanje CS1: samodejni prevod datuma (povezava) - ↑ Martin P; Leibovich SJ (november 2005). »Inflammatory cells during wound repair: the good, the bad and the ugly«. Trends in Cell Biology. 15 (11): 599–607. doi:10.1016/j.tcb.2005.09.002. PMID 16202600.
{{navedi časopis}}: Vzdrževanje CS1: samodejni prevod datuma (povezava) - ↑ 32,0 32,1 Rus H; Cudrici C; Niculescu F (2005). »The role of the complement system in innate immunity«. Immunologic Research. 33 (2): 103–12. doi:10.1385/IR:33:2:103. PMID 16234578.
- ↑ Degn SE; Thiel S (Avgust 2013). »Humoral pattern recognition and the complement system«. Scandinavian Journal of Immunology. 78 (2): 181–93. doi:10.1111/sji.12070. PMID 23672641.
- ↑ 34,0 34,1 34,2 34,3 34,4 34,5 Janeway CA, Jr. (2005). Immunobiology (6. izd.). Garland Science. ISBN 0-443-07310-4.
- ↑ Liszewski MK; Farries TC; Lublin DM; Rooney IA; Atkinson JP (1996). »Control of the complement system«. Advances in Immunology. Advances in Immunology. 61: 201–83. doi:10.1016/S0065-2776(08)60868-8. ISBN 978-0-12-022461-6. PMID 8834497.
- ↑ Sim RB; Tsiftsoglou SA (Februar 2004). »Proteases of the complement system« (PDF). Biochemical Society Transactions. 32 (Pt 1): 21–7. doi:10.1042/BST0320021. PMID 14748705.
- ↑ Withers DR (Junij 2016). »Innate lymphoid cell regulation of adaptive immunity«. Immunology. 149: 123–30. doi:10.1111/imm.12639. PMC 5011676. PMID 27341319.
- ↑ Ryter A (1985). »Relationship between ultrastructure and specific functions of macrophages«. Comparative Immunology, Microbiology and Infectious Diseases. 8 (2): 119–33. doi:10.1016/0147-9571(85)90039-6. PMID 3910340.
- ↑ Langermans JA; Hazenbos WL; van Furth R (september 1994). »Antimicrobial functions of mononuclear phagocytes«. Journal of Immunological Methods. 174 (1–2): 185–94. doi:10.1016/0022-1759(94)90021-3. PMID 8083520.
{{navedi časopis}}: Vzdrževanje CS1: samodejni prevod datuma (povezava) - ↑ May RC; Machesky LM (Marec 2001). »Phagocytosis and the actin cytoskeleton«. Journal of Cell Science. 114 (Pt 6): 1061–77. doi:10.1242/jcs.114.6.1061. PMID 11228151.
- ↑ Salzet M; Tasiemski A; Cooper E (2006). »Innate immunity in lophotrochozoans: the annelids«. Current Pharmaceutical Design. 12 (24): 3043–50. doi:10.2174/138161206777947551. PMID 16918433.
- ↑ Zen K; Parkos CA (Oktober 2003). »Leukocyte-epithelial interactions«. Current Opinion in Cell Biology. 15 (5): 557–64. doi:10.1016/S0955-0674(03)00103-0. PMID 14519390.
- ↑ 43,0 43,1 Stvrtinová V, Jakubovský J, Hulín I (1995). Inflammation and Fever from Pathophysiology: Principles of Disease. Computing Centre, Slovak Academy of Sciences: Academic Electronic Press. ISBN 80-967366-1-2. Arhivirano iz prvotnega spletišča dne 11. julija 2001. Pridobljeno 1. januarja 2007.
- ↑ Rua R; McGavern DB (september 2015). »Elucidation of monocyte/macrophage dynamics and function by intravital imaging«. Journal of Leukocyte Biology. 98 (3): 319–32. doi:10.1189/jlb.4RI0115-006RR. PMC 4763596. PMID 26162402.
{{navedi časopis}}: Vzdrževanje CS1: samodejni prevod datuma (povezava) - ↑ 45,0 45,1 Guermonprez P; Valladeau J; Zitvogel L; Théry C; Amigorena S (2002). »Antigen presentation and T cell stimulation by dendritic cells«. Annual Review of Immunology. 20 (1): 621–67. doi:10.1146/annurev.immunol.20.100301.064828. PMID 11861614.
- ↑ Krishnaswamy G; Ajitawi O; Chi DS (2006). »The human mast cell: an overview«. Methods in Molecular Biology. 315: 13–34. doi:10.1385/1-59259-967-2:013. PMID 16110146.
- ↑ Kariyawasam HH; Robinson DS (april 2006). »The eosinophil: the cell and its weapons, the cytokines, its locations«. Seminars in Respiratory and Critical Care Medicine. 27 (2): 117–27. doi:10.1055/s-2006-939514. PMID 16612762.
{{navedi časopis}}: Vzdrževanje CS1: samodejni prevod datuma (povezava) - ↑ Middleton D; Curran M; Maxwell L (Avgust 2002). »Natural killer cells and their receptors«. Transplant Immunology. 10 (2–3): 147–64. doi:10.1016/S0966-3274(02)00062-X. PMID 12216946.
- ↑ Gabrielli S; Ortolani C; Del Zotto G; Luchetti F; Canonico B; Buccella F; Artico M; Papa S; Zamai L (2016). »The Memories of NK Cells: Innate-Adaptive Immune Intrinsic Crosstalk«. Journal of Immunology Research. 2016: 1376595. doi:10.1155/2016/1376595. PMC 5204097. PMID 28078307.
- ↑ Rajalingam R (2012). »Overview of the killer cell immunoglobulin-like receptor system«. Methods in Molecular Biology. Methods in Molecular Biology™. 882: 391–414. doi:10.1007/978-1-61779-842-9_23. ISBN 978-1-61779-841-2. PMID 22665247.
- ↑ Pancer Z; Cooper MD (2006). »The evolution of adaptive immunity«. Annual Review of Immunology. 24 (1): 497–518. doi:10.1146/annurev.immunol.24.021605.090542. PMID 16551257.
- ↑ 52,0 52,1 Holtmeier W; Kabelitz D (2005). »gammadelta T cells link innate and adaptive immune responses«. Chemical Immunology and Allergy. Chemical Immunology and Allergy. 86: 151–83. doi:10.1159/000086659. ISBN 3-8055-7862-8. PMID 15976493.
- ↑ Venturi, S.; Venturi, M. (2009). »Iodine, thymus, and immunity«. Nutrition. 25 (9): 977–979. doi:10.1016/j.nut.2009.06.002.
- ↑ Harty JT; Tvinnereim AR; White DW (2000). »CD8+ T cell effector mechanisms in resistance to infection«. Annual Review of Immunology. 18 (1): 275–308. doi:10.1146/annurev.immunol.18.1.275. PMID 10837060.
- ↑ 55,0 55,1 Radoja S; Frey AB; Vukmanovic S (2006). »T-cell receptor signaling events triggering granule exocytosis«. Critical Reviews in Immunology. 26 (3): 265–90. doi:10.1615/CritRevImmunol.v26.i3.40. PMID 16928189.
- ↑ Abbas AKMurphy KM; Sher A (Oktober 1996). »Functional diversity of helper T lymphocytes«. Nature. 383 (6603): 787–93. Bibcode:1996Natur.383..787A. doi:10.1038/383787a0. PMID 8893001.
- ↑ McHeyzer-Williams LJ; Malherbe LP; McHeyzer-Williams MG (2006). »Helper T cell-regulated B cell immunity«. Current Topics in Microbiology and Immunology. Current Topics in Microbiology and Immunology. 311: 59–83. doi:10.1007/3-540-32636-7_3. ISBN 978-3-540-32635-9. PMID 17048705.
- ↑ Kovacs B; Maus MV; Riley JL; Derimanov GS; Koretzky GA; June CH; Finkel TH (november 2002). »Human CD8+ T cells do not require the polarization of lipid rafts for activation and proliferation«. Proceedings of the National Academy of Sciences of the United States of America. 99 (23): 15006–11. Bibcode:2002PNAS...9915006K. doi:10.1073/pnas.232058599. PMC 137535. PMID 12419850.
{{navedi časopis}}: Vzdrževanje CS1: samodejni prevod datuma (povezava) - ↑ Grewal IS; Flavell RA (1998). »CD40 and CD154 in cell-mediated immunity«. Annual Review of Immunology. 16 (1): 111–35. doi:10.1146/annurev.immunol.16.1.111. PMID 9597126.
- ↑ Girardi M (Januar 2006). »Immunosurveillance and immunoregulation by gammadelta T cells«. The Journal of Investigative Dermatology. 126 (1): 25–31. doi:10.1038/sj.jid.5700003. PMID 16417214.
- ↑ »Understanding the Immune System: How it Works« (PDF). National Institute of Allergy and Infectious Diseases (NIAID). Arhivirano iz prvotnega spletišča (PDF) dne 3. januarja 2007. Pridobljeno 1. januarja 2007.
- ↑ Sproul TW; Cheng PC; Dykstra ML; Pierce SK (2000). »A role for MHC class II antigen processing in B cell development«. International Reviews of Immunology. 19 (2–3): 139–55. doi:10.3109/08830180009088502. PMID 10763706.
- ↑ Kehry MR; Hodgkin PD (1994). »B-cell activation by helper T-cell membranes«. Critical Reviews in Immunology. 14 (3–4): 221–38. doi:10.1615/CritRevImmunol.v14.i3-4.20. PMID 7538767.
- ↑ Murphy K, Weaver C (2016). »10: The Humoral Immune Response«. Immunobiology (9 izd.). Garland Science. ISBN 978-0-8153-4505-3.
- ↑ Alder MN; Rogozin IB; Iyer LM; Glazko GV; Cooper MD; Pancer Z (december 2005). »Diversity and function of adaptive immune receptors in a jawless vertebrate«. Science. 310 (5756): 1970–3. Bibcode:2005Sci...310.1970A. doi:10.1126/science.1119420. PMID 16373579.
{{navedi časopis}}: Vzdrževanje CS1: samodejni prevod datuma (povezava) - ↑ Saji F; Samejima Y; Kamiura S; Koyama M (Maj 1999). »Dynamics of immunoglobulins at the feto-maternal interface«. Reviews of Reproduction. 4 (2): 81–9. doi:10.1530/ror.0.0040081. PMID 10357095.
- ↑ Van de Perre P (Julij 2003). »Transfer of antibody via mother's milk«. Vaccine. 21 (24): 3374–6. doi:10.1016/S0264-410X(03)00336-0. PMID 12850343.
- ↑ Keller MA; Stiehm ER (Oktober 2000). »Passive immunity in prevention and treatment of infectious diseases«. Clinical Microbiology Reviews. 13 (4): 602–14. doi:10.1128/CMR.13.4.602-614.2000. PMC 88952. PMID 11023960.
- ↑ Death and DALY estimates for 2002 by cause for WHO Member States. World Health Organization. Retrieved on 1 January 2007.
- ↑ Singh M; O'Hagan D (november 1999). »Advances in vaccine adjuvants«. Nature Biotechnology. 17 (11): 1075–81. doi:10.1038/15058. PMID 10545912.
{{navedi časopis}}: Vzdrževanje CS1: samodejni prevod datuma (povezava) - ↑ Aw D; Silva AB; Palmer DB (april 2007). »Immunosenescence: emerging challenges for an ageing population«. Immunology. 120 (4): 435–46. doi:10.1111/j.1365-2567.2007.02555.x. PMC 2265901. PMID 17313487.
{{navedi časopis}}: Vzdrževanje CS1: samodejni prevod datuma (povezava) - ↑ Chandra RK (Avgust 1997). »Nutrition and the immune system: an introduction«. The American Journal of Clinical Nutrition. 66 (2): 460S–463S. PMID 9250133.
- ↑ Miller JF (Julij 2002). »The discovery of thymus function and of thymus-derived lymphocytes«. Immunological Reviews. 185 (1): 7–14. doi:10.1034/j.1600-065X.2002.18502.x. PMID 12190917.
- ↑ Joos L; Tamm M (2005). »Breakdown of pulmonary host defense in the immunocompromised host: cancer chemotherapy«. Proceedings of the American Thoracic Society. 2 (5): 445–8. doi:10.1513/pats.200508-097JS. PMID 16322598.
- ↑ Copeland KF; Heeney JL (december 1996). »T helper cell activation and human retroviral pathogenesis«. Microbiological Reviews. 60 (4): 722–42. PMC 239461. PMID 8987361.
{{navedi časopis}}: Vzdrževanje CS1: samodejni prevod datuma (povezava) - ↑ Miller JF (1993). »Self-nonself discrimination and tolerance in T and B lymphocytes«. Immunologic Research. 12 (2): 115–30. doi:10.1007/BF02918299. PMID 8254222.
- ↑ 77,0 77,1 Ghaffar, Abdul (2006). »Immunology — Chapter Seventeen: Hypersensitivity States«. Microbiology and Immunology On-line. University of South Carolina School of Medicine. Pridobljeno 29. maja 2016.
- ↑ Bickle TA; Krüger DH (Junij 1993). »Biology of DNA restriction«. Microbiological Reviews. 57 (2): 434–50. PMC 372918. PMID 8336674.
- ↑ Barrangou R; Fremaux C; Deveau H; Richards M; Boyaval P; Moineau S; Romero DA; Horvath P (Marec 2007). »CRISPR provides acquired resistance against viruses in prokaryotes«. Science. 315 (5819): 1709–12. Bibcode:2007Sci...315.1709B. doi:10.1126/science.1138140. PMID 17379808.
- ↑ Hille, Frank; Charpentier, Emmanuelle (2016). »CRISPR-Cas: biology, mechanisms and relevance«. Philosophical Transactions of the Royal Society B. 371: 20150496. doi:10.1098/rstb.2015.0496.
- ↑ Koonin, Eugene V. (2017). »Evolution of RNA- and DNA-guided antivirus defense systems in prokaryotes and eukaryotes: common ancestry vs convergence«. Biology Direct. 12: 5. doi:10.1186/s13062-017-0177-2. ISSN 1745-6150. PMC 5303251. PMID 28187792.
- ↑ Bayne CJ (2003). »Origins and evolutionary relationships between the innate and adaptive arms of immune systems«. Integr. Comp. Biol. 43 (2): 293–299. doi:10.1093/icb/43.2.293. PMID 21680436.
- ↑ Stram Y; Kuzntzova L (Junij 2006). »Inhibition of viruses by RNA interference«. Virus Genes. 32 (3): 299–306. doi:10.1007/s11262-005-6914-0. PMID 16732482.
- ↑ Beck, Gregory; Habicht, Gail S. (1996). »Immunity and the Invertebrates« (PDF). Scientific American. 275 (5): 60–66. doi:10.1038/scientificamerican1196-60.
- ↑ Schneider, David. »Innate Immunity — Lecture 4: Plant immune responses« (PDF). Stanford University Department of Microbiology and Immunology. Pridobljeno 1. januarja 2007.
- ↑ Jones JD; Dangl JL (november 2006). »The plant immune system«. Nature. 444 (7117): 323–9. Bibcode:2006Natur.444..323J. doi:10.1038/nature05286. PMID 17108957.
{{navedi časopis}}: Vzdrževanje CS1: samodejni prevod datuma (povezava) - ↑ Jones, Jonathan D.G.; Dangl, Jeffery L. (2006). »The plant immune system«. Nature. Zv. 444. str. 323–329. doi:10.1038/nature05286.
- ↑ Baulcombe D (september 2004). »RNA silencing in plants«. Nature. 431 (7006): 356–63. Bibcode:2004Natur.431..356B. doi:10.1038/nature02874. PMID 15372043.
{{navedi časopis}}: Vzdrževanje CS1: samodejni prevod datuma (povezava) - ↑ Morgan, Richard A.; Dudley, Mark E.; Wunderlich, John R.; Hughes, Marybeth S.; Yang, James C.; in sod. (Oktober 2006). »Cancer Regression in Patients After Transfer of Genetically Engineered Lymphocytes«. Science. 314 (5796): 126–9. Bibcode:2006Sci...314..126M. doi:10.1126/science.1129003. PMC 2267026. PMID 16946036.
- ↑ Andersen MH; Schrama D; Thor Straten P; Becker JC (Januar 2006). »Cytotoxic T cells«. The Journal of Investigative Dermatology. 126 (1): 32–41. doi:10.1038/sj.jid.5700001. PMID 16417215.
- ↑ Boon T; van der Bruggen P (Marec 1996). »Human tumor antigens recognized by T lymphocytes«. The Journal of Experimental Medicine. 183 (3): 725–9. doi:10.1084/jem.183.3.725. PMC 2192342. PMID 8642276.
- ↑ Castelli C; Rivoltini L; Andreola G; Carrabba M; Renkvist N; Parmiani G (Marec 2000). »T-cell recognition of melanoma-associated antigens«. Journal of Cellular Physiology. 182 (3): 323–31. doi:10.1002/(SICI)1097-4652(200003)182:3<323::AID-JCP2>3.0.CO;2-#. PMID 10653598.
- ↑ Romero P; Cerottini JC; Speiser DE (2006). »The human T cell response to melanoma antigens«. Advances in Immunology. Advances in Immunology. 92: 187–224. doi:10.1016/S0065-2776(06)92005-7. ISBN 978-0-12-373636-9. PMID 17145305.
- ↑ Guevara-Patiño JA; Turk MJ; Wolchok JD; Houghton AN (2003). »Immunity to cancer through immune recognition of altered self: studies with melanoma«. Advances in Cancer Research. Advances in Cancer Research. 90: 157–77. doi:10.1016/S0065-230X(03)90005-4. ISBN 978-0-12-006690-2. PMID 14710950.
- ↑ Renkvist N; Castelli C; Robbins PF; Parmiani G (Marec 2001). »A listing of human tumor antigens recognized by T cells«. Cancer Immunology, Immunotherapy. 50 (1): 3–15. doi:10.1007/s002620000169. PMID 11315507.
- ↑ Gerloni M; Zanetti M (Junij 2005). »CD4 T cells in tumor immunity«. Springer Seminars in Immunopathology. 27 (1): 37–48. doi:10.1007/s00281-004-0193-z. PMID 15965712.
- ↑ Seliger B; Ritz U; Ferrone S (Januar 2006). »Molecular mechanisms of HLA class I antigen abnormalities following viral infection and transformation«. International Journal of Cancer. 118 (1): 129–38. doi:10.1002/ijc.21312. PMID 16003759.
- ↑ Hayakawa Y; Smyth MJ (2006). »Innate immune recognition and suppression of tumors«. Advances in Cancer Research. Advances in Cancer Research. 95: 293–322. doi:10.1016/S0065-230X(06)95008-8. ISBN 978-0-12-006695-7. PMID 16860661.
- ↑ Syn, Nicholas L; Teng, Michele W L; Mok, Tony S K; Soo, Ross A (2017). »De-novo and acquired resistance to immune checkpoint targeting«. The Lancet Oncology (v angleščini). 18 (12): e731–e741. doi:10.1016/s1470-2045(17)30607-1.
- ↑ Seliger B (2005). »Strategies of tumor immune evasion«. BioDrugs. 19 (6): 347–54. doi:10.2165/00063030-200519060-00002. PMID 16392887.
- ↑ Frumento G; Piazza T; Di Carlo E; Ferrini S (september 2006). »Targeting tumor-related immunosuppression for cancer immunotherapy«. Endocrine, Metabolic & Immune Disorders Drug Targets. 6 (3): 233–7. doi:10.2174/187153006778250019. PMID 17017974.
{{navedi časopis}}: Vzdrževanje CS1: samodejni prevod datuma (povezava) - ↑ Stix G (Julij 2007). »A malignant flame. Understanding chronic inflammation, which contributes to heart disease, Alzheimer's and a variety of other ailments, may be a key to unlocking the mysteries of cancer« (PDF). Scientific American. 297 (1): 60–7. doi:10.1038/scientificamerican0707-60. PMID 17695843. Arhivirano iz prvotnega spletišča (PDF) dne 16. julija 2011.
- ↑ WICK, G.; HU, Y.; SCHWARZ, S.; KROEMER, G. (1. oktober 1993). »Immunoendocrine Communication via the Hypothalamo-Pituitary-Adrenal Axis in Autoimmune Diseases*«. Endocrine Reviews (v angleščini). 14 (5): 539–563. doi:10.1210/edrv-14-5-539. ISSN 0163-769X.
- ↑ Kroemer, Guido; Brezinschek, Hans-Peter; Faessler, Reinhard; Schauenstein, Konrad; Wick, Georg (1. januar 1988). »Physiology and pathology of an immunoendocrine feedback loop«. Immunology Today. 9 (6): 163–165. doi:10.1016/0167-5699(88)91289-3.
- ↑ Trakhtenberg, Ephraim F.; Goldberg, Jeffrey L. (7. oktober 2011). »Neuroimmune Communication«. Science (v angleščini). 334 (6052): 47–48. doi:10.1126/science.1213099. ISSN 0036-8075. PMID 21980100.
- ↑ Veiga-Fernandes, Henrique; Mucida, Daniel (5. maj 2016). »Neuro-Immune Interactions at Barrier Surfaces«. Cell. 165 (4): 801–811. doi:10.1016/j.cell.2016.04.041. ISSN 1097-4172. PMC 4871617. PMID 27153494.
- ↑ »Neuroimmune communication«. Nature Neuroscience (v angleščini). 20 (2): 127–127. Februar 2017. doi:10.1038/nn.4496. ISSN 1097-6256.
- ↑ Wira CR, Crane-Godreau M, Grant K (2004). »Endocrine regulation of the mucosal immune system in the female reproductive tract«. V Ogra PL, Mestecky J, Lamm ME, Strober W, McGhee JR, Bienenstock J (ur.). Mucosal Immunology. San Francisco: Elsevier. ISBN 0-12-491543-4.
- ↑ Lang TJ (december 2004). »Estrogen as an immunomodulator«. Clinical Immunology. 113 (3): 224–30. doi:10.1016/j.clim.2004.05.011. PMID 15507385.
{{navedi časopis}}: Vzdrževanje CS1: samodejni prevod datuma (povezava)
Moriyama A; Shimoya K; Ogata I; Kimura T; Nakamura T; Wada H; Ohashi K; Azuma C; Saji F; Murata Y (Julij 1999). »Secretory leukocyte protease inhibitor (SLPI) concentrations in cervical mucus of women with normal menstrual cycle«. Molecular Human Reproduction. 5 (7): 656–61. doi:10.1093/molehr/5.7.656. PMID 10381821.
Cutolo M; Sulli A; Capellino S; Villaggio B; Montagna P; Seriolo B; Straub RH (2004). »Sex hormones influence on the immune system: basic and clinical aspects in autoimmunity«. Lupus. 13 (9): 635–8. doi:10.1191/0961203304lu1094oa. PMID 15485092.
King AE; Critchley HO; Kelly RW (Februar 2000). »Presence of secretory leukocyte protease inhibitor in human endometrium and first trimester decidua suggests an antibacterial protective role«. Molecular Human Reproduction. 6 (2): 191–6. doi:10.1093/molehr/6.2.191. PMID 10655462. - ↑ Fimmel S; Zouboulis CC (2005). »Influence of physiological androgen levels on wound healing and immune status in men«. The Aging Male. 8 (3–4): 166–74. doi:10.1080/13685530500233847. PMID 16390741.
- ↑ Dorshkind K; Horseman ND (Junij 2000). »The roles of prolactin, growth hormone, insulin-like growth factor-I, and thyroid hormones in lymphocyte development and function: insights from genetic models of hormone and hormone receptor deficiency«. Endocrine Reviews. 21 (3): 292–312. doi:10.1210/er.21.3.292. PMID 10857555.
- ↑ Nagpal S; Na S; Rathnachalam R (Avgust 2005). »Noncalcemic actions of vitamin D receptor ligands«. Endocrine Reviews. 26 (5): 662–87. doi:10.1210/er.2004-0002. PMID 15798098.
- ↑ »Vitamin D controls T cell antigen receptor signaling and activation of human T cells«. Nature Immunology. Zv. 11, št. 4. april 2010. str. 344–9. doi:10.1038/ni.1851. PMID 20208539.
{{navedi revijo}}: Vzdrževanje CS1: samodejni prevod datuma (povezava) - ↑ »DCs metabolize sunlight-induced vitamin D3 to 'program' T cell attraction to the epidermal chemokine CCL27«. Marec 2007. str. 285–93. doi:10.1038/ni1433. PMID 17259988. Arhivirano iz prvotnega spletišča dne 9. maja 2014. Pridobljeno 28. aprila 2014.
- ↑ »The "multiple hormone deficiency" theory of aging: is human senescence caused mainly by multiple hormone deficiencies?«. Annals of the New York Academy of Sciences. Zv. 1057, št. 1. december 2005. str. 448–65. Bibcode:2005NYASA1057..448H. doi:10.1196/annals.1322.035. PMID 16399912.
{{navedi revijo}}: Vzdrževanje CS1: samodejni prevod datuma (povezava) - ↑ »The immune system as a regulator of thyroid hormone activity«. Experimental Biology and Medicine. Zv. 231, št. 3. Marec 2006. str. 229–36. doi:10.1016/j.plefa.2005.04.005. PMC 2768616. PMID 16514168.
- ↑ »Vitamin D and the elderly«. Clinical Endocrinology. Zv. 62, št. 3. Marec 2005. str. 265–81. doi:10.1111/j.1365-2265.2005.02226.x. PMID 15730407.
- ↑ »Sleep enhances the human antibody response to hepatitis A vaccination« (PDF). Psychosomatic Medicine. Zv. 65, št. 5. 2003. str. 831–5. doi:10.1097/01.PSY.0000091382.61178.F1. PMID 14508028.
- ↑ »Sick and tired: Does sleep have a vital role in the immune system?«. Nature Reviews. Immunology. Zv. 4, št. 6. Junij 2004. str. 457–67. doi:10.1038/nri1369. PMID 15173834.
- ↑ »Humoral links between sleep and the immune system: research issues«. Annals of the New York Academy of Sciences. Zv. 992, št. 1. Maj 2003. str. 9–20. Bibcode:2003NYASA.992....9K. doi:10.1111/j.1749-6632.2003.tb03133.x. PMID 12794042.
- ↑ »Links between the innate immune system and sleep«. The Journal of Allergy and Clinical Immunology. Zv. 116, št. 6. december 2005. str. 1188–98. doi:10.1016/j.jaci.2005.08.005. PMID 16337444.
{{navedi revijo}}: Vzdrževanje CS1: samodejni prevod datuma (povezava) - ↑ Burzyn, Dalia; Kuswanto, Wilson; Kolodin, Dmitriy; Shadrach, Jennifer L.; Cerletti, Massimiliano; Jang, Young; Sefik, Esen; Tan, Tze Guan; Wagers, Amy J. (5. december 2013). »Sleep's Effects On Your Immune System Revealed In New Body Clock Study«. Cell (v angleščini). Zv. 155, št. 6. str. 1282–1295. doi:10.1016/j.cell.2013.10.054. ISSN 0092-8674. PMC 3894749. PMID 24315098. Pridobljeno 28. aprila 2014.
- ↑ 123,0 123,1 Wynn, Thomas A.; Vannella, Kevin M. (Januar 2012). »Sleep and immune function«. Pflügers Archiv (v angleščini). Zv. 463, št. 1. str. 121–37. doi:10.1007/s00424-011-1044-0. ISSN 1074-7613. PMC 3256323. PMID 22071480.
- ↑ »Can Better Sleep Mean Catching fewer Colds?«. Arhivirano iz prvotnega spletišča dne 9. maja 2014. Pridobljeno 28. aprila 2014.
- ↑ R.M. Suskind, C.L. Lachney, J.N. Udall, Jr., "Malnutrition and the Immune Response", in: Dairy products in human health and nutrition, M. Serrano-Ríos, ed., CRC Press, 1994, pp. 285–300
- ↑ »Adipose tissue and the immune system«. Prostaglandins, Leukotrienes, and Essential Fatty Acids. Zv. 73, št. 1. Julij 2005. str. 17–30. doi:10.1016/j.plefa.2005.04.005. PMID 15946832.
- ↑ »Diet and the developing immune system«. Lupus. Zv. 15, št. 11. 2006. str. 746–52. doi:10.1177/0961203306070001. PMID 17153845.
- ↑ Park, Julie E.; Barbul, Adrian (1. maj 2004). Understanding the role of immune regulation in wound healing (v angleščini). Zv. 187. New York: Oxford University Press. str. S11–S16. doi:10.1016/s0002-9610(03)00296-4. ISSN 0002-9610. PMID 15147986.
- ↑ Burzyn, Dalia; Kuswanto, Wilson; Kolodin, Dmitriy; Shadrach, Jennifer L. (5. december 2013). A Special Population of Regulatory T Cells Potentiates Muscle Repair (v angleščini). Zv. 155. Cambridge: Cambridge University Press. str. 1282–1295. doi:10.1016/j.cell.2013.10.054. ISSN 0092-8674. PMC 3894749. PMID 24315098.
- ↑ Leoni, G.; Neumann, P.-A.; Sumagin, R.; Denning, T. L.; Nusrat, A. (september 2015). »Wound repair: role of immune–epithelial interactions«. Mucosal Immunology (v angleščini). Zv. 8, št. 5. str. 959–968. doi:10.1038/mi.2015.63. ISSN 1933-0219. PMC 4916915. PMID 26174765.
{{navedi revijo}}: Vzdrževanje CS1: samodejni prevod datuma (povezava) - ↑ Wynn, Thomas A.; Vannella, Kevin M. (15. marec 2016). »Macrophages in Tissue Repair, Regeneration, and Fibrosis«. Immunity (v angleščini). Zv. 44, št. 3. str. 450–462. doi:10.1016/j.immuni.2016.02.015. ISSN 1074-7613. PMC 4794754. PMID 26982353.
- ↑ 132,0 132,1 Laurent, Paôline; Jolivel, Valérie; Manicki, Pauline; Chiu, Lynn; Contin-Bordes, Cécile; Truchetet, Marie-Elise; Pradeu, Thomas (2017). »Immune-Mediated Repair: A Matter of Plasticity«. Frontiers in Immunology (v angleščini). 8. doi:10.3389/fimmu.2017.00454. ISSN 1664-3224. PMC 5403426. PMID 28484454.
- ↑ Eming, Sabine A.; Hammerschmidt, Matthias; Krieg, Thomas; Roers, Axel (1. julij 2009). »Interrelation of immunity and tissue repair or regeneration«. Seminars in Cell & Developmental Biology. Regenerative Biology and Medicine: INuclear Transport in Development and Disease. Zv. 20, št. 5. str. 517–527. doi:10.1016/j.semcdb.2009.04.009.
- ↑ Godwin, James W.; Pinto, Alexander R.; Rosenthal, Nadia A. (Januar 2017). »Chasing the recipe for a pro-regenerative immune system«. Seminars in Cell & Developmental Biology. Innate immune pathways in wound healingPeromyscus as a model system. Zv. 61. str. 71–79. doi:10.1016/j.semcdb.2016.08.008. PMC 5338634. PMID 27521522.
- ↑ 135,0 135,1 »Immunosuppressive agents in solid organ transplantation: Mechanisms of action and therapeutic efficacy«. Critical Reviews in Oncology/Hematology. Zv. 56, št. 1. Oktober 2005. str. 23–46. doi:10.1016/j.critrevonc.2005.03.012. PMC 534292. PMID 16039869.
- ↑ Burnet FM (Marec 2006). Corticosteroids: the drugs to beat. Zv. 533. Cambridge: Cambridge University Press. str. 2–14. doi:10.1016/j.ejphar.2005.12.052. PMID 16436275.
- ↑ Pradeu (Julij 2003). The mosaic of immunosuppressive drugs. Zv. 39. New York: Oxford University Press. str. 1073–7. doi:10.1016/S0161-5890(03)00075-0. PMID 12835079.
- ↑ Silverstein A (1989). A History of Immunology. New York: Academic Press.
- ↑ Tauber AI; Chernyak L (1991). Metchnikoff and the Origins of Immunology. New York: Oxford University Press.
- ↑ Tauber AI (1994). The Immune Self: Theory or Metaphor?. Cambridge: Cambridge University Press.
- ↑ »THE NATURAL-SELECTION THEORY OF ANTIBODY FORMATION«. Proceedings of the National Academy of Sciences of the United States of America. Zv. 41, št. 11. november 1955. str. 849–57. doi:10.1073/pnas.41.11.849. PMC 534292. PMID 16589759.
{{navedi revijo}}: Vzdrževanje CS1: samodejni prevod datuma (povezava) - ↑ Burnet FM, Thomas; Vivier, Eric (1959). »The Clonal Selection Theory of Acquired Immunity«. Science Immunology (v angleščini). Zv. 1, št. 1. Cambridge: Cambridge University Press. str. aag0479–aag0479. doi:10.1126/sciimmunol.aag0479. ISSN 2470-9468. PMC 5321532. PMID 28239677. Arhivirano iz prvotnega spletišča dne 14. maja 2021. Pridobljeno 9. februarja 2022.
- ↑ Burnet FM (1969). »Cellular Immunology: Self and Notself«. Current Biology. Zv. 6, št. 5. Cambridge: Cambridge University Press. str. 519–22. doi:10.1016/S0960-9822(02)00531-6. PMID 8805259.
- ↑ Bretscher P; Cohn M (1970). »A theory of self-nonself discrimination«. Science. Zv. 169, št. 3950. str. 1042–49. doi:10.1126/science.169.3950.1042.
- ↑ »The danger model: a renewed sense of self« (PDF). Science. Zv. 296, št. 5566. april 2002. str. 301–5. doi:10.1126/science.1071059. PMC 3443751. PMID 11951032.
{{navedi revijo}}: Vzdrževanje CS1: samodejni prevod datuma (povezava) - ↑ Pradeu (2012). The Limits of the Self: Immunology and Biological Identity. New York: Oxford University Press.
- ↑ »A minimal model for the self-nonself discrimination: a return to the basics«. Seminars in Immunology. Zv. 12, št. 3. Junij 2000. str. 189–95, discussion 257–344. doi:10.1006/smim.2000.0231. PMID 10910739.
- ↑ Clark WR (2008). In Defense of Self: How the Immune System Really Works. New York: Oxford University Press.
- ↑ Coutinho A; in sod. (1984). »From an antigen-centered, clonal perspective of immune responses to an organism-centered network perspective of autonomous reactivity of self-referential immune systems«. Immunological Reviews. Zv. 79. str. 151–168. doi:10.1111/j.1600-065x.1984.tb00492.x.
- ↑ Irun C (2000). Tending Adam’s garden: Evolving the cognitive immune self. San Diego: Academic Press.
- ↑ »On the definition of a criterion of immunogenicity«. Proceedings of the National Academy of Sciences of the United States of America. Zv. 103, št. 47. november 2006. str. 17858–61. doi:10.1073/pnas.0608683103. PMC 1693837. PMID 17101995.
{{navedi revijo}}: Vzdrževanje CS1: samodejni prevod datuma (povezava) - ↑ »The speed of change: towards a discontinuity theory of immunity?«. Nature Reviews. Immunology. Zv. 13, št. 10. Oktober 2013. str. 764–9. doi:10.1038/nri3521. PMID 23995627.
- ↑ Pradeu, Thomas; Vivier, Eric (14. julij 2016). »The discontinuity theory of immunity«. Science Immunology (v angleščini). Zv. 1, št. 1. str. aag0479–aag0479. doi:10.1126/sciimmunol.aag0479. ISSN 2470-9468. PMC 5321532. PMID 28239677. Arhivirano iz prvotnega spletišča dne 14. maja 2021. Pridobljeno 9. februarja 2022.
- ↑ »Danger - pathogen on the premises! Immunological tolerance«. Current Biology. Zv. 6, št. 5. Maj 1996. str. 519–22. doi:10.1016/S0960-9822(02)00531-6. PMID 8805259.
- ↑ Vance RE (2000). »Cutting edge commentary: a Copernican revolution? Doubts about the danger theory«. Journal of Immunology. Zv. 165, št. 4. str. 1725–1728. doi:10.4049/jimmunol.165.4.1725.
- ↑ »The evolution of the danger theory. Interview by Lauren Constable, Commissioning Editor«. Expert Review of Clinical Immunology. Zv. 8, št. 4. Maj 2012. str. 311–7. doi:10.1586/eci.12.21. PMID 22607177.
- ↑ »The danger theory: 20 years later«. Frontiers in Immunology. Zv. 3. 2012. str. 287. doi:10.3389/fimmu.2012.00287. PMC 3443751. PMID 23060876.
- ↑ »Prediction of sequential antigenic regions in proteins«. FEBS Letters. Zv. 188, št. 2. september 1985. str. 215–8. doi:10.1016/0014-5793(85)80374-4. PMID 2411595.
{{navedi revijo}}: Vzdrževanje CS1: samodejni prevod datuma (povezava) - ↑ »Machine learning approaches for prediction of linear B-cell epitopes on proteins«. Journal of Molecular Recognition. Zv. 19, št. 3. 2006. str. 200–8. doi:10.1002/jmr.771. PMID 16598694.
- ↑ »Bcipep: a database of B-cell epitopes«. BMC Genomics. Zv. 6. 2005. str. 79. doi:10.1186/1471-2164-6-79. PMC 1173103. PMID 15921533.
- ↑ »Immunoinformatics and the prediction of immunogenicity«. Applied Bioinformatics. Zv. 1, št. 4. 2002. str. 167–76. PMID 15130835.
- ↑ »Anti-immunology: evasion of the host immune system by bacterial and viral pathogens«. Cell. Zv. 124, št. 4. Februar 2006. str. 767–82. doi:10.1016/j.cell.2006.01.034. PMID 16497587.
- ↑ »Type II secretion: a protein secretion system for all seasons«. Trends in Microbiology. Zv. 13, št. 12. december 2005. str. 581–8. doi:10.1016/j.tim.2005.09.005. PMID 16216510.
{{navedi revijo}}: Vzdrževanje CS1: samodejni prevod datuma (povezava) - ↑ »Type III secretion systems and pathogenicity islands«. Journal of Medical Microbiology. Zv. 50, št. 2. Februar 2001. str. 116–26. doi:10.1099/0022-1317-50-2-116. PMID 11211218.
- ↑ »Common themes in microbial pathogenicity revisited«. Microbiology and Molecular Biology Reviews. Zv. 61, št. 2. Junij 1997. str. 136–69. PMC 232605. PMID 9184008.
- ↑ »Airway biofilms: implications for pathogenesis and therapy of respiratory tract infections«. Treatments in Respiratory Medicine. Zv. 4, št. 4. 2005. str. 241–53. doi:10.2165/00151829-200504040-00003. PMID 16086598.
- ↑ »Immunoglobulin-binding domains: Protein L from Peptostreptococcus magnus« (PDF). Biochemical Society Transactions. Zv. 31, št. Pt 3. Junij 2003. str. 716–8. doi:10.1042/BST0310716. PMID 12773190.
- ↑ »Antibody vs. HIV in a clash of evolutionary titans«. Proceedings of the National Academy of Sciences of the United States of America. Zv. 102, št. 42. Oktober 2005. str. 14943–8. Bibcode:2005PNAS..10214943B. doi:10.1073/pnas.0505126102. PMC 1257708. PMID 16219699.
- ↑ »Switching trypanosome coats: what's in the wardrobe?«. Trends in Genetics. Zv. 22, št. 11. november 2006. str. 614–20. doi:10.1016/j.tig.2006.08.003. PMID 16908087.
{{navedi revijo}}: Vzdrževanje CS1: samodejni prevod datuma (povezava) - ↑ »Plunder and stowaways: incorporation of cellular proteins by enveloped viruses«. Journal of Virology. Zv. 79, št. 11. Junij 2005. str. 6577–87. doi:10.1128/JVI.79.11.6577-6587.2005. PMC 1112128. PMID 15890896.
Glej tudi
[uredi | uredi kodo]Zunanje povezave
[uredi | uredi kodo]- Imunski Sistem Arhivirano 2019-05-02 na Wayback Machine. – iz Univerze v Hartford (visoka šola/dodiplomski ravni)
- Za mikrobiologijo in Imunologijo On-Line, Učbenik, – iz University of South Carolina School of Medicine (dodiplomski ravni)
- Immunobiology; Peta Izdaja – Online različici učbenika za Charles Janeway (Advanced dodiplomski/podiplomski ravni)
- Stanley Falkow je govoril: "Gostitelj-Patogen Interakcije in Človeških Bolezni"