Теломеры
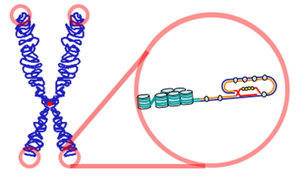
Теломеры (от греч. telos — конец и meros — часть) — концевые участки хромосом. Теломерные участки хромосом характеризуются отсутствием способности к соединению с другими хромосомами или их фрагментами и выполняют защитную функцию.
Термин «теломера» предложил Г.Мёллер в 1932 г[1].
У большинства эукариот теломеры состоят из специализированной линейной хромосомной ДНК, состоящей из коротких тандемных повторов. В теломерных участках хромосом ДНК вместе со специфически связывающимися с теломерными ДНК-повторами белками образует нуклеопротеидный комплекс — конститутивный (структурный) теломерный гетерохроматин. Теломерные повторы — весьма консервативные последовательности, например повторы всех позвоночных состоят из шести нуклеотидов TTAGGG, повторы всех насекомых — TTAGG, повторы большинства растений — TTTAGGG.
Ученые из университета Кардиффа (Cardiff University) установили, что критическая длина человеческой теломеры, при которой хромосомы начинают соединяться друг с другом, составляет 12,8 теломерных повторов[2].
В каждом цикле деления теломеры клетки укорачиваются из-за неспособности ДНК-полимеразы синтезировать копию ДНК с самого конца. Она в состоянии лишь добавлять нуклеотиды к уже существующей 3’-гидроксильной группе. По этой причине ДНК-полимераза нуждается в праймере, к которому она могла бы добавить первый нуклеотид. Данный феномен носит название концевой недорепликации и является одним из важнейших факторов биологического старения. Тем не менее, вследствие этого явления теломеры должны укорачиваться весьма медленно - по несколько (3-6) нуклеотидов за клеточный цикл, т.е. за количество делений, соответствующее пределу Хейфлика, они укоротятся всего на 150-300 нуклеотидов. В настоящее время предложена эпигенетическая теория старения, которая предполагает, что эрозия теломер ускоряется в десятки и сотни раз из-за рекомбинаций в их ДНК, вызванных функционированием клеточных систем репарации ДНК. Активность данных систем инициируется повреждением ДНК, обусловленном прежде всего дерепрессирующимися с возрастом мобильными элементами генома, что и предопределяет старение как биологический феномен[3].
Существует специальный фермент — теломераза, который при помощи собственной РНК-матрицы достраивает теломерные повторы и удлиняет теломеры. В большинстве дифференцированных клеток теломераза заблокирована, однако активна в стволовых и половых клетках.
За открытие защитных механизмов хромосом от концевой недорепликации с помощью теломер и теломеразы в 2009 году присуждена Нобелевская премия по физиологии и медицине австралийке, работающей в США, Элизабет Блекберн (Elizabeth Blackburn), американке Кэрол Грейдер (Carol Greider) и ее соотечественнику Джеку Шостак (Jack Szostack).
Примечания
- ↑ Muller H.J. Further studies on the nature and causes of gene mutations // Proc. Sixth Int. Congr. Genet. 1932. V.1. P.213-255.
- ↑ The nature of telomere fusion and a definition of the critical telomere length in human cells.
- ↑ Галицкий В.А. (2009). "Эпигенетическая природа старения" (PDF). Цитология. 51: 388—397.
См. также
Ссылки
Это заготовка статьи по цитологии. Помогите Википедии, дополнив её. |
В статье не хватает ссылок на источники (см. рекомендации по поиску). |