Рилин
| Рилин | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | uniprot:P78509RELN | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние ID | GeneCards: [1] | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
Рили́н (англ. reelin) — белок, содержащийся в мозге и в других тканях и органах тела человека и других животных. Этот гликопротеин выполняет множество функций, важнейшей из которых является регулировка миграции и позиционирования нервных стволовых клеток в период фетального и раннего послеродового развития, необходимая для нормального формирования коры и других структур головного мозга. Во взрослом мозге рилин регулирует позиционирование нейронов, образуемых в процессе взрослого нейрогенеза, а также вносит вклад в работу механизмов памяти и обучения, модулируя синаптическую пластичность, усиливая и поддерживая долговременную потенциацию[1][2], стимулируя развитие дендритов[3][4] и дендритных шипиков[5].
Название «рилин» происходит от английского глагола to reel — кружиться, вертеться, идти нетвёрдой походкой. Именно такая, «закрученная», неровная походка была отмечена у мышей с генетически обусловленным недостатком рилина. Острая нехватка белка ведет к нарушению миграции нейронов. Если ген, кодирующий синтез рилина, отключён полностью (гомозиготный генотип), наблюдается инверсия слоев коры головного мозга. При гетерозиготном генотипе нарушения мозга у мышей менее заметны, однако напоминают нарушения человеческого мозга при психотических расстройствах[6][7]. У людей генетически обусловленное отсутствие рилина приводит к лиссэнцефалии, тяжелой умственной отсталости и эпилепсии. Значительный недостаток рилина наблюдается при посмертных исследованиях мозга людей, которым при жизни были поставлены диагнозы шизофрении и биполярного расстройства[8], однако следует отметить возможное воздействие медикаментов[9]. Есть данные о возможной связи полиморфизмов гена RELN с шизофренией[10] и болезнью Альцгеймера[11].
История открытия и исследований
[править | править код] |
 |

Исследование мышей-мутантов позволило ученым заглянуть в глубинные механизмы развития центральной нервной системы. Идентификацией спонтанных мутаций у мышей впервые занялись нейрофизиологи, исследовавшие моторное поведение. Обнаружить нужных мышей в помёте было относительно легко: мутанты не были способны нормально передвигаться по клетке. Было найдено несколько подобных мышей, получивших имена в соответствии с характером нарушений моторики — reeler («крутящийся»), weaver («качающийся»), lurcher («кренящийся»), nervous («нервный»), и staggerer(«пошатывающийся»).
Мышь, названная reeler, была впервые описана в 1951 году британским генетиком Дугласом Скоттом Фальконером.[12] В 1960-е годы было обнаружено, что мозжечок у этих мышей намного меньше нормы, к тому же нарушена нормальная организация нейрональных слоёв.[13] Мутация особенно заинтересовала исследователей после того, как было обнаружено, что слои нейронов у мышей выстраиваются «наоборот»: более молодые нейроны были не состоянии преодолеть слои уже «осевших» на своем уровне клеток.[14]
В 1994 году положение гена было уточнено с помощью инсерционного мутагенеза,[15] что позволило в 1995 году обнаружить ген RELN, располагающийся на хромосоме 7q22.[16] В том же году японскими учеными из медицинской школы города Коти было успешно создано первое моноклональное антитело к рилину, названное CR-50.[17] Они отметили, что клетки Кахаля — Ретциуса, функция которых к тому времени была неизвестна, демонстрировали особенно сильную реакцию на CR-50.
Клеточные рецепторы, реагирующие на рилин, апоЕ-рецептор 2 (apolipoprotein E receptor 2, apoER2) и рецептор липопротеинов очень низкой плотности (very-low-density lipoprotein receptor, VLDLR), были обнаружены случайно в ходе эксперимента, проводимого Тромсдорфом и коллегами в 1997 году.[18] У использованных в эксперименте мутантов, так называемых «нокаутных» мышей с отсутствующими рецепторами apoER2 и VLDLR, обнаружились дефекты в строении коры головного мозга, идентичные дефектам мыши reeler.
С целью глубже изучить механизм сигнального пути белка рилина и найти остальные его элементы, ученые использовали два других типа мутантных мышей — yotari и scrambler. Эти мыши по фенотипу схожи с мышью reeler, но сам ген RELN, кодирующий белок рилин, у них нормален. Исследования этих мутантов выявили нарушения в гене DAB1, который кодирует одноимённый протеин. Мыши yotari, как оказалось, были лишены белка Dab1 полностью, а у мышей scrambler его удавалось обнаружить с трудом.[19] Целенаправленное разрушение гена DAB1 также вызывало фенотип, аналогичный фенотипу мыши reeler. Определение того, что именно DAB1 является ключевым регулятором сигнального каскада рилина, положило начало доскональному изучению его сложных внутриклеточных взаимодействий.
Открытие возможной связи рилина с шизофренией и биполярным расстройством, болезнью Альцгеймера и аутизмом, другими дисфункциями, а также перспектива раскрытия механизмов, обусловивших возникновение сложно организованного человеческого мозга, привели к активному исследованию белка и его сигнальных взаимодействий. К началу второго десятилетия после открытия гена RELN, число научных статей о рилине исчислялось сотнями,[20] а в 2008 году был опубликован сборник, многочисленные авторы которого рассматривают различные структурные и функциональные особенности рилина в норме и при патологиях.[9]
Секреция и локализация белка
[править | править код]
Рилин является секретируемым элементом внеклеточного матрикса. Скорость секреции рилина связана со скоростью его производства и не зависит от деполяризации мембраны. Рилин обнаруживается в секреторных гранулах аппарата Гольджи и отсутствует в синаптических пузырьках, что характерно для белков внеклеточного матрикса.
В период развития мозга рилин в коре головного мозга и гиппокампе синтезируется клетками Кахаля — Ретциуса, а также клетками Кахаля и клетками Ретциуса.[21] Рилин-производящие клетки в пренатальном и раннем постнатальном мозге преимущественно размещаются в маргинальной зоне (англ. marginal zone, MZ) коры и во временном субпиальном гранулярном слое (англ. subpial granular layer, SGL), наиболее развитом у человека,[22] а в гиппокампе — в слое stratum lacunosum-moleculare и верхнем маргинальном слое зубчатой извилины.
В мозжечке рилин производится во внешнем слое гранулярных клеток перед миграцией гранулярных клеток во внутренний слой.[23]
В целом в постнатальном периоде происходит переход от послойной к диффузной экспрессии рилина. Во взрослом мозге белок синтезируется корковыми ГАМК-ергическими интернейронами, экспрессирующими калретинин и калбиндин, такими как клетки Мартинотти. Парвальбумин-содержащие ГАМКергические интернейроны, такие как канделяберные и корзинчатые клетки, никогда не производят рилин либо делают это крайне редко.[24][25] В мозжечке у взрослых рилин производится глутаматергическими гранулярными нейронами внутреннего слоя.[26] Некоторые из немногочисленных нейронов, расположенных в толще белого вещества, также синтезируют рилин.[27]
За пределами мозга рилин у взрослых млекопитающих обнаруживается в крови, печени, средней доле гипофиза и хромаффинных клетках надпочечников.[28] В печени рилин был обнаружен в клетках Ито.[29] При повреждении печени уровень мРНК белка резко повышается и снижается по завершении процесса восстановления.[30]
В глазах рилин вырабатывается в слое ганглионарных клеток сетчатки и эндотелиальном слое роговицы.[31] Как и в печени, экспрессия белка повышается при повреждениях.
Рилин производится также одонтобластами, клетками, расположенными на периферии зубной пульпы. Белок обнаруживается здесь как в процессе одонтогенеза, так и во взрослом зубе.[32] Как предполагают некоторые авторы, одонтобласты являются сенсорными клетками, способными передавать болевые сигналы нервным окончаниям.[33] Согласно их гипотезе, рилин играет роль в этом процессе,[9] так как он может способствовать установлению контакта одонтобластов с нервными терминалями.[34]

Структура белка
[править | править код]
Рилин состоит из 3461 аминокислот и имеет относительную молекулярную массу 388 кДа. В мышином гене RELN 65 экзонов, простирающихся примерно на 450 kb.[36] N-терминальные экзоны разделены большими интронами, остальные расположены ближе друг к другу. Один экзон, кодирующий всего две аминокислоты рядом с C-окончанием, подвержен альтернативному сплайсингу, но неясно, как это отражается на функции белка.[9] В структуре гена идентифицированы два основных сайта инициации транскрипции и два участка полиаденилирования.[36]
Молекула белка начинается сигнальным пептидом длиной в 27 аминокислотных остатков. Затем следует участок, по строению схожий с белком внеклеточной матрицы F-спондином (на схеме отмечен как SP, аминокислотные остатки 28—190). Затем — уникальный для рилина участок (сегмент H на схеме), после которого идут подряд 8 участков похожей структуры, так называемые «рилиновые повторы», длиной примерно 350 аминокислот каждый. В центре каждого повтора располагается EGF-подобное включение, делящее повтор на два субповтора, A и B, структура которых имеет мало общего. Несмотря на разделение, эти субповторы контактируют друг с другом, что придает компактность общей структуре рилина.[37] Последним идёт короткий участок длиной в 32 аминокислоты, богатый основными остатками (англ. C-Terminal Region, CTR; отмечен на схеме знаком «плюс»). Этот участок отличается высокой эволюционной консервативностью: он на 100 % идентичен у всех млекопитающих с установленной структурой гена RELN. Ранее считалось, что этот участок необходим для выделения белка из клетки, так как секреция рилина нарушена у мыши reeler подтипа Orleans, вырабатывающей неполный белок — без части 8-го повтора и без CTR. Было установлено, что секреция нарушается в первую очередь из-за отсечения белка посередине любого из повторов, а чистое отсечение CTR приводит лишь к снижению секреции. [38]
В организме рилин подвергается процессингу и разделяется на три части. Позиции разделения находятся приблизительно между 2 и 3 рилиновым повтором и между повторами 6 и 7 (на схеме отмечены стрелками).[39] Расщепление рилина не снижает его активности, напротив, оно может быть необходимо для правильного кортикогенеза.[40] Конструкции, образованные центральными сегментами белка (повторы 3—6) эффективно связываются с липопротеиновыми рецепторами, вызывают последующее фосфорилирование DAB1 и способствуют развитию кортикальной пластинки так же, как и целый белок.[41]
Эпитопы рилина, взаимодействующие с антителами 142, G-10, CR-50, 12 и 14, также помечены на схеме.
Функции белка и механизм его действия
[править | править код]Наиболее важная предполагаемая роль рилина — участие в регулировании правильного построения слоев мозга, позиционирования клеток и образования связей в пренатальном периоде развития. Однако белок задействован во множестве других, ещё не изученных, процессов.
Функции белка
[править | править код]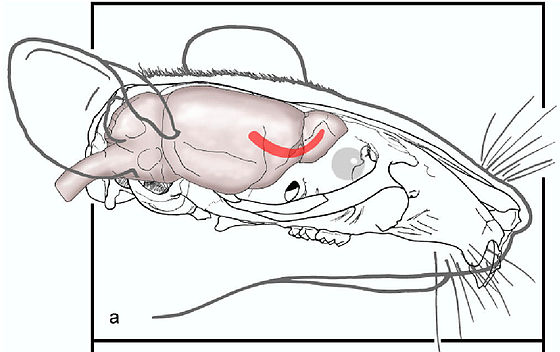
Выявление всех функций белка осложнено предполагаемым многообразием его ролей и широкой распространённостью в организме. Можно условно разделить сферы действия белка по двум показателям — временному (стадия развития организма) и пространственному (локализация в организме).
На ранних периодах развития экспрессия рилина временно обнаруживается во множестве развивающихся органов за пределами центральной нервной системы, исчезая по завершении их формирования. Роль белка в этих процессах изучена недостаточно, так как нарушение выработки рилина у нокаутных мышей-мутантов не приводит к явным патологиям этих органов. Во взрослом организме, присутствие рилина отмечается в меньшем количестве органов, причём всплеск активности белка зачастую наблюдается при повреждении органа.[30][31] Точная функция рилина и в этих случаях продолжает оставаться объектом научных исследований.


Роль рилина в раннем развитии нервной системы более глубоко изучена. Белок способствует дифференциации и ориентации волокон радиальной глии, вдоль которых мигрируют нейробласты.[45] Положение слоя рилин-производящих клеток играет большую роль, так как радиальная глия ориентирует свои волокна в направлении большей концентрации рилина.[44] Второй процесс в развитии мозга, зависящий от наличия рилина — кортикогенез, в частности, расщепление предпластинки (англ. preplate) на маргинальную зону и субпластинку и заселение пространства между ними — кортикальной пластинки — пятью горизонтальными слоями нейронов в «обратном» порядке. Обратный порядок построения слоев кортикальной пластинки, при котором более молодые нейробласты преодолевают ряды уже прижившихся клеток и выстраивают свой слой выше, отличает мозг млекопитающих от более эволюционно древнего мозга пресмыкающихся, в котором слои выстраиваются «снаружи внутрь». При отсутствии рилина, кортикальные слои мыши-мутанта reeler также выстраиваются снаружи внутрь, причём более молодые клетки не могут преодолеть уже созданные кортикальные слои. При этом в пространстве, расположенном под мягкой мозговой оболочкой, образуется так называемая «суперпластинка» — перенаселённый слой, в котором смешаны неправильно расположенные нейроны субпластинки, клетки Кахаля-Ретциуса, и нейроны, которые должны были преодолеть субпластинку и остановиться непосредственно за ней, оставив место вверху для следующих слоев.
Нет единого мнения касательно роли рилина в правильном построении слоев. Изначальное предположение о том, что белок служит стоп-сигналом для мигрирующих клеток, подтверждается его способностью провоцировать разъединение нейронов,[46] его ролью в создании ровного слоя гранулярных клеток в зубчатой извилине гиппокампа, а также тем, что мигрирующие нейробласты избегают внедрения в зоны, насыщенные рилином. Однако данные о том, что нормальный кортикогенез восстанавливается независимо от положения слоя рилин-производящих клеток в опытах на мышах,[47] а также отсутствие экспериментальных доказательств воздействия белка на конусы роста и ведущие отростки нейронов, породили дополнительные гипотезы. По одной из них, рилин повышает чувствительность клеток к ещё неоткрытому позиционному сигналу.
Исследуется роль рилина в развитии спинного мозга: в одном исследовании отмечается, что положение и концентрация рилина влияет на миграцию симпатических преганглионарных нейронов.[48]

Роль рилина в нервной системе взрослого организма связана с двумя наиболее активными участками нейрогенеза во взрослом мозге — субвентрикулярной зоной и зубчатой извилиной. Цепочки нейробластов, осуществляющие в мозге некоторых видов животных тангенциальную миграцию по ростральному миграционному тракту (РМТ) из субвентрикулярной зоны в обонятельную луковицу, под воздействием рилина распадаются на отдельные клетки. Эти клетки приобретают способность преодолевать уже существующие слои нейронов и осуществляют радиальную миграцию вдоль глиальных волокон. Есть данные, что в самом РМТ воздействие на оба рилиновых рецептора осуществляется не рилином,[50] а другим лигандом, вероятно, тромбоспондином 1.[42] В зубчатой извилине рилин отвечает за поддержание компактного слоя гранулярных клеток, постоянно пополняемого новыми нейронами, которые зарождаются в субгранулярной зоне.[51]
Рилин у взрослых также продолжает выделяться ГАМКергическими интернейронами коры мозга, зародившимися в медиальном ганглионарном бугорке. Выделяемый ими рилин усиливает синаптическую пластичность и долговременную потенциацию,[2] взаимодействуя с рецепторами ApoER2 и VLDLR.
По данным французских исследователей,[52] рилин может участвовать в возрастном изменении композиции NMDA-рецептора, повышая мобильность рецепторов, содержащих субъединицу NR2B и тем самым уменьшая время их пребывания в синапсе.[53][54] По их мнению, это способствует «переключению NR2B->NR2A» (англ. NR2B-NR2A switch), отмечаемому[55] при постнатальном развитии мозга. В 2009 году они предположили на основании нового исследования, что и во взрослом гиппокампе постоянная секреция рилина необходима для поддержания на низком уровне числа NR2B-содержащих NMDA-рецепторов.[49]
Эволюционное значение рилина
[править | править код]
Как предполагается, сигнальные взаимодействия рилин-DAB1 сыграли ключевую роль в эволюции архитектуры коры головного мозга, прошедшей путь от однослойного рептилоподобного кортекса у общего эволюционного прародителя амниотов до многослойного кортекса у современных млекопитающих.[56] В исследованиях разных видов показано, что при движении к более сложному кортексу интенсивность экспрессии рилина возрастает, достигая максимума у человека, у которого к тому же отмечено значительное усложнение аксонального сплетения клеток Кахаля-Ретциуса.[57] Рилин присутствует в конечном мозге всех исследованных на данный момент позвоночных, однако характер экспрессии значительно варьирует: у рыб D. rerio не обнаружено клеток Кахаля-Ретциуса и рилин выделяется другими нейронами.[58][59] Выраженного слоя клеток Кахаля-Ретциуса нет и у земноводных, у которых также крайне слабо выражена радиальная миграция.[58]
С усложнением коры и развитием извилин растёт роль миграции нейронов вдоль волокон радиальной глии, и здесь, как считается, важную эволюционную роль сыграло появление отчетливого слоя рилин-производящих клеток.[44] Противоречивые данные о важности этого слоя[47] объясняются некоторыми исследователями либо наличием более тонкого механизма позиционирования, вступающего во взаимодействие с рилиновым каскадом или противодействующего ему,[47] либо избыточностью распределённого производства рилина в мозге мышей[60]в противовес более локализованному синтезу у человека.[22]
В клетках Кахаля-Ретциуса, подавляющая часть которых исчезает к моменту рождения, одновременно с рилином экспрессируется ген HAR1, структура которого у человека наиболее сильно отличается от шимпанзе; это наиболее «эволюционно ускоренный» ген из т. н. зон ускоренного развития у человека (англ. Human Accelerated Regions, HARs).[61] Эволюция сигнального пути рилина продолжается: в исследовании 2007 года было отмечено недавнее эволюционное изменение гена DAB1, распространившееся в китайской популяции, но не затронувшее остальные.[62][63]
Механизм действия
[править | править код]

Доказано, что рилин воздействует на рецепторы VLDLR и ApoER2. N-терминальный участок рилина связывается с альфа-3-бета-1-интегрином.[66] Предполагалось также действие на кадгерин-связанные нейрональные рецепторы (англ. cadherin-related neuronal receptors; CNR-receptors),[67] но последнее было поставлено под сомнение.[41] Внутриклеточные сегменты рецепторов VLDLR и ApoER2 вызывают фосфорилирование адаптерного белка цитоплазмы DAB1 двумя киназами src-семейства, Src[68] и Fyn.[69]
Предположительно, фосфорилированный DAB1 стимулирует перестройку актинового цитоскелета клетки и изменяет насыщенность клеточной поверхности рецепторами альфа-3-бета-1-интегрина, что снижает силу сцепления мигрирующего нейрона с волокнами радиальной глии. В одном исследовании делается вывод, что для правильного построения слоёв важно наличие бета-1 интегриновых рецепторов не на самих перемещающихся нейробластах, а в первую очередь на глиальных клетках.[70] Радиальная глия, по данным одного исследования, содержит столько же рецепторов ApoER2, как и нейроны, но вдесятеро меньшее число рецепторов VLDLR.[45]
Фосфорилирование DAB1 через некоторое время вызывает его убиквитинирование и последующую деградацию,[71] поэтому при недостатке рилина его концентрация растёт; такая отрицательная обратная связь может играть важную роль в построении кортикальных слоёв.[72] Под воздействием двух антител, распознающих основные рецепторы, DAB1 фосфорилируется, но последующего снижения его концентрации и исправления reeler-фенотипа не происходит, что может говорить о передаче части сигнала помимо DAB1.[41]
С внутриклеточным сегментом VLDLR также связывается белок LIS1, известный благодаря своей роли в развитии лиссэнцефалии.[64] Отслеживание путей миграции говорит о том, что VLDLR опосредует выполнение стоп-сигнала, а ApoER2 жизненно необходим для миграции поздно зародившихся нейронов неокортекса.[73]
Вызываемое рилином усиление дендритогенеза опосредовано киназами семейства Src и зависит от экспрессии Crk и CrkL, по результатам одного исследования,[4] согласующегося с ранними сообщениями о взаимодействии этих регуляторов с тирозин-фосфорилированным DAB1.[74] Более того, в одном исследовании с использованием Cre-LoxP-рекомбинации заглушка экспрессии Crk и CrkL в нейронах вызвала у мыши reeler-фенотип, следовательно, эти адаптерные белки расположены между DAB1 и Akt в сигнальной цепочке рилина.[75]
Показано, что рилиновый сигнальный каскад неустановленным пока образом активирует каскад трансмембранного рецептора Notch-1, что приводит к индукции экспрессии липид-связывающего белка мозга FABP7 и переходу нейрональных клеток-предшественников к фенотипу радиальной глии.[65]
Было показано, что молекулы рилина объединяются в олигомеры, необходимые для эффективного фосфорилирования DAB1.[76][77] Более того, два основных рецептора рилина также способны образовывать кластеры,[78] и этот процесс может быть важен для передачи сигнала, так как ведёт к объединению DAB1 в димеры или олигомеры, что активирует цепочку даже в отсутствие рилина.[78]
С другой стороны, рилин одновременно является сериновой протеазой, имея способность разрушать белки расщеплением пептидных связей между составляющими их аминокислотами,[79] что может играть роль в регулировании сцепления и миграции нейронов.
Как показано в одной работе, правильный кортикогенез происходит лишь при разделении рилина на фрагменты, осуществляемое неидентифицированными металлопротеиназами, которые выделяются эмбриональными нейронами[40] и, возможно, с участием ещё менее известных механизмов протеолиза.[80] Как предполагается, полноразмерный рилин цепляется за волокна внеклеточного матрикса, а протеиназы позволяют высвободить важную центральную часть белка.[40] Возможно, центральная часть, проникая в глубинные слои, в большей степени способствует миграции нейробластов, а приближаясь к верхнему слою, клетки прекращают дальнейшую миграцию либо из-за повышенной общей концентрации рилина, либо из-за того, что его полноразмерные молекулы и гомодимеры, закреплённые в матриксе, действуют отлично от центральных фрагментов.[9]
Как и другие белки суперсемейства липопротеиновых рецепторов, VLDLR и ApoER2 содержат в своей структуре так называемые интернализационные домены — мотивы NPxY, позволяющие захватывать лиганды, в том числе и рилин, и осуществлять их эндоцитоз. По данным одного исследования, после эндоцитоза рилина его N-терминальный участок может снова секретироваться клеткой.[81] Этот фрагмент белка, согласно другому исследованию, может препятствовать излишнему разрастанию апикальных дендритов пирамидальных нейронов II\III слоя, активируя сигнальную цепочку, не связанную с основными рецепторами рилина.[82]
По данным одной группы исследователей, активация рилинового каскада приводит к фосфорилированию внутриклеточного белка кофилин-1 в позиции ser3, что может приводить к стабилизации актинового цитоскелета и останавливать рост нейрональных отростков при нейромиграции.[83][84][85]
Усиление долговременной потенциации
[править | править код]Вызываемое рилином усиление долговременной потенциации происходит при взаимодействии ApoER2 с глутаматным NMDA-рецептором. Было показано, что для этого взаимодействия рецептору ApoER2 необходим внутриклеточный домен, кодируемый экзоном 19. Наличие экзона 19 зависит от альтернативного сплайсинга гена; экспрессия ApoER2 с экзоном 19 у мышей возрастает при повышении активности (бодрствование, прием пищи).[86] В одном исследовании показано, что при необходимости формирования памяти экспрессия рилина в гиппокампе быстро возрастает под воздействием деметилаз.[87]
Взаимодействие с Cdk5
[править | править код]Циклин-зависимая киназа 5 (Cdk5), важный регулятор нейромиграции и позиционирования нейронов, взаимодействует с сигнальной цепочкой рилина,[9]:глава 9 фосфорилируя адаптер DAB1.[88][89][90] Она также фосфорилирует некоторые молекулярные мишени, подверженные действию рилина, такие как Тау,[91] активируемый рилином через деактивацию GSK3beta,[92] и Nudel,[93] ассоциированный с Lis1, одной из мишеней DAB1. Усиление LTP, обычно вызываемое рилином в срезах гиппокампа, по данным одного исследования не происходит при нокауте p35, основного активатора Cdk5.[94] Также при двойных нокаутах p35/Dab1, p35/RELN, p35/ApoER2, p35/VLDLR отмечаются усиленные дефекты нейромиграции,[94][95] что говорит о параллельной работе цепочки рилин->ApoER2/VLDLR->DAB1 с цепочкой p35/p39->Cdk5 в правильном построении слоёв при развитии мозга.
Роль в заболеваниях
[править | править код]Выявлены две отдельные мутации гена RELN, приводящие к аутосомно-рецессивной форме лиссенцефалии с гипоплазией мозжечка.[96][97] Мутации нарушают сплайсинг кДНК рилина и вызывают синдром Норман-Робертс. Уровень рилина падает, иногда наличие белка установить не удается. Фенотип пациентов характеризуется гипотонией, атаксией, задержкой физического развития, невозможностью поддерживать равновесие в сидячем положении без посторонней помощи и тяжелой задержкой психического развития с минимальными способностями к вербальному общению либо без них. Также наблюдаются судороги и врожденная лимфедема. В 2007 году описана новая гомозиготная сбалансированная хромосомная транслокация, нарушающая структуру гена и связанная с родственными браками.[98]
Снижение экспрессии рилина и его мРНК в тканях мозга больных шизофренией, достигающее 50 % в некоторых областях и сопровождающееся снижением концентрации фермента глутаматдекарбоксилазы-67 (GAD67), было отмечено впервые в 1998[99] и в 2000 годах.[100] Позже эти данные получили независимое подтверждение в ходе посмертных исследований тканей гиппокампа,[101] базальных ганглиев,[102] мозжечка[103] и других областей головного мозга больных шизофренией.[8][104][105][106] В 2001 году в 14 различных лабораториях было проведено исследование тканей префронтальных областей больных шизофренией, предоставленных Невропатологическим Консорциумом Фонда Стэнли. После многомерного анализа полученных данных снижение уровней мРНК рилина было названо наиболее статистически значимым отклонением.[107]
Предположительной причиной сниженного уровня рилина, согласно эпигенетической гипотезе патофизиологии шизофрении (авт. D.R.Grayson, A.Guidotti, E.Costa),[108][109] считается гиперметилирование промотора гена RELN,[110][111] хотя в двух исследованиях подтвердить гиперметилирование не удалось.[112][113] Гиперметилирование ДНК при шизофрении может быть виновно в усилении симптоматики у 60—70 % больных после приёма метионина.[114][115][116][117] Ингибиторы метилирования, а также ингибиторы дезацетилазы гистонов, например, вальпроевая кислота, увеличивают уровни мРНК рилина,[118][119][120] в то время как L-метионин понижает фенотипическую экспрессию белка.[121] В одном исследовании 2007 года было обнаружено повышенное содержание дезацетилазы гистонов HDAC1 в гиппокампах больных шизофренией.[122] Дезацетилазы гистонов подавляют промоторные комплексы генов. На мышиных моделях было показано, что гиперацетилирование гистонов ведет к деметилированию промоторов как рилина, так и GAD67.[123]
У больных шизофренией также отмечено повышение внутриклеточного уровня метилирующего фермента ДНК-метилтрансферазы 1 в вырабатывающих рилин ГАМКергических интернейронах, не распространяющееся на близлежащие пирамидальные нейроны.[124][125] Степень чрезмерной экспрессии DNMT1 различна в разных слоях коры головного мозга больных и коррелирует со снижением уровней рилина и GAD67.[126] Применение ингибиторов DNMT1 у подопытных животных приводит к росту экспрессии рилина и GAD67,[127] а воздействие как DNMT-, так и HDAC-ингибиторов на нейрональные клетки-предшественники активирует оба гена.[128] По данным одного исследования, концентрация S-аденозил-метионина в префронтальной коре больных шизофренией и биполярным расстройством повышена вдвое. [129] S-аденозил-метионин является донором метильных групп, необходимым для работы ДНК-метилтрансфераз. Таким образом, эпигенетическое воздействие повышенного DNMT1 на уровень рилина очень вероятно, но причины повышения DNMT1 при шизофрении пока неизвестны.
Проводится оценка эпигенетического воздействия нейролептиков: например, в одном исследовании показано,[130] что клозапин и сульпирид, но не галоперидол и оланзапин активируют деметилирование промоторов RELN и GAD67.
Уровни рилина и его изоформ в крови больных шизофренией и другими психотическими заболеваниями также отличаются от нормальных, по данным одного исследования.[131]
Данные о том, что заболевание гриппом во время второго триместра беременности приводит к увеличению риска шизофрении у плода, коррелируют с результатами исследования беременных мышей, зараженных человеческим вирусом гриппа на девятый день беременности[132] либо подвергнутых искусственной активации иммунной системы.[133] Выработка рилина клетками Кахаля-Ретциуса в мозге новорожденных мышат значительно снижается, несмотря на нормальный синтез теми же клетками белка калретинина и фермента nNOS.
Отключение транскрипционного фактора NPAS3, нарушение структуры которого было обнаружено у двух больных шизофренией родственников,[134] и схожего с ним по структуре белка NPAS1 приводит у нокаутных мышей к снижению уровня рилина.[135] Механизм воздействия NPAS1 и NPAS3 на уровень рилина неизвестен. У мыши, нокаутной по связанному с шизофренией гену MTHFR, наблюдается снижение уровня рилина в мозжечке.[136]
Хромосомный регион 7q22, в котором располагается ген RELN, связан с развитием шизофрении.[137] Была обнаружена ассоциация аллельных вариантов гена с показателями памяти, вербальной и визуальной рабочей памяти и исполнительных функций у членов нуклеарных семей больных шизофренией.[137] Ассоциация рабочей памяти с одним из аллелей была впоследствии реплицирована.[138] Одно крупное исследование нескольких популяций говорит об ассоциации однонуклеотидного полиморфизма rs7341475 гена RELN с риском шизофрении у женщин, но не у мужчин.[139] По данным исследования, риск развития болезни у обладательниц этого распространённого полиморфизма повышен в 1.4 раза. В исследовании 25 пациентов с использованием МРТ была отмечена ассоциация полиморфизма Val997Leu гена RELN с увеличением размеров правого и левого желудочков мозга.[140]
По данным одного исследования, у нелеченных пациентов наблюдается сниженная экспрессия рилинового рецептора VLDLR в периферических лимфоцитах.[141] После шести месяцев терапии экспрессия повышается. По словам исследователей, уровень экспрессии VLDLR может служить надежным периферийным биомаркером заболевания.
Учитывая роль белка в дендритогенезе,[3][4] выдвигаются предположения о том, что наблюдаемое при шизофрении значительное локализованное снижение количества дендритных шипиков[142][143] может быть связано с дефицитом рилина.[144][145]
Ген, кодирующий NMDAr-субъединицу NR2B, на мобильность которой влияет рилин в процессе возрастной смены конфигурации NMDA-рецептора,[54] по совокупности генетических исследований[146] является одним из наиболее устойчивых кандидатов в числе возможных генов риска шизофрении. Также RELN и NR2B объединяет то, что оба гена имеют в своей структуре «T-элемент» и оттого подвержены регуляции со стороны транскрипционного фактора TBR1.[147]
У гетерозиготных мышей reeler, гаплонедостаточных по гену RELN, отмечается ряд нейрохимических и поведенческих отклонений, типичных для шизофрении и биполярного расстройства,[6][7] но эти свойства сочтены недостаточными для использования данных мышей в качестве генетической модели шизофрении.[148]
Снижение экспрессии рилина и фермента GAD67 совместно с повышением экспрессии DNMT1, наблюдающееся при шизофрении, характерно лишь для психотической формы биполярного расстройства.[100] Одно исследование позволяет предположить, что при психотическом биполярном расстройстве, в отличие от шизофрении, указанные комплексные нарушения обнаруживаются в коре, но не затрагивают глубинные структуры мозга. В ГАМКергических нейронах базальных ганглиев больных не наблюдалось повышения уровня DNMT1 и сопутствующего эпигенетического подавления экспрессии рилина и GAD67.[102]
При депрессивной непсихотической форме биполярного расстройства снижения уровня рилина не наблюдается, что может говорить о специфической связи нарушения с психозами.
В 2009 году в ходе генетического анализа получено предварительное, требующее репликации, свидетельство в пользу того, что вариация гена RELN (SNP rs362719)[149] ассоциирована с предрасположенностью к биполярному расстройству у женщин.
Височная эпилепсия: дисперсия гранулярных клеток
[править | править код]Характерная патология мозга при височной эпилепсии — дисперсия гранулярных клеток гиппокампа. Дисперсия встречается, по разным оценкам, у 45 %-73 % пациентов. Тяжесть этой патологии напрямую связана с недостатком рилина[150][151][152] и, по данным одного небольшого исследования, коррелирует с гиперметилированием промотора гена RELN.[153] В мышиной модели медиальной височной эпилепсии, длительные судороги приводят к потере рилин-вырабатывающих интернейронов и последующему нарушению позиционирования новорожденных гранулярных клеток зубчатой извилины. Сниженная концентрация рилина не позволяет цепочкам мигрирующих нейробластов вовремя разъединиться и прекратить миграцию.[154] Более того, в одном исследовании с использованием каинатной мышиной модели эпилепсии экзогенная подача рилина в гиппокамп значительно снизила дисперсию гранулярных клеток.[155]
По данным одного исследования, при болезни Альцгеймера изменён характер экспрессии и гликозилирования рилина. В коре головного мозга пациентов уровень рилина повышен на 40 %, в то время как мозжечковый уровень рилина остаётся нормальным.[156] Присутствие рилина до этого было отмечено в бета-амилоидных бляшках у трансгенных мышей, служащих моделью болезни Альцгеймера.[157] Одно генетическое исследование 2008 года говорит об ассоциации гена RELN с болезнью Альцгеймера; воздействие вариантов гена оказалось более выраженным у женщин.[11] При болезни Альцгеймера отмечено значительное сокращение числа клеток Кахаля-Ретциуса в первом слое коры.[158][159] Показано, что рилин взаимодействует с прекурсором бета-амилоида,[160] и, по данным одного исследования in vitro, сдерживает вызываемое бета-амилоидом подавление активности NMDA-рецептора.[161] Некоторые авторы предполагают, что сигнальный путь рилина связывает болезнь Альцгеймера с шизофренией.[162] По данным одного исследования, рилин противодействует снижению активности NMDA-рецептора, вызываемому бета-амилоидом.[163]
Данные о связи рилина с аутизмом противоречивы. В трёх исследованиях подобной связи не выявлено,[164][165][166] в двух исследованиях обнаружено предполагаемое воздействие гена RELN.[167][168] Одно посмертное исследование показало значительное снижение уровня рилина в коре мозжечка у пятерых людей, страдавших аутизмом.[169] В исследовании 2002 года отмечено значительное снижение уровней рилина в крови как у пациентов с аутизмом, так и у их родственников.[170]
В клетках злокачественных опухолей часто нарушаются паттерны метилирования ДНК. По результатам одного исследования, при раке поджелудочной железы наблюдается подавление экспрессии гена RELN и других компонентов сигнального пути белка рилин, а искусственная заглушка сигнального пути рилина в раковых клетках усиливает их подвижность, инвазивность, склонность к формированию колоний.[171] С другой стороны, в раковых клетках простаты экспрессия рилина, наоборот, повышена и коррелирует со степенью агрессивности опухоли по шкале Глисона. [172] Повышена экспрессия рилина и в клетках ретинобластомы.[173]
Другие состояния
[править | править код]В одном исследовании отмечена ассоциация гена RELN с отосклерозом.[174] Согласно статистическому генетическому исследованию 2007 года, экспрессия RELN и DAB1 повышена в мозге мышей, устойчивых к поражению мозга малярией, в отличие от восприимчивых мышей. Авторы исследования предположили, что сигнальная цепочка рилина способна оказывать защитный эффект.[175]
Одна группа исследователей в 2009 году сообщила о том, что при старении у грызунов и приматов в мозге отмечаются амилоидоподобные отложения рилина.[176][177]
Регуляция экспрессии рилина
[править | править код]
Помимо общего числа рилин-производящих клеток, на экспрессию белка влияет множество факторов. На эпигенетическом уровне фактор транскрипции TBR1 регулирует экспрессию RELN и других генов, имеющих в своей структуре «T-элемент».[147] Диета, богатая метионином, может активировать эпигенетические механизмы подавления экспрессии. Исследуется также роль поведения и общения: так, отмечена корреляция материнской заботы (груминга[вд]) у крыс с уровнем рилина как в гиппокампе,[179] так и в коре мозга детёнышей.[178] У крыс, выращенных в условиях социальной изоляции, к 80 дню отмечается сниженный уровень рилина в мозге и нарушенное преимпульсное ингибирование, что может говорить о крайней важности социальных взаимодействий для установления нормальной карты связей в развивающейся префронтальной коре.[180] В одном исследовании отмечено значительное снижение уровня рилина в гиппокампах мышей, длительно получавших кортикостерон:[181] избыточное воздействие глюкокортикоидов на гиппокамп является одним из гипотетических механизмов депрессивных расстройств. В одном небольшом посмертном исследовании отмечено повышенное метилирование гена RELN в коре мозга психически здоровых лиц, прошедших пубертат, по сравнению с теми, кто ещё не вступил в период полового созревания.[182] По данным одного исследования, инъекция трийодтиронина усилила экспрессию рилина и BDNF в гиппокампе крыс.[183]
Психотропные медикаменты
[править | править код]Оценка влияния медикаментов важна, так как уровень рилина обычно измеряется посмертно у пациентов, проходивших длительную терапию. Возлагаются надежды на то, что средства, активирующие деметилирование, такие как клозапин, сульпирид[130] и вальпроевая кислота, будут способны лучше влиять на симптоматику, однако другие исследования говорят о том, что психотропные вещества могут снижать экспрессию белка, и данные по метилированию промотеров в одних публикациях не согласуются с данными экспрессии в других, что говорит о необходимости дальнейшего изучения механизмов, связанных с рилином и GAD67.
Например, Fatemi et al. была проведена оценка воздействия психотропных медикаментов на экспрессию мРНК и белка рилина во фронтальной коре крыс (цитируется по 22й главе сборника, стр. 328[9]):
| Клозапин | Флуоксетин | Галоперидол | Литий | Оланзапин | Вальпроевая кислота | ||
| Reelin: | Protein | ↓ | без изм. | ↓ | ↓ | ↑ | без изм. |
| mRNA | ↑ | ↑ | ↓ | ↑ | ↑ | ↓ |
В 2009 году группа Fatemi et al. опубликовала результаты расширенных исследований, в ходе которых, кроме рилина, анализировалась экспрессия молекул-участников рилиновой цепочки (VLDLR, DAB1, GSK3beta), а также ферментов GAD65 и GAD67 под воздействием тех же медикаментов.[184]
Книги и монографии по теме
[править | править код]- «Гликопротеин рилин: структура, биология, роли в нормальном функционировании и при заболеваниях». Под редакцией Хусейна Фатеми. На английском языке. Reelin Glycoprotein: Structure, Biology and Roles in Health and Disease by Fatemi, S. Hossein (Ed.) 2008, XXIV, 444 p. 111 illus., 36 in color., Hardcover ISBN 978-0-387-76760-4
Обзорные статьи
[править | править код]- Роль процессов метилирования в этиологии и патогенезе шизофрении (недоступная ссылка) А. В. Наумов Ю. Е. Разводовский; Журнал неврологии и психиатрии им. С. С. Корсакова 8/2009
- Forster E, Jossin Y, Zhao S, Chai X, Frotscher M, Goffinet AM. (2006) Recent progress in understanding the role of Reelin in radial neuronal migration, with specific emphasis on the dentate gyrus. Eur J Neurosci. 23(4):901-9. Review. PMID 16519655 (полный текст в открытом доступе (англ.)) — «Недавний прогресс в понимании роли, играемой белком Рилин в радиальной миграции нейронов. Особый разбор роли белка в зубчатой извилине. Обзорная статья.»
- Роль рилина при аутизме, обзор 2009 года: Kelemenova S., Ostatnikova D. Neuroendocrine pathways altered in autism. Special role of reelin (англ.) // Neuro Endocrinol Lett[англ.] : journal. — 2009. — October (vol. 30, no. 4). — PMID 20010491.
Ссылки
[править | править код]- С сайта Schizophrenia Research Forum:
- Are You Reelin in the Years? Not without Alternative Splicing — обзор двух публикаций.[2][86] Перевод: Кто старое помянет? О пользе альтернативного сплайсинга (недоступная ссылка).
- On Again, Off Again—DNA Methylation, Genes, and Plasticity — обзор двух публикаций.[87][123] Перевод: И включаем, и выключаем. Метилирование ДНК, гены и пластичность (недоступная ссылка).
- An Epigenetic Hypothesis for Schizophrenia Pathophysiology (англ.), Dennis R. Grayson, Alessandro Guidotti, Erminio Costa. Перевод: Эпигенетическая гипотеза патофизиологии шизофрении; описывается возможный механизм снижения экспрессии рилина при шизофрении.
- С сайта «Форум исследования болезни Альцгеймера»:
- Reelin, Aβ, α7 Play Yin and Yang Around NMDA Receptors — синопсис трёх исследований, одно (Durakoglugil et al., 2009)[163] связано с рилином.
- Изображения:
- Экспрессия рилина в мозге мыши (англ.) — Brain Gene Expression Map (BGEM)
- Экспрессия рилина в мозге мыши (недоступная ссылка) (англ.) — Allen Brain Atlas.
- Экспрессия рилина в коре мозга человека (недоступная ссылка) (англ.) — проект Gene Expression in the Human Cortex, Институт Аллена.
- Corticogenesis in wild-type, reeler mutant and β1 deficient mice — иллюстрация к статье Brain development: Integrins and the Reelin pathway (недоступная ссылка), показывающая отличия в строении слоев коры мозга у здоровой мыши, у мыши «reeler», и у мыши с недостатком бета-1 интегрина (рилин опосредованно изменяет насыщенность клеточной мембраны интегриновыми рецепторами).
- Reelin Pathway (Cajal-Retzius cells)
- The real role of reelin (англ.) — «Реальная роль белка рилин», публикация в Журнале Клеточной Биологии за 7 октября 2002 года. Перевод: Реальная роль белка рилин.
- Gabriella D’Arcangelo — страница Габриэлы Д`Арканджело, первооткрывателя гена RELN и белка Reelin.
Примечания
[править | править код]- ↑ Weeber E. J., Beffert U., Jones C., Christian J. M., Forster E., Sweatt J. D., Herz J. Reelin and ApoE receptors cooperate to enhance hippocampal synaptic plasticity and learning. (англ.) // The Journal Of Biological Chemistry. — 2002. — 18 October (vol. 277, no. 42). — P. 39944—39952. — doi:10.1074/jbc.M205147200. — PMID 12167620.
- ↑ 1 2 3 D'Arcangelo G. Apoer2: a reelin receptor to remember. (англ.) // Neuron. — 2005. — 18 August (vol. 47, no. 4). — P. 471—473. — doi:10.1016/j.neuron.2005.08.001. — PMID 16102527. Полнотекстовая статья в открытом доступе (недоступная ссылка) (англ.)
- ↑ 1 2 Niu S., Renfro A., Quattrocchi C. C., Sheldon M., D'Arcangelo G. Reelin promotes hippocampal dendrite development through the VLDLR/ApoER2-Dab1 pathway. (англ.) // Neuron. — 2004. — 8 January (vol. 41, no. 1). — P. 71—84. — PMID 14715136.
- ↑ 1 2 3 Matsuki T., Pramatarova A., Howell B.W. Reduction of Crk and CrkL expression blocks reelin-induced dendritogenesis (англ.) // Journal of Cell Science[англ.] : journal. — The Company of Biologists[англ.], 2008. — May. — doi:10.1242/jcs.027334. — PMID 18477607.
- ↑ Niu S., Yabut O., D'Arcangelo G. The Reelin signaling pathway promotes dendritic spine development in hippocampal neurons (англ.) // The Journal of neuroscience : the official journal of the Society for Neuroscience[англ.] : journal. — 2008. — October (vol. 28, no. 41). — P. 10339—10348. — doi:10.1523/JNEUROSCI.1917-08.2008. — PMID 18842893.
- ↑ 1 2 Tueting P., Doueiri M.S., Guidotti A., Davis J.M., Costa E. Reelin down-regulation in mice and psychosis endophenotypes (англ.) // Neurosci Biobehav Rev[англ.] : journal. — 2006. — Vol. 30, no. 8. — P. 1065—1077. — doi:10.1016/j.neubiorev.2006.04.001. — PMID 16769115. Архивировано 28 августа 2017 года.
- ↑ 1 2 Torrey E.F., Barci B.M., Webster M.J., Bartko J.J., Meador-Woodruff J.H., Knable M.B. Neurochemical markers for schizophrenia, bipolar disorder, and major depression in postmortem brains (англ.) // Biol. Psychiatry[англ.] : journal. — 2005. — February (vol. 57, no. 3). — P. 252—260. — doi:10.1016/j.biopsych.2004.10.019. — PMID 15691526. Архивировано 28 августа 2017 года.
- ↑ 1 2 3 4 5 6 7 Reelin Glycoprotein: Structure, Biology and Roles in Health and Disease / под ред. Hossein S. Fatemi. — Springer, 2008. — 444 с. — ISBN 978-0-387-76760-4. Архивировано 6 июня 2011 года.
- ↑ Gene Overview of All Published Schizophrenia-Association Studies for RELN Архивная копия от 21 февраля 2009 на Wayback Machine, «Обзор публикаций по ассоциации гена RELN с шизофренией», база данных Schizophrenia Gene
- ↑ 1 2 Seripa D., Matera M.G., Franceschi M., et al. The RELN locus in Alzheimer's disease (англ.) // J. Alzheimers Dis.[англ.] : journal. — 2008. — July (vol. 14, no. 3). — P. 335—344. — PMID 18599960.
- ↑ FALCONER DS. Two new mutants, 'trembler' and 'reeler', with neurological actions in the house mouse (Mus musculus L.). (англ.) // Journal Of Genetics. — 1951. — January (vol. 50, no. 2). — P. 192—201. — PMID 24539699.
- ↑ HAMBURGH M. ANALYSIS OF THE POSTNATAL DEVELOPMENTAL EFFECTS OF "REELER," A NEUROLOGICAL MUTATION IN MICE. A STUDY IN DEVELOPMENTAL GENETICS. (англ.) // Developmental Biology. — 1963. — October (vol. 8). — P. 165—185. — doi:10.1016/0012-1606(63)90040-x. — PMID 14069672.
- ↑ Caviness Jr. V. S. Patterns of cell and fiber distribution in the neocortex of the reeler mutant mouse. (англ.) // The Journal Of Comparative Neurology. — 1976. — 15 December (vol. 170, no. 4). — P. 435—447. — doi:10.1002/cne.901700404. — PMID 1002868.
- ↑ Miao G.G., Smeyne R.J., D'Arcangelo G., Copeland N.G., Jenkins N.A., Morgan J.I., Curran T. Isolation of an allele of reeler by insertional mutagenesis (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1994. — November (vol. 91, no. 23). — P. 11050—11054. — PMID 7972007. — PMC 45164.
- ↑ D'Arcangelo G., Miao G. G., Chen S. C., Soares H. D., Morgan J. I., Curran T. A protein related to extracellular matrix proteins deleted in the mouse mutant reeler. (англ.) // Nature. — 1995. — 20 April (vol. 374, no. 6524). — P. 719—723. — doi:10.1038/374719a0. — PMID 7715726.
- ↑ Ogawa M., Miyata T., Nakajima K., Yagyu K., Seike M., Ikenaka K., Yamamoto H., Mikoshiba K. The reeler gene-associated antigen on Cajal-Retzius neurons is a crucial molecule for laminar organization of cortical neurons. (англ.) // Neuron. — 1995. — May (vol. 14, no. 5). — P. 899—912. — doi:10.1016/0896-6273(95)90329-1. — PMID 7748558.
- ↑ Trommsdorff M., Gotthardt M., Hiesberger T., Shelton J., Stockinger W., Nimpf J., Hammer R. E., Richardson J. A., Herz J. Reeler/Disabled-like disruption of neuronal migration in knockout mice lacking the VLDL receptor and ApoE receptor 2. (англ.) // Cell. — 1999. — 11 June (vol. 97, no. 6). — P. 689—701. — doi:10.1016/s0092-8674(00)80782-5. — PMID 10380922.
- ↑ Sheldon M., Rice D. S., D'Arcangelo G., Yoneshima H., Nakajima K., Mikoshiba K., Howell B. W., Cooper J. A., Goldowitz D., Curran T. Scrambler and yotari disrupt the disabled gene and produce a reeler-like phenotype in mice. (англ.) // Nature. — 1997. — 16 October (vol. 389, no. 6652). — P. 730—733. — doi:10.1038/39601. — PMID 9338784.
- ↑ Поиск по строке «reelin» в названиях научных статей Архивная копия от 16 октября 2015 на Wayback Machine — Google Scholar
- ↑ Meyer G., Goffinet A. M., Fairén A. What is a Cajal-Retzius cell? A reassessment of a classical cell type based on recent observations in the developing neocortex. (англ.) // Cerebral Cortex (New York, N.Y. : 1991). — 1999. — December (vol. 9, no. 8). — P. 765—775. — doi:10.1093/cercor/9.8.765. — PMID 10600995.
- ↑ 1 2 Meyer G., Goffinet A.M. Prenatal development of reelin-immunoreactive neurons in the human neocortex (англ.) // J. Comp. Neurol.[англ.] : journal. — 1998. — July (vol. 397, no. 1). — P. 29—40. — PMID 9671277.
- ↑ Schiffmann S. N., Bernier B., Goffinet A. M. Reelin mRNA expression during mouse brain development. (англ.) // The European Journal Of Neuroscience. — 1997. — May (vol. 9, no. 5). — P. 1055—1071. — doi:10.1111/j.1460-9568.1997.tb01456.x. — PMID 9182958.
- ↑ Alcántara S., Ruiz M., D'Arcangelo G., Ezan F., de Lecea L., Curran T., Sotelo C., Soriano E. Regional and cellular patterns of reelin mRNA expression in the forebrain of the developing and adult mouse. (англ.) // The Journal Of Neuroscience : The Official Journal Of The Society For Neuroscience. — 1998. — 1 October (vol. 18, no. 19). — P. 7779—7799. — doi:10.1523/JNEUROSCI.18-19-07779.1998. — PMID 9742148.
- ↑ Pesold C., Liu W. S., Guidotti A., Costa E., Caruncho H. J. Cortical bitufted, horizontal, and Martinotti cells preferentially express and secrete reelin into perineuronal nets, nonsynaptically modulating gene expression. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 1999. — 16 March (vol. 96, no. 6). — P. 3217—3222. — doi:10.1073/pnas.96.6.3217. — PMID 10077664.
- ↑ Pesold C., Impagnatiello F., Pisu M. G., Uzunov D. P., Costa E., Guidotti A., Caruncho H. J. Reelin is preferentially expressed in neurons synthesizing gamma-aminobutyric acid in cortex and hippocampus of adult rats. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 1998. — 17 March (vol. 95, no. 6). — P. 3221—3226. — doi:10.1073/pnas.95.6.3221. — PMID 9501244.
- ↑ Suárez-Solá M.L., González-Delgado F.J., Pueyo-Morlans M., Medina-Bolívar O.C., Hernández-Acosta N.C., González-Gómez M., Meyer G. Neurons in the white matter of the adult human neocortex (англ.) // Front Neuroanat : journal. — 2009. — Vol. 3. — P. 7. — doi:10.3389/neuro.05.007.2009. — PMID 19543540. — PMC 2697018.
- ↑ Smalheiser N. R., Costa E., Guidotti A., Impagnatiello F., Auta J., Lacor P., Kriho V., Pappas G. D. Expression of reelin in adult mammalian blood, liver, pituitary pars intermedia, and adrenal chromaffin cells. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 2000. — 1 February (vol. 97, no. 3). — P. 1281—1286. — doi:10.1073/pnas.97.3.1281. — PMID 10655522.
- ↑ Samama B., Boehm N. Reelin immunoreactivity in lymphatics and liver during development and adult life. (англ.) // The Anatomical Record. Part A, Discoveries In Molecular, Cellular, And Evolutionary Biology. — 2005. — July (vol. 285, no. 1). — P. 595—599. — doi:10.1002/ar.a.20202. — PMID 15912522.
- ↑ 1 2 Kobold D., Grundmann A., Piscaglia F., Eisenbach C., Neubauer K., Steffgen J., Ramadori G., Knittel T. Expression of reelin in hepatic stellate cells and during hepatic tissue repair: a novel marker for the differentiation of HSC from other liver myofibroblasts. (англ.) // Journal Of Hepatology. — 2002. — May (vol. 36, no. 5). — P. 607—613. — doi:10.1016/s0168-8278(02)00050-8. — PMID 11983443.
- ↑ 1 2 Pulido J.S., Sugaya I., Comstock J., Sugaya K. Reelin expression is upregulated following ocular tissue injury (англ.) // Graefes Arch. Clin. Exp. Ophthalmol. : journal. — 2007. — June (vol. 245, no. 6). — P. 889—893. — doi:10.1007/s00417-006-0458-4. — PMID 17120005.
- ↑ Buchaille R., Couble M.L., Magloire H., Bleicher F. A substractive PCR-based cDNA library from human odontoblast cells: identification of novel genes expressed in tooth forming cells (англ.) // Matrix biology : journal of the International Society for Matrix Biology : journal. — 2000. — September (vol. 19, no. 5). — P. 421—430. — PMID 10980418. Архивировано 28 августа 2017 года.
- ↑ Allard B., Magloire H., Couble M.L., Maurin J.C., Bleicher F. Voltage-gated sodium channels confer excitability to human odontoblasts: possible role in tooth pain transmission (англ.) // The Journal of biological chemistry : journal. — 2006. — September (vol. 281, no. 39). — P. 29002—29010. — doi:10.1074/jbc.M601020200. — PMID 16831873.
- ↑ Maurin J.C., Couble M.L., Didier-Bazes M., Brisson C., Magloire H., Bleicher F. Expression and localization of reelin in human odontoblasts (англ.) // Matrix biology : journal of the International Society for Matrix Biology : journal. — 2004. — August (vol. 23, no. 5). — P. 277—285. — doi:10.1016/j.matbio.2004.06.005. — PMID 15464360. Архивировано 28 августа 2017 года.
- ↑ Yasui N., Nogi T., Kitao T., Nakano Y., Hattori M., Takagi J. Structure of a receptor-binding fragment of reelin and mutational analysis reveal a recognition mechanism similar to endocytic receptors (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2007. — June (vol. 104, no. 24). — P. 9988—9993. — doi:10.1073/pnas.0700438104. — PMID 17548821. — PMC 1891246.
- ↑ 1 2 Royaux I., Lambert de Rouvroit C., D'Arcangelo G., Demirov D., Goffinet A.M. Genomic organization of the mouse reelin gene (англ.) // Genomics. — Academic Press, 1997. — December (vol. 46, no. 2). — P. 240—250. — PMID 9417911. Архивировано 28 августа 2017 года.
- ↑ Nogi T., Yasui N., Hattori M., Iwasaki K., Takagi J. Structure of a signaling-competent reelin fragment revealed by X-ray crystallography and electron tomography. (англ.) // The EMBO Journal. — 2006. — 9 August (vol. 25, no. 15). — P. 3675—3683. — doi:10.1038/sj.emboj.7601240. — PMID 16858396.
- ↑ Nakano Y., Kohno T., Hibi T., Kohno S., Baba A., Mikoshiba K., Nakajima K., Hattori M. The extremely conserved C-terminal region of Reelin is not necessary for secretion but is required for efficient activation of downstream signaling. (англ.) // The Journal Of Biological Chemistry. — 2007. — 13 July (vol. 282, no. 28). — P. 20544—20552. — doi:10.1074/jbc.M702300200. — PMID 17504759.
- ↑ Lambert de Rouvroit C., de Bergeyck V., Cortvrindt C., Bar I., Eeckhout Y., Goffinet A. M. Reelin, the extracellular matrix protein deficient in reeler mutant mice, is processed by a metalloproteinase. (англ.) // Experimental Neurology. — 1999. — March (vol. 156, no. 1). — P. 214—217. — doi:10.1006/exnr.1998.7007. — PMID 10192793.
- ↑ 1 2 3 Jossin Y., Gui L., Goffinet A.M. Processing of Reelin by embryonic neurons is important for function in tissue but not in dissociated cultured neurons (англ.) // J. Neurosci.[англ.] : journal. — 2007. — April (vol. 27, no. 16). — P. 4243—4252. — doi:10.1523/JNEUROSCI.0023-07.2007. — PMID 17442808.
- ↑ 1 2 3 Jossin Y., Ignatova N., Hiesberger T., Herz J., Lambert de Rouvroit C., Goffinet A.M. The central fragment of Reelin, generated by proteolytic processing in vivo, is critical to its function during cortical plate development (англ.) // J. Neurosci.[англ.] : journal. — 2004. — January (vol. 24, no. 2). — P. 514—521. — doi:10.1523/JNEUROSCI.3408-03.2004. — PMID 14724251. Архивировано 7 октября 2008 года.
- ↑ 1 2 Blake S.M., Strasser V., Andrade N., et al. Thrombospondin-1 binds to ApoER2 and VLDL receptor and functions in postnatal neuronal migration (англ.) // The EMBO journal : journal. — 2008. — October. — doi:10.1038/emboj.2008.223. — PMID 18946489.
- ↑ Lennington J.B., Yang Z., Conover J.C. Neural stem cells and the regulation of adult neurogenesis (англ.) // Reprod. Biol. Endocrinol. : journal. — 2003. — November (vol. 1). — P. 99. — doi:10.1186/1477-7827-1-99. — PMID 14614786. — PMC 293430.
- ↑ 1 2 3 4 5 Nomura T., Takahashi M., Hara Y., Osumi N. Patterns of neurogenesis and amplitude of Reelin expression are essential for making a mammalian-type cortex (англ.) // PLoS ONE : journal. — 2008. — Vol. 3, no. 1. — P. e1454. — doi:10.1371/journal.pone.0001454. — PMID 18197264. — PMC 2175532. Архивировано 16 октября 2008 года.
- ↑ 1 2 Hartfuss E., Förster E., Bock H.H., et al. Reelin signaling directly affects radial glia morphology and biochemical maturation (англ.) // Development : journal. — 2003. — October (vol. 130, no. 19). — P. 4597—4609. — doi:10.1242/dev.00654. — PMID 12925587.
- ↑ Hack I., Bancila M., Loulier K., Carroll P., Cremer H. Reelin is a detachment signal in tangential chain-migration during postnatal neurogenesis (англ.) // Nat. Neurosci. : journal. — 2002. — October (vol. 5, no. 10). — P. 939—945. — doi:10.1038/nn923. — PMID 12244323.
- ↑ 1 2 3 Yoshida M., Assimacopoulos S., Jones K.R., Grove E.A. Massive loss of Cajal-Retzius cells does not disrupt neocortical layer order (неопр.) // Development. — 2006. — February (т. 133, № 3). — С. 537—545. — doi:10.1242/dev.02209. — PMID 16410414.
- ↑ Yip Y.P., Mehta N., Magdaleno S., Curran T., Yip J.W. Ectopic expression of reelin alters migration of sympathetic preganglionic neurons in the spinal cord (англ.) // J. Comp. Neurol.[англ.] : journal. — 2009. — March (vol. 515, no. 2). — P. 260—268. — doi:10.1002/cne.22044. — PMID 19412957.
- ↑ 1 2 Campo C.G., Sinagra M., Verrier D., Manzoni O.J., Chavis P. Reelin secreted by GABAergic neurons regulates glutamate receptor homeostasis (англ.) // PLoS ONE : journal. — 2009. — Vol. 4, no. 5. — P. e5505. — doi:10.1371/journal.pone.0005505. — PMID 19430527.
- ↑ Andrade N., Komnenovic V., Blake S.M., et al. ApoER2/VLDL receptor and Dab1 in the rostral migratory stream function in postnatal neuronal migration independently of Reelin (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2007. — May (vol. 104, no. 20). — P. 8508—8513. — doi:10.1073/pnas.0611391104. — PMID 17494763. — PMC 1895980.
- ↑ Frotscher M., Haas C. A., Förster E. Reelin controls granule cell migration in the dentate gyrus by acting on the radial glial scaffold. (англ.) // Cerebral Cortex (New York, N.Y. : 1991). — 2003. — June (vol. 13, no. 6). — P. 634—640. — doi:10.1093/cercor/13.6.634. — PMID 12764039.
- ↑ INSERM — Olivier Manzoni — Physiopathology of Synaptic Transmission and Plasticity Архивная копия от 25 ноября 2006 на Wayback Machine — группа Оливьера Манцони, нейронаучный институт Бордо.
- ↑ Sinagra M., Verrier D., Frankova D., Korwek K.M., Blahos J., Weeber E.J., Manzoni O.J., Chavis P. Reelin, very-low-density lipoprotein receptor, and apolipoprotein E receptor 2 control somatic NMDA receptor composition during hippocampal maturation in vitro (англ.) // J. Neurosci.[англ.] : journal. — 2005. — June (vol. 25, no. 26). — P. 6127—6136. — doi:10.1523/JNEUROSCI.1757-05.2005. — PMID 15987942.
- ↑ 1 2 Groc L., Choquet D., Stephenson F.A., Verrier D., Manzoni O.J., Chavis P. NMDA receptor surface trafficking and synaptic subunit composition are developmentally regulated by the extracellular matrix protein Reelin (англ.) // J. Neurosci.[англ.] : journal. — 2007. — Vol. 27, no. 38. — P. 10165—10175. — doi:10.1523/JNEUROSCI.1772-07.2007. — PMID 17881522.
- ↑ Liu X.B., Murray K.D., Jones E.G. Switching of NMDA receptor 2A and 2B subunits at thalamic and cortical synapses during early postnatal development (англ.) // J. Neurosci.[англ.] : journal. — 2004. — October (vol. 24, no. 40). — P. 8885—8895. — doi:10.1523/JNEUROSCI.2476-04.2004. — PMID 15470155.
- ↑ Bar I., Lambert de Rouvroit C., Goffinet A.M. The evolution of cortical development. An hypothesis based on the role of the Reelin signaling pathway (англ.) // Trends Neurosci.[англ.] : journal. — 2000. — December (vol. 23, no. 12). — P. 633—638. — PMID 11137154. Архивировано 28 августа 2017 года.
- ↑ Molnár Z., Métin C., Stoykova A., et al. [www.blackwell-synergy.com/openurl?genre=article&sid=nlm:pubmed&issn=0953-816X&date=2006&volume=23&issue=4&spage=921 Comparative aspects of cerebral cortical development] (англ.) // Eur. J. Neurosci.[англ.] : journal. — 2006. — February (vol. 23, no. 4). — P. 921—934. — doi:10.1111/j.1460-9568.2006.04611.x. — PMID 16519657. — PMC 1931431. (недоступная ссылка)
- ↑ 1 2 Pérez-García C.G., González-Delgado F.J., Suárez-Solá M.L., et al. Reelin-immunoreactive neurons in the adult vertebrate pallium (неопр.) // J. Chem. Neuroanat.. — 2001. — January (т. 21, № 1). — С. 41—51. — PMID 11173219. Архивировано 28 августа 2017 года.
- ↑ Costagli A., Kapsimali M., Wilson S.W., Mione M. Conserved and divergent patterns of Reelin expression in the zebrafish central nervous system (англ.) // J. Comp. Neurol.[англ.] : journal. — 2002. — August (vol. 450, no. 1). — P. 73—93. — doi:10.1002/cne.10292. — PMID 12124768.
- ↑ Goffinet A.M. What makes us human? A biased view from the perspective of comparative embryology and mouse genetics (англ.) // J Biomed Discov Collab : journal. — 2006. — Vol. 1. — P. 16. — doi:10.1186/1747-5333-1-13. — PMID 17132178. — PMC 1769396.
- ↑ Pollard K.S., Salama S.R., Lambert N., et al. An RNA gene expressed during cortical development evolved rapidly in humans (англ.) // Nature : journal. — 2006. — September (vol. 443, no. 7108). — P. 167—172. — doi:10.1038/nature05113. — PMID 16915236.
- ↑ Williamson S.H., Hubisz M.J., Clark A.G., Payseur B.A., Bustamante C.D., Nielsen R. Localizing recent adaptive evolution in the human genome (англ.) // PLoS Genet.[англ.] : journal. — 2007. — June (vol. 3, no. 6). — P. e90. — doi:10.1371/journal.pgen.0030090. — PMID 17542651. — PMC 1885279. Архивировано 7 сентября 2008 года.
- ↑ Humans Have Spread Globally, and Evolved Locally (англ.) «Люди распространились глобально и эволюционировали локально» — The New York Times, 26 June 2007
- ↑ 1 2 Zhang G., Assadi A. H., McNeil R. S., Beffert U., Wynshaw-Boris A., Herz J., Clark G. D., D'Arcangelo G. The Pafah1b complex interacts with the reelin receptor VLDLR. (англ.) // PloS One. — 2007. — 28 February (vol. 2, no. 2). — P. e252—252. — doi:10.1371/journal.pone.0000252. — PMID 17330141.
- ↑ 1 2 Keilani S., Sugaya K. Reelin induces a radial glial phenotype in human neural progenitor cells by activation of Notch-1 (англ.) // BMC Dev. Biol. : journal. — 2008. — July (vol. 8, no. 1). — P. 69. — doi:10.1186/1471-213X-8-69. — PMID 18593473.
- ↑ Schmid R.S., Jo R., Shelton S., Kreidberg J.A., Anton E.S. Reelin, integrin and DAB1 interactions during embryonic cerebral cortical development (англ.) // Cereb. Cortex[англ.] : journal. — 2005. — October (vol. 15, no. 10). — P. 1632—1636. — doi:10.1093/cercor/bhi041. — PMID 15703255.
- ↑ Senzaki K., Ogawa M., Yagi T. Proteins of the CNR family are multiple receptors for Reelin (англ.) // Cell : journal. — Cell Press, 1999. — December (vol. 99, no. 6). — P. 635—647. — PMID 10612399. Архивировано 28 августа 2017 года.
- ↑ Howell B.W., Gertler F.B., Cooper J.A. Mouse disabled (mDab1): a Src binding protein implicated in neuronal development (англ.) // EMBO J. : journal. — 1997. — January (vol. 16, no. 1). — P. 121—132. — doi:10.1093/emboj/16.1.121. — PMID 9009273. — PMC 1169619.
- ↑ Arnaud L., Ballif B.A., Förster E., Cooper J.A. Fyn tyrosine kinase is a critical regulator of disabled-1 during brain development (англ.) // Curr. Biol. : journal. — 2003. — January (vol. 13, no. 1). — P. 9—17. — PMID 12526739. Архивировано 28 августа 2017 года.
- ↑ Belvindrah R., Graus-Porta D., Goebbels S., Nave K.A., Müller U. Beta1 integrins in radial glia but not in migrating neurons are essential for the formation of cell layers in the cerebral cortex (англ.) // J. Neurosci.[англ.] : journal. — 2007. — December (vol. 27, no. 50). — P. 13854—13865. — doi:10.1523/JNEUROSCI.4494-07.2007. — PMID 18077697.
- ↑ Feng L., Allen N.S., Simo S., Cooper J.A. Cullin 5 regulates Dab1 protein levels and neuron positioning during cortical development (англ.) // Genes Dev. : journal. — 2007. — November (vol. 21, no. 21). — P. 2717—2730. — doi:10.1101/gad.1604207. — PMID 17974915. — PMC 2045127.
- ↑ Kerjan G., Gleeson J. G. A missed exit: Reelin sets in motion Dab1 polyubiquitination to put the break on neuronal migration. (англ.) // Genes & Development. — 2007. — 15 November (vol. 21, no. 22). — P. 2850—2854. — doi:10.1101/gad.1622907. — PMID 18006681.
- ↑ Hack I., Hellwig S., Junghans D., Brunne B., Bock H.H., Zhao S., Frotscher M. Divergent roles of ApoER2 and Vldlr in the migration of cortical neurons (англ.) // Development : journal. — 2007. — Vol. 134, no. 21. — P. 3883—3891. — doi:10.1242/dev.005447. — PMID 17913789.
- ↑ Ballif B.A., Arnaud L., Arthur W.T., Guris D., Imamoto A., Cooper J.A. Activation of a Dab1/CrkL/C3G/Rap1 pathway in Reelin-stimulated neurons (англ.) // Curr. Biol. : journal. — 2004. — April (vol. 14, no. 7). — P. 606—610. — doi:10.1016/j.cub.2004.03.038. — PMID 15062102.
- ↑ Park T.J., Curran T. Crk and crk-like play essential overlapping roles downstream of disabled-1 in the reelin pathway (англ.) // J. Neurosci.[англ.] : journal. — 2008. — December (vol. 28, no. 50). — P. 13551—13562. — doi:10.1523/JNEUROSCI.4323-08.2008. — PMID 19074029.
- ↑ Utsunomiya-Tate N., Kubo K., Tate S., Kainosho M., Katayama E., Nakajima K., Mikoshiba K. Reelin molecules assemble together to form a large protein complex, which is inhibited by the function-blocking CR-50 antibody. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 2000. — 15 August (vol. 97, no. 17). — P. 9729—9734. — doi:10.1073/pnas.160272497. — PMID 10920200.
- ↑ Kubo K., Mikoshiba K., Nakajima K. Secreted Reelin molecules form homodimers. (англ.) // Neuroscience Research. — 2002. — August (vol. 43, no. 4). — P. 381—388. — doi:10.1016/s0168-0102(02)00068-8. — PMID 12135781.
- ↑ 1 2 Strasser V., Fasching D., Hauser C., et al. Receptor clustering is involved in Reelin signaling (англ.) // Molecular and cellular biology : journal. — 2004. — February (vol. 24, no. 3). — P. 1378—1386. — PMID 14729980. — PMC 321426.
- ↑ Quattrocchi C. C., Wannenes F., Persico A. M., Ciafré S. A., D'Arcangelo G., Farace M. G., Keller F. Reelin is a serine protease of the extracellular matrix. (англ.) // The Journal Of Biological Chemistry. — 2002. — 4 January (vol. 277, no. 1). — P. 303—309. — doi:10.1074/jbc.M106996200. — PMID 11689558.
- ↑ Lugli G., Krueger J.M., Davis J.M., Persico A.M., Keller F., Smalheiser N.R. Methodological factors influencing measurement and processing of plasma reelin in humans (англ.) // BMC Biochem. : journal. — 2003. — September (vol. 4). — P. 9. — doi:10.1186/1471-2091-4-9. — PMID 12959647. — PMC 200967. Архивировано 25 июня 2006 года.
- ↑ Hibi T., Hattori M. The N-terminal fragment of Reelin is generated after endocytosis and released through the pathway regulated by Rab11 (англ.) // FEBS Lett.[англ.] : journal. — 2009. — March. — doi:10.1016/j.febslet.2009.03.024. — PMID 19303411. Архивировано 28 августа 2017 года.
- ↑ Chameau P., Inta D., Vitalis T., Monyer H., Wadman W.J., van Hooft J.A. The N-terminal region of reelin regulates postnatal dendritic maturation of cortical pyramidal neurons (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2009. — April. — doi:10.1073/pnas.0810764106. — PMID 19366679.
- ↑ Chai X., Förster E., Zhao S., Bock H.H., Frotscher M. Reelin stabilizes the actin cytoskeleton of neuronal processes by inducing n-cofilin phosphorylation at serine3 (англ.) // J. Neurosci.[англ.] : journal. — 2009. — January (vol. 29, no. 1). — P. 288—299. — doi:10.1523/JNEUROSCI.2934-08.2009. — PMID 19129405.
- ↑ Frotscher M., Chai X., Bock H.H., Haas C.A., Förster E., Zhao S. Role of Reelin in the development and maintenance of cortical lamination (англ.) // J Neural Transm[англ.] : journal. — 2009. — April. — doi:10.1007/s00702-009-0228-7. — PMID 19396394.
- ↑ Chai X., Förster E., Zhao S., Bock H. H., Frotscher M. Reelin acts as a stop signal for radially migrating neurons by inducing phosphorylation of n-cofilin at the leading edge. (англ.) // Communicative & Integrative Biology. — 2009. — July (vol. 2, no. 4). — P. 375—377. — doi:10.4161/cib.2.4.8614. — PMID 19721896.
- ↑ 1 2 Beffert U., Weeber E. J., Durudas A., Qiu S., Masiulis I., Sweatt J. D., Li W. P., Adelmann G., Frotscher M., Hammer R. E., Herz J. Modulation of synaptic plasticity and memory by Reelin involves differential splicing of the lipoprotein receptor Apoer2. (англ.) // Neuron. — 2005. — 18 August (vol. 47, no. 4). — P. 567—579. — doi:10.1016/j.neuron.2005.07.007. — PMID 16102539.
- ↑ 1 2 Miller C.A., Sweatt J.D. Covalent modification of DNA regulates memory formation (англ.) // Neuron : journal. — Cell Press, 2007. — March (vol. 53, no. 6). — P. 857—869. — doi:10.1016/j.neuron.2007.02.022. — PMID 17359920. Архивировано 7 июня 2008 года.
- ↑ Arnaud L., Ballif B.A., Cooper J.A. Regulation of protein tyrosine kinase signaling by substrate degradation during brain development (англ.) // Mol. Cell. Biol. : journal. — 2003. — December (vol. 23, no. 24). — P. 9293—9302. — PMID 14645539. — PMC 309695.
- ↑ Ohshima T., Suzuki H., Morimura T., Ogawa M., Mikoshiba K. Modulation of Reelin signaling by Cyclin-dependent kinase 5 (англ.) // Brain Res.[англ.] : journal. — 2007. — April (vol. 1140). — P. 84—95. — doi:10.1016/j.brainres.2006.01.121. — PMID 16529723. Архивировано 28 августа 2017 года.
- ↑ Keshvara L., Magdaleno S., Benhayon D., Curran T. Cyclin-dependent kinase 5 phosphorylates disabled 1 independently of Reelin signaling (англ.) // J. Neurosci.[англ.] : journal. — 2002. — June (vol. 22, no. 12). — P. 4869—4877. — PMID 12077184.
- ↑ Kobayashi S., Ishiguro K., Omori A., Takamatsu M., Arioka M., Imahori K., Uchida T. A cdc2-related kinase PSSALRE/cdk5 is homologous with the 30 kDa subunit of tau protein kinase II, a proline-directed protein kinase associated with microtubule (англ.) // FEBS Lett.[англ.] : journal. — 1993. — December (vol. 335, no. 2). — P. 171—175. — PMID 8253190. Архивировано 28 августа 2017 года.
- ↑ Beffert U., Morfini G., Bock H.H., Reyna H., Brady S.T., Herz J. Reelin-mediated signaling locally regulates protein kinase B/Akt and glycogen synthase kinase 3beta (англ.) // J. Biol. Chem. : journal. — 2002. — December (vol. 277, no. 51). — P. 49958—49964. — doi:10.1074/jbc.M209205200. — PMID 12376533.
- ↑ Sasaki S., Shionoya A., Ishida M., Gambello M.J., Yingling J., Wynshaw-Boris A., Hirotsune S. A LIS1/NUDEL/cytoplasmic dynein heavy chain complex in the developing and adult nervous system (англ.) // Neuron : journal. — Cell Press, 2000. — December (vol. 28, no. 3). — P. 681—696. — PMID 11163259. Архивировано 28 августа 2017 года.
- ↑ 1 2 Beffert U., Weeber E.J., Morfini G., Ko J., Brady S.T., Tsai L.H., Sweatt J.D., Herz J. Reelin and cyclin-dependent kinase 5-dependent signals cooperate in regulating neuronal migration and synaptic transmission (англ.) // J. Neurosci.[англ.] : journal. — 2004. — February (vol. 24, no. 8). — P. 1897—1906. — doi:10.1523/JNEUROSCI.4084-03.2004. — PMID 14985430.
- ↑ Ohshima T., Ogawa M., Veeranna, Hirasawa M., Longenecker G., Ishiguro K., Pant H.C., Brady R.O., Kulkarni A.B., Mikoshiba K. Synergistic contributions of cyclin-dependant kinase 5/p35 and Reelin/Dab1 to the positioning of cortical neurons in the developing mouse brain (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2001. — February (vol. 98, no. 5). — P. 2764—2769. — doi:10.1073/pnas.051628498. — PMID 11226314. — PMC 30213.
- ↑ Hong S. E., Shugart Y. Y., Huang D. T., Shahwan S. A., Grant P. E., Hourihane J. O., Martin N. D., Walsh C. A. Autosomal recessive lissencephaly with cerebellar hypoplasia is associated with human RELN mutations. (англ.) // Nature genetics. — 2000. — Vol. 26, no. 1. — P. 93—96. — doi:10.1038/79246. — PMID 10973257.
- ↑ Crino P. New RELN Mutation Associated with Lissencephaly and Epilepsy. (англ.) // Epilepsy Currents. — 2001. — November (vol. 1, no. 2). — P. 72—72. — doi:10.1046/j.1535-7597.2001.00017.x. — PMID 15309195.
- ↑ Zaki M., Shehab M., El-Aleem A.A., et al. Identification of a novel recessive RELN mutation using a homozygous balanced reciprocal translocation (англ.) // Am. J. Med. Genet. A[англ.] : journal. — 2007. — May (vol. 143A, no. 9). — P. 939—944. — doi:10.1002/ajmg.a.31667. — PMID 17431900.
- ↑ Impagnatiello F., Guidotti A. R., Pesold C., Dwivedi Y., Caruncho H., Pisu M. G., Uzunov D. P., Smalheiser N. R., Davis J. M., Pandey G. N., Pappas G. D., Tueting P., Sharma R. P., Costa E. A decrease of reelin expression as a putative vulnerability factor in schizophrenia. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 1998. — 22 December (vol. 95, no. 26). — P. 15718—15723. — doi:10.1073/pnas.95.26.15718. — PMID 9861036.
- ↑ 1 2 Guidotti A., Auta J., Davis J. M., Di-Giorgi-Gerevini V., Dwivedi Y., Grayson D. R., Impagnatiello F., Pandey G., Pesold C., Sharma R., Uzunov D., Costa E. Decrease in reelin and glutamic acid decarboxylase67 (GAD67) expression in schizophrenia and bipolar disorder: a postmortem brain study. (англ.) // Archives Of General Psychiatry. — 2000. — November (vol. 57, no. 11). — P. 1061—1069. — doi:10.1001/archpsyc.57.11.1061. — PMID 11074872.
- ↑ Fatemi S. H., Earle J. A., McMenomy T. Reduction in Reelin immunoreactivity in hippocampus of subjects with schizophrenia, bipolar disorder and major depression. (англ.) // Molecular Psychiatry. — 2000. — November (vol. 5, no. 6). — P. 654—663. — PMID 11126396.
- ↑ 1 2 Veldic M., Kadriu B., Maloku E., Agis-Balboa R. C., Guidotti A., Davis J. M., Costa E. Epigenetic mechanisms expressed in basal ganglia GABAergic neurons differentiate schizophrenia from bipolar disorder. (англ.) // Schizophrenia Research. — 2007. — March (vol. 91, no. 1-3). — P. 51—61. — doi:10.1016/j.schres.2006.11.029. — PMID 17270400.
- ↑ Fatemi S.H., Hossein Fatemi S., Stary J.M., Earle J.A., Araghi-Niknam M., Eagan E. GABAergic dysfunction in schizophrenia and mood disorders as reflected by decreased levels of glutamic acid decarboxylase 65 and 67 kDa and Reelin proteins in cerebellum (англ.) // Schizophrenia Research[англ.] : journal. — Elsevier, 2005. — January (vol. 72, no. 2—3). — P. 109—122. — doi:10.1016/j.schres.2004.02.017. — PMID 15560956. Архивировано 28 августа 2017 года.
- ↑ Eastwood S. L., Harrison P. J. Interstitial white matter neurons express less reelin and are abnormally distributed in schizophrenia: towards an integration of molecular and morphologic aspects of the neurodevelopmental hypothesis. (англ.) // Molecular Psychiatry. — 2003. — September (vol. 8, no. 9). — P. 769—821. — doi:10.1038/sj.mp.4001371. — PMID 12931209.
- ↑ Abdolmaleky H. M., Cheng K. H., Russo A., Smith C. L., Faraone S. V., Wilcox M., Shafa R., Glatt S. J., Nguyen G., Ponte J. F., Thiagalingam S., Tsuang M. T. Hypermethylation of the reelin (RELN) promoter in the brain of schizophrenic patients: a preliminary report. (англ.) // American Journal Of Medical Genetics. Part B, Neuropsychiatric Genetics : The Official Publication Of The International Society Of Psychiatric Genetics. — 2005. — 5 April (vol. 134B, no. 1). — P. 60—66. — doi:10.1002/ajmg.b.30140. — PMID 15717292.
- ↑ Eastwood S. L., Harrison P. J. Cellular basis of reduced cortical reelin expression in schizophrenia. (англ.) // The American Journal Of Psychiatry. — 2006. — March (vol. 163, no. 3). — P. 540—542. — doi:10.1176/appi.ajp.163.3.540. — PMID 16513881.
- ↑ Knable M. B., Torrey E. F., Webster M. J., Bartko J. J. Multivariate analysis of prefrontal cortical data from the Stanley Foundation Neuropathology Consortium. (англ.) // Brain Research Bulletin. — 2001. — 15 July (vol. 55, no. 5). — P. 651—659. — PMID 11576762.
- ↑ An Epigenetic Hypothesis for Schizophrenia Pathophysiology Архивная копия от 17 сентября 2008 на Wayback Machine (англ.), Dennis R. Grayson, Alessandro Guidotti, Erminio Costa. Перевод: Эпигенетическая гипотеза патофизиологии шизофрении Архивная копия от 16 января 2009 на Wayback Machine
- ↑ Grayson D.R., Chen Y., Dong E., Kundakovic M., Guidotti A. From trans-methylation to cytosine methylation: Evolution of the methylation hypothesis of schizophrenia (англ.) // Epigenetics : journal. — 2009. — April (vol. 4, no. 3). — PMID 19395859.
- ↑ Grayson D. R., Jia X., Chen Y., Sharma R. P., Mitchell C. P., Guidotti A., Costa E. Reelin promoter hypermethylation in schizophrenia. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 2005. — 28 June (vol. 102, no. 26). — P. 9341—9346. — doi:10.1073/pnas.0503736102. — PMID 15961543.
- ↑ Dong E., Agis-Balboa R. C., Simonini M. V., Grayson D. R., Costa E., Guidotti A. Reelin and glutamic acid decarboxylase67 promoter remodeling in an epigenetic methionine-induced mouse model of schizophrenia. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 2005. — 30 August (vol. 102, no. 35). — P. 12578—12583. — doi:10.1073/pnas.0505394102. — PMID 16113080.
- ↑ Tochigi M., Iwamoto K., Bundo M., Komori A., Sasaki T., Kato N., Kato T. Methylation Status of the Reelin Promoter Region in the Brain of Schizophrenic Patients (англ.) : journal. — 2007. — doi:10.1016/j.biopsych.2007.07.003. — PMID 17870056.
- ↑ Mill J., Tang T., Kaminsky Z., Khare T., Yazdanpanah S., Bouchard L., Jia P., Assadzadeh A., Flanagan J., Schumacher A., Wang S.C., Petronis A. Epigenomic profiling reveals DNA-methylation changes associated with major psychosis (англ.) // Am. J. Hum. Genet.[англ.] : journal. — 2008. — Vol. 82, no. 3. — P. 696—711. — doi:10.1016/j.ajhg.2008.01.008. — PMID 18319075.
- ↑ Pollin, W., Cardon, P.V., and Kety, S.S. (1961) Effects of amino acid feedings in schizophrenia patients treated with iproniazid. Science 133 , 104—105.
- ↑ BRUNE GG, HIMWICH HE. Effects of methionine loading on the behavior of schizophrenic patients. (англ.) // The Journal Of Nervous And Mental Disease. — 1962. — May (vol. 134). — P. 447—450. — PMID 13873983.
- ↑ PARK LC, BALDESSARINI RJ, KETY SS. METHIONINE EFFECTS ON CHRONIC SCHIZOPHRENICS: PATIENTS TREATED WITH MONOAMINE OXIDASE INHIBITORS. (англ.) // Archives Of General Psychiatry. — 1965. — April (vol. 12). — P. 346—351. — doi:10.1001/archpsyc.1965.01720340018003. — PMID 14258360.
- ↑ Antun F. T., Burnett G. B., Cooper A. J., Daly R. J., Smythies J. R., Zealley A. K. The effects of L-methionine (without MAOI) in schizophrenia. (англ.) // Journal Of Psychiatric Research. — 1971. — June (vol. 8, no. 2). — P. 63—71. — PMID 4932991.
- ↑ Tremolizzo L., Doueiri M. S., Dong E., Grayson D. R., Davis J., Pinna G., Tueting P., Rodriguez-Menendez V., Costa E., Guidotti A. Valproate corrects the schizophrenia-like epigenetic behavioral modifications induced by methionine in mice. (англ.) // Biological Psychiatry. — 2005. — 1 March (vol. 57, no. 5). — P. 500—509. — doi:10.1016/j.biopsych.2004.11.046. — PMID 15737665.
- ↑ Chen Y., Sharma R. P., Costa R. H., Costa E., Grayson D. R. On the epigenetic regulation of the human reelin promoter. (англ.) // Nucleic Acids Research. — 2002. — 1 July (vol. 30, no. 13). — P. 2930—2939. — doi:10.1093/nar/gkf401. — PMID 12087179.
- ↑ Mitchell C. P., Chen Y., Kundakovic M., Costa E., Grayson D. R. Histone deacetylase inhibitors decrease reelin promoter methylation in vitro. (англ.) // Journal Of Neurochemistry. — 2005. — April (vol. 93, no. 2). — P. 483—492. — doi:10.1111/j.1471-4159.2005.03040.x. — PMID 15816871.
- ↑ Tremolizzo L., Carboni G., Ruzicka W. B., Mitchell C. P., Sugaya I., Tueting P., Sharma R., Grayson D. R., Costa E., Guidotti A. An epigenetic mouse model for molecular and behavioral neuropathologies related to schizophrenia vulnerability. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 2002. — 24 December (vol. 99, no. 26). — P. 17095—17100. — doi:10.1073/pnas.262658999. — PMID 12481028.
- ↑ Benes F.M., Lim B., Matzilevich D., Walsh J.P., Subburaju S., Minns M. Regulation of the GABA cell phenotype in hippocampus of schizophrenics and bipolars (англ.) : journal. — 2007. — doi:10.1073/pnas.0703806104. — PMID 17553960. free full text PDF Архивная копия от 30 сентября 2007 на Wayback Machine
- ↑ 1 2 Dong E., Guidotti A., Grayson D. R., Costa E. Histone hyperacetylation induces demethylation of reelin and 67-kDa glutamic acid decarboxylase promoters. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 2007. — 13 March (vol. 104, no. 11). — P. 4676—4681. — doi:10.1073/pnas.0700529104. — PMID 17360583.
- ↑ Veldic M., Caruncho H. J., Liu W. S., Davis J., Satta R., Grayson D. R., Guidotti A., Costa E. DNA-methyltransferase 1 mRNA is selectively overexpressed in telencephalic GABAergic interneurons of schizophrenia brains. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 2004. — 6 January (vol. 101, no. 1). — P. 348—353. — doi:10.1073/pnas.2637013100. — PMID 14684836.
- ↑ Veldic M., Guidotti A., Maloku E., Davis J. M., Costa E. In psychosis, cortical interneurons overexpress DNA-methyltransferase 1. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 2005. — 8 February (vol. 102, no. 6). — P. 2152—2157. — doi:10.1073/pnas.0409665102. — PMID 15684088.
- ↑ Ruzicka W. B., Zhubi A., Veldic M., Grayson D. R., Costa E., Guidotti A. Selective epigenetic alteration of layer I GABAergic neurons isolated from prefrontal cortex of schizophrenia patients using laser-assisted microdissection. (англ.) // Molecular Psychiatry. — 2007. — April (vol. 12, no. 4). — P. 385—397. — doi:10.1038/sj.mp.4001954. — PMID 17264840.
- ↑ Kundakovic M., Chen Y., Costa E., Grayson D. R. DNA methyltransferase inhibitors coordinately induce expression of the human reelin and glutamic acid decarboxylase 67 genes. (англ.) // Molecular Pharmacology. — 2007. — March (vol. 71, no. 3). — P. 644—653. — doi:10.1124/mol.106.030635. — PMID 17065238.
- ↑ Kundakovic M., Chen Y., Guidotti A., Grayson D.R. The reelin and GAD67 promoters are activated by epigenetic drugs that facilitate the disruption of local repressor complexes (англ.) // Mol. Pharmacol.[англ.] : journal. — 2008. — November. — doi:10.1124/mol.108.051763. — PMID 19029285.
- ↑ Guidotti A., Ruzicka W., Grayson D. R., Veldic M., Pinna G., Davis J. M., Costa E. S-adenosyl methionine and DNA methyltransferase-1 mRNA overexpression in psychosis. (англ.) // Neuroreport. — 2007. — 8 January (vol. 18, no. 1). — P. 57—60. — doi:10.1097/WNR.0b013e32800fefd7. — PMID 17259861.
- ↑ 1 2 Dong E., Nelson M., Grayson D.R., Costa E., Guidotti A. Clozapine and sulpiride but not haloperidol or olanzapine activate brain DNA demethylation (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2008. — August. — doi:10.1073/pnas.0805493105. — PMID 18757738.
- ↑ Fatemi S. H., Kroll J. L., Stary J. M. Altered levels of Reelin and its isoforms in schizophrenia and mood disorders. (англ.) // Neuroreport. — 2001. — 29 October (vol. 12, no. 15). — P. 3209—3215. — doi:10.1097/00001756-200110290-00014. — PMID 11711858.
- ↑ Fatemi S. H., Emamian E. S., Kist D., Sidwell R. W., Nakajima K., Akhter P., Shier A., Sheikh S., Bailey K. Defective corticogenesis and reduction in Reelin immunoreactivity in cortex and hippocampus of prenatally infected neonatal mice. (англ.) // Molecular Psychiatry. — 1999. — March (vol. 4, no. 2). — P. 145—154. — doi:10.1038/sj.mp.4000520. — PMID 10208446.
- ↑ Meyer U., Nyffeler M., Yee B.K., Knuesel I., Feldon J. Adult brain and behavioral pathological markers of prenatal immune challenge during early/middle and late fetal development in mice (англ.) // Brain Behav Immun[англ.] : journal. — 2007. — doi:10.1016/j.bbi.2007.09.012. — PMID 18023140.
- ↑ Kamnasaran D., Muir W. J., Ferguson-Smith M. A., Cox D. W. Disruption of the neuronal PAS3 gene in a family affected with schizophrenia. (англ.) // Journal Of Medical Genetics. — 2003. — May (vol. 40, no. 5). — P. 325—332. — doi:10.1136/jmg.40.5.325. — PMID 12746393.
- ↑ Erbel-Sieler C., Dudley C., Zhou Y., Wu X., Estill S. J., Han T., Diaz-Arrastia R., Brunskill E. W., Potter S. S., McKnight S. L. Behavioral and regulatory abnormalities in mice deficient in the NPAS1 and NPAS3 transcription factors. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 2004. — 14 September (vol. 101, no. 37). — P. 13648—13653. — doi:10.1073/pnas.0405310101. — PMID 15347806.
- ↑ Chen Z., Schwahn B.C., Wu Q., He X., Rozen R. Postnatal cerebellar defects in mice deficient in methylenetetrahydrofolate reductase (англ.) // Int. J. Dev. Neurosci.[англ.] : journal. — 2005. — August (vol. 23, no. 5). — P. 465—474. — doi:10.1016/j.ijdevneu.2005.05.007. — PMID 15979267. Архивировано 28 августа 2017 года.
- ↑ 1 2 Wedenoja J., Loukola A., Tuulio-Henriksson A., Paunio T., Ekelund J., Silander K., Varilo T., Heikkilä K., Suvisaari J., Partonen T., Lönnqvist J., Peltonen L. Replication of linkage on chromosome 7q22 and association of the regional Reelin gene with working memory in schizophrenia families (англ.) // Mol Psychiatry[англ.] : journal. — 2007. — doi:10.1038/sj.mp.4002047. — PMID 17684500.
- ↑ Wedenoja J., Tuulio-Henriksson A., Suvisaari J., Loukola A., Paunio T., Partonen T., Varilo T., Lönnqvist J., Peltonen L. Replication of Association Between Working Memory and Reelin, a Potential Modifier Gene in Schizophrenia (англ.) // Biol. Psychiatry[англ.] : journal. — 2009. — November. — doi:10.1016/j.biopsych.2009.09.026. — PMID 19922905. Архивировано 28 августа 2017 года.
- ↑ Shifman S., Johannesson M., Bronstein M., Chen S.X., Collier D.A., Craddock N.J., Kendler K.S., Li T., O'Donovan M., O'Neill F.A., Owen M.J., Walsh D., Weinberger D.R., Sun C., Flint J., Darvasi A. Genome-Wide Association Identifies a Common Variant in the Reelin Gene That Increases the Risk of Schizophrenia Only in Women (англ.) // PLoS Genet.[англ.] : journal. — 2008. — Vol. 4, no. 2. — P. e28. — doi:10.1371/journal.pgen.0040028. — PMID 18282107. free full text Архивировано 13 января 2013 года.
- ↑ Gregório S.P., Sallet P.C., Do K.A., Lin E., Gattaz W.F., Dias-Neto E. Polymorphisms in genes involved in neurodevelopment may be associated with altered brain morphology in schizophrenia: Preliminary evidence (англ.) // Psychiatry Res[англ.] : journal. — 2008. — December. — doi:10.1016/j.psychres.2007.08.011. — PMID 19054571. Архивировано 28 августа 2017 года.
- ↑ Suzuki K., Nakamura K., Iwata Y., Sekine Y., Kawai M., Sugihara G., Tsuchiya K.J., Suda S., Matsuzaki H., Takei N., Hashimoto K., Mori N. Decreased expression of reelin receptor VLDLR in peripheral lymphocytes of drug-naive schizophrenic patients (англ.) : journal. — 2007. — doi:10.1016/j.schres.2007.09.029. — PMID 17936586.
- ↑ Sweet R.A., Henteleff R.A., Zhang W., Sampson A.R., Lewis D.A. Reduced Dendritic Spine Density in Auditory Cortex of Subjects with Schizophrenia (англ.) // Neuropsychopharmacology[англ.] : journal. — Nature Publishing Group, 2008. — May. — doi:10.1038/npp.2008.67. — PMID 18463626.
- ↑ Glantz L.A., Lewis D.A. Decreased dendritic spine density on prefrontal cortical pyramidal neurons in schizophrenia (англ.) // Arch. Gen. Psychiatry[англ.] : journal. — 2000. — January (vol. 57, no. 1). — P. 65—73. — PMID 10632234. (недоступная ссылка)
- ↑ Rodriguez M.A., Pesold C., Liu W.S., et al. Colocalization of integrin receptors and reelin in dendritic spine postsynaptic densities of adult nonhuman primate cortex (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2000. — March (vol. 97, no. 7). — P. 3550—3555. — doi:10.1073/pnas.050589797. — PMID 10725376. — PMC 16277.
- ↑ Costa E., Davis J., Grayson D.R., Guidotti A., Pappas G.D., Pesold C. Dendritic spine hypoplasticity and downregulation of reelin and GABAergic tone in schizophrenia vulnerability (англ.) // Neurobiol. Dis.[англ.] : journal. — 2001. — October (vol. 8, no. 5). — P. 723—742. — doi:10.1006/nbdi.2001.0436. — PMID 11592844. Архивировано 28 августа 2017 года.
- ↑ Gene Overview of All Published Schizophrenia-Association Studies for GRIN2B Архивная копия от 13 сентября 2010 на Wayback Machine — Сводка публикаций по гену GRIN2B в генетической базе данных по шизофрении.
- ↑ 1 2 Wang G.S., Hong C.J., Yen T.Y., Huang H.Y., Ou Y., Huang T.N., Jung W.G., Kuo T.Y., Sheng M., Wang T.F., Hsueh Y.P. Transcriptional modification by a CASK-interacting nucleosome assembly protein (англ.) // Neuron : journal. — Cell Press, 2004. — April (vol. 42, no. 1). — P. 113—128. — PMID 15066269. Архивировано 28 августа 2017 года.
- ↑ Podhorna J., Didriksen M. The heterozygous reeler mouse: behavioural phenotype (англ.) // Behav. Brain Res.[англ.] : journal. — 2004. — August (vol. 153, no. 1). — P. 43—54. — doi:10.1016/j.bbr.2003.10.033. — PMID 15219705.
- ↑ Goes F. S., Willour V. L., Zandi P. P., Belmonte P. L., MacKinnon D. F., Mondimore F. M., Schweizer B., National Institute of Mental Health Genetics Initiative Bipolar Disorder Consortium, DePaulo Jr. J. R., Gershon E. S., McMahon F. J., Potash J. B. Sex-specific association of the Reelin gene with bipolar disorder. (англ.) // American Journal Of Medical Genetics. Part B, Neuropsychiatric Genetics : The Official Publication Of The International Society Of Psychiatric Genetics. — 2010. — 5 March (vol. 153B, no. 2). — P. 549—553. — doi:10.1002/ajmg.b.31018. — PMID 19691043.
- ↑ Haas C. A., Dudeck O., Kirsch M., Huszka C., Kann G., Pollak S., Zentner J., Frotscher M. Role for reelin in the development of granule cell dispersion in temporal lobe epilepsy. (англ.) // The Journal Of Neuroscience : The Official Journal Of The Society For Neuroscience. — 2002. — 15 July (vol. 22, no. 14). — P. 5797—5802. — doi:10.1523/JNEUROSCI.22-14-05797.2002. — PMID 12122039.
- ↑ Heinrich C., Nitta N., Flubacher A., Müller M., Fahrner A., Kirsch M., Freiman T., Suzuki F., Depaulis A., Frotscher M., Haas C. A. Reelin deficiency and displacement of mature neurons, but not neurogenesis, underlie the formation of granule cell dispersion in the epileptic hippocampus. (англ.) // The Journal Of Neuroscience : The Official Journal Of The Society For Neuroscience. — 2006. — 26 April (vol. 26, no. 17). — P. 4701—4713. — doi:10.1523/JNEUROSCI.5516-05.2006. — PMID 16641251.
- ↑ Haas C.A., Frotscher M. Reelin deficiency causes granule cell dispersion in epilepsy (англ.) // Exp Brain Res[англ.] : journal. — 2009. — July. — doi:10.1007/s00221-009-1948-5. — PMID 19633980.
- ↑ Kobow K., Jeske I., Hildebrandt M., Hauke J., Hahnen E., Buslei R., Buchfelder M., Weigel D., Stefan H., Kasper B., Pauli E., Blümcke I. Increased Reelin Promoter Methylation Is Associated With Granule Cell Dispersion in Human Temporal Lobe Epilepsy (англ.) // J. Neuropathol. Exp. Neurol.[англ.] : journal. — 2009. — March. — doi:10.1097/NEN.0b013e31819ba737. — PMID 19287316.
- ↑ Gong C., Wang T. W., Huang H. S., Parent J. M. Reelin regulates neuronal progenitor migration in intact and epileptic hippocampus. (англ.) // The Journal Of Neuroscience : The Official Journal Of The Society For Neuroscience. — 2007. — 21 February (vol. 27, no. 8). — P. 1803—1811. — doi:10.1523/JNEUROSCI.3111-06.2007. — PMID 17314278.
- ↑ Müller M.C., Osswald M., Tinnes S., Häussler U., Jacobi A., Förster E., Frotscher M., Haas C.A. Exogenous reelin prevents granule cell dispersion in experimental epilepsy (англ.) // Exp. Neurol.[англ.] : journal. — 2009. — January. — doi:10.1016/j.expneurol.2008.12.029. — PMID 19185570. Архивировано 28 августа 2017 года.
- ↑ Botella-López A., Burgaya F., Gavín R., García-Ayllón M. S., Gómez-Tortosa E., Peña-Casanova J., Ureña J. M., Del Río J. A., Blesa R., Soriano E., Sáez-Valero J. Reelin expression and glycosylation patterns are altered in Alzheimer's disease. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 2006. — 4 April (vol. 103, no. 14). — P. 5573—5578. — doi:10.1073/pnas.0601279103. — PMID 16567613.
- ↑ Wirths O., Multhaup G., Czech C., Blanchard V., Tremp G., Pradier L., Beyreuther K., Bayer T. A. Reelin in plaques of beta-amyloid precursor protein and presenilin-1 double-transgenic mice. (англ.) // Neuroscience Letters. — 2001. — 28 December (vol. 316, no. 3). — P. 145—148. — doi:10.1016/s0304-3940(01)02399-0. — PMID 11744223.
- ↑ Baloyannis S.J. Morphological and morphometric alterations of Cajal-Retzius cells in early cases of Alzheimer's disease: a Golgi and electron microscope study (англ.) // Int. J. Neurosci.[англ.] : journal. — 2005. — July (vol. 115, no. 7). — P. 965—980. — doi:10.1080/00207450590901396. — PMID 16051543.
- ↑ Baloyannis S.J., Costa V., Mauroudis I., Psaroulis D., Manolides S.L., Manolides L.S. Dendritic and spinal pathology in the acoustic cortex in Alzheimer's disease: morphological and morphometric estimation by Golgi technique and electron microscopy (англ.) // Acta Otolaryngol.[англ.] : journal. — 2007. — April (vol. 127, no. 4). — P. 351—354. — doi:10.1080/00016480601126986. — PMID 17453452.
- ↑ Публикация, обзор работы с комментариями исследователей на форуме исследования болезни Альцгеймера, и новостная заметка в научном блоге:
- Hoe H.S., Lee K.J., Carney R.S., Lee J., Markova A., Lee J.Y., Howell B.W., Hyman B.T., Pak D.T., Bu G., Rebeck G.W. Interaction of reelin with amyloid precursor protein promotes neurite outgrowth (англ.) // J. Neurosci.[англ.] : journal. — 2009. — June (vol. 29, no. 23). — P. 7459—7473. — doi:10.1523/JNEUROSCI.4872-08.2009. — PMID 19515914.
- Another Take on APP and Neurite Outgrowth—The Role of Reelin (Tom Fagan, 19 June 2009). Дата обращения: 24 июня 2009. Архивировано 26 января 2012 года.
- Protein linked to Alzheimer's disease doesn't act alone (www.sciencecodex.com). Дата обращения: 16 июня 2009. Архивировано 26 января 2012 года.; перевод: Взаимодействие предшественника бета-амилоида с рилином (недоступная ссылка) (блог neuroscience.ru)
- ↑ Научная публикация, описание в новостной заметке:
- Durakoglugil M. S., Chen Y., White C. L., Kavalali E. T., Herz J. Reelin signaling antagonizes beta-amyloid at the synapse. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 2009. — 15 September (vol. 106, no. 37). — P. 15938—15943. — doi:10.1073/pnas.0908176106. — PMID 19805234.
- New findings about brain proteins suggest possible way to fight Alzheimer’s Архивная копия от 10 августа 2014 на Wayback Machine — Science Centric
- ↑ AOKI Takeya, MIZUKI Yasushi, TERASHIMA Toshio. Relation between schizophrenia and Alzheimer's disease: the reelin signaling pathway (англ.) // Psychogeriatrics. — 2005. — June (vol. 5, no. 2). — P. 42—47. — ISSN 1346-3500. — doi:10.1111/j.1479-8301.2005.00091.x.
- ↑ 1 2 Durakoglugil M. S., Chen Y., White C. L., Kavalali E. T., Herz J. Reelin signaling antagonizes beta-amyloid at the synapse. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 2009. — 15 September (vol. 106, no. 37). — P. 15938—15943. — doi:10.1073/pnas.0908176106. — PMID 19805234.
- ↑ Devlin B., Bennett P., Dawson G., Figlewicz D. A., Grigorenko E. L., McMahon W., Minshew N., Pauls D., Smith M., Spence M. A., Rodier P. M., Stodgell C., Schellenberg G. D., CPEA Genetics Network. Alleles of a reelin CGG repeat do not convey liability to autism in a sample from the CPEA network. (англ.) // American Journal Of Medical Genetics. Part B, Neuropsychiatric Genetics : The Official Publication Of The International Society Of Psychiatric Genetics. — 2004. — 1 April (vol. 126B, no. 1). — P. 46—50. — doi:10.1002/ajmg.b.20125. — PMID 15048647.
- ↑ Li J., Nguyen L., Gleason C., Lotspeich L., Spiker D., Risch N., Myers R. M. Lack of evidence for an association between WNT2 and RELN polymorphisms and autism. (англ.) // American Journal Of Medical Genetics. Part B, Neuropsychiatric Genetics : The Official Publication Of The International Society Of Psychiatric Genetics. — 2004. — 1 April (vol. 126B, no. 1). — P. 51—57. — doi:10.1002/ajmg.b.20122. — PMID 15048648.
- ↑ Dutta S., Sinha S., Ghosh S., Chatterjee A., Ahmed S., Usha R. Genetic analysis of reelin gene (RELN) SNPs: No association with autism spectrum disorder in the Indian population (англ.) // Neurosci. Lett.[англ.] : journal. — 2008. — August (vol. 441, no. 1). — P. 56—60. — doi:10.1016/j.neulet.2008.06.022. — PMID 18597938. Архивировано 28 августа 2017 года.
- ↑ Serajee F. J., Zhong H., Mahbubul Huq A. H. Association of Reelin gene polymorphisms with autism. (англ.) // Genomics. — 2006. — January (vol. 87, no. 1). — P. 75—83. — doi:10.1016/j.ygeno.2005.09.008. — PMID 16311013.
- ↑ Skaar D. A., Shao Y., Haines J. L., Stenger J. E., Jaworski J., Martin E. R., DeLong G. R., Moore J. H., McCauley J. L., Sutcliffe J. S., Ashley-Koch A. E., Cuccaro M. L., Folstein S. E., Gilbert J. R., Pericak-Vance M. A. Analysis of the RELN gene as a genetic risk factor for autism. (англ.) // Molecular Psychiatry. — 2005. — June (vol. 10, no. 6). — P. 563—571. — doi:10.1038/sj.mp.4001614. — PMID 15558079.
- ↑ Fatemi S.H., Stary J.M., Halt A.R., Realmuto G.R. Dysregulation of Reelin and Bcl-2 proteins in autistic cerebellum (англ.) // J Autism Dev Disord[англ.] : journal. — 2001. — December (vol. 31, no. 6). — P. 529—535. — PMID 11814262.
- ↑ Fatemi S.H., Stary J.M., Egan E.A. Reduced blood levels of reelin as a vulnerability factor in pathophysiology of autistic disorder (англ.) // Cell. Mol. Neurobiol. : journal. — 2002. — April (vol. 22, no. 2). — P. 139—152. — PMID 12363196.
- ↑ Sato Norihiro, Fukushima Noriyoshi, Chang Rubens, Matsubayashi Hiroyuki, Goggins Michael. Differential and Epigenetic Gene Expression Profiling Identifies Frequent Disruption of the RELN Pathway in Pancreatic Cancers (англ.) // Gastroenterology. — 2006. — February (vol. 130, no. 2). — P. 548—565. — ISSN 0016-5085. — doi:10.1053/j.gastro.2005.11.008. — PMID 16472607.
- ↑ Perrone Giuseppe, Vincenzi Bruno, Zagami Mariagiovanna, Santini Daniele, Panteri Roger, Flammia Gerardo, Verzì Alfio, Lepanto Daniela, Morini Sergio, Russo Antonio, Bazan Vivian, Tomasino Rosa M, Morello Vincenza, Tonini Giuseppe, Rabitti Carla. Reelin expression in human prostate cancer: a marker of tumor aggressiveness based on correlation with grade (англ.) // Modern Pathology. — 2007. — 2 February (vol. 20, no. 3). — P. 344—351. — ISSN 0893-3952. — doi:10.1038/modpathol.3800743. — PMID 17277764.
- ↑ Seigel G.M., Hackam A.S., Ganguly A., Mandell L.M., Gonzalez-Fernandez F. Human embryonic and neuronal stem cell markers in retinoblastoma (англ.) // Mol. Vis.[англ.] : journal. — 2007. — Vol. 13. — P. 823—832. — PMID 17615543. Архивировано 9 мая 2008 года.
- ↑ Schrauwen I., Ealy M., Huentelman M.J., Thys M., Homer N., Vanderstraeten K., Fransen E., Corneveaux J.J., Craig D.W., Claustres M., Cremers C.W., Dhooge I., Van de Heyning P., Vincent R., Offeciers E., Smith R.J., Van Camp G. A Genome-wide Analysis Identifies Genetic Variants in the RELN Gene Associated with Otosclerosis (англ.) // Am. J. Hum. Genet.[англ.] : journal. — 2009. — February. — doi:10.1016/j.ajhg.2009.01.023. — PMID 19230858.
- ↑ Delahaye N.F., Coltel N., Puthier D., Barbier M., Benech P., Joly F., Iraqi F.A., Grau G.E., Nguyen C., Rihet P. Gene expression analysis reveals early changes in several molecular pathways in cerebral malaria-susceptible mice versus cerebral malaria-resistant mice (англ.) // BMC Genomics[англ.] : journal. — 2007. — Vol. 8. — P. 452. — doi:10.1186/1471-2164-8-452. — PMID 18062806. — PMC 2246131. Архивировано 15 октября 2009 года.
- ↑ Knuesel Irene, Nyffeler Myriel, Mormède Cecile, Muhia Mary, Meyer Urs, Pietropaolo Susanna, Yee Benjamin K., Pryce Christopher R., LaFerla Frank M., Marighetto Aline, Feldon Joram. Age-related accumulation of Reelin in amyloid-like deposits (англ.) // Neurobiology of Aging. — 2009. — May (vol. 30, no. 5). — P. 697—716. — ISSN 0197-4580. — doi:10.1016/j.neurobiolaging.2007.08.011. — PMID 17904250.
- ↑ Madhusudan Amrita, Sidler Corinne, Knuesel Irene. Accumulation of reelin-positive plaques is accompanied by a decline in basal forebrain projection neurons during normal aging (англ.) // European Journal of Neuroscience. — 2009. — September (vol. 30, no. 6). — P. 1064—1076. — ISSN 0953-816X. — doi:10.1111/j.1460-9568.2009.06884.x. — PMID 19735296.
- ↑ 1 2 Smit-Rigter L.A., Champagne D.L., van Hooft J.A. Lifelong impact of variations in maternal care on dendritic structure and function of cortical layer 2/3 pyramidal neurons in rat offspring (англ.) // PLoS ONE : journal. — 2009. — Vol. 4, no. 4. — P. e5167. — doi:10.1371/journal.pone.0005167. — PMID 19357777. — PMC 2663818.
- ↑ Weaver I.C., Meaney M.J., Szyf M. Maternal care effects on the hippocampal transcriptome and anxiety-mediated behaviors in the offspring that are reversible in adulthood (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2006. — February (vol. 103, no. 9). — P. 3480—3485. — doi:10.1073/pnas.0507526103. — PMID 16484373. — PMC 1413873.
- ↑ Cassidy A.W., Mulvany S.K., Pangalos M.N., Murphy K.J., Regan C.M. Developmental emergence of reelin deficits in the prefrontal cortex of Wistar rats reared in social isolation (англ.) // Neuroscience[англ.] : journal. — Elsevier, 2009. — December. — doi:10.1016/j.neuroscience.2009.12.045. — PMID 20035841. Архивировано 28 августа 2017 года.
- ↑ Lussier A.L., Caruncho H.J., Kalynchuk L.E. Repeated exposure to corticosterone, but not restraint, decreases the number of reelin-positive cells in the adult rat hippocampus (англ.) // Neurosci. Lett.[англ.] : journal. — 2009. — May. — doi:10.1016/j.neulet.2009.05.050. — PMID 19477232.
- ↑ Lintas C., Persico A.M. Neocortical RELN promoter methylation increases significantly after puberty (англ.) // Neuroreport[англ.] : journal. — 2009. — December. — doi:10.1097/WNR.0b013e328334b343. — PMID 19952965. Архивировано 31 июля 2017 года.
- ↑ Sui L., Ren W.W., Li B.M. Administration of thyroid hormone increases reelin and brain-derived neurotrophic factor expression in rat hippocampus in vivo (англ.) // Brain Res[англ.] : journal. — 2009. — December. — doi:10.1016/j.brainres.2009.12.010. — PMID 20018181. Архивировано 28 августа 2017 года.
- ↑ Fatemi S.H., Reutiman T.J., Folsom T.D. Chronic psychotropic drug treatment causes differential expression of Reelin signaling system in frontal cortex of rats (англ.) // Schizophrenia Research[англ.] : journal. — Elsevier, 2009. — April. — doi:10.1016/j.schres.2009.03.002. — PMID 19359144. Архивировано 28 августа 2017 года.
Эта статья входит в число хороших статей русскоязычного раздела Википедии. |
Некоторые внешние ссылки в этой статье ведут на сайты, занесённые в спам-лист |