Signaalpeptide
Een signaalpeptide (ook wel signaalsequentie of lokaliseringssequentie genoemd) is een korte peptideketen, gewoonlijk 16–30 aminozuren lang,[1] die voorkomt aan de N-terminus (begin) van veel nieuw gesynthetiseerde secretie-eiwitten.[2] Secretie-eiwitten zijn eiwitten die vrijkomen uit (klier)cellen, een rol spelen in organellen zoals het endoplasmatisch reticulum en golgiapparaat, en in celmembranen worden ingebouwd. Signaalpeptiden zorgen ervoor dat deze eiwitten naar hun juiste bestemming worden vervoerd. Bij aankomst worden de signaalpeptiden gewoonlijk afgesplitst.
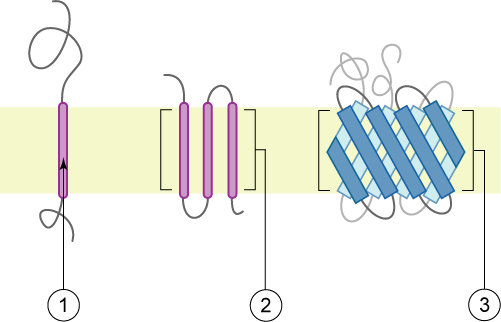
Signaalpeptiden komen voor op zogenaamde type I-membraaneiwitten.[a] Bij de membraaneiwitten type II en type III is geen echt signaalpeptide aanwezig: daar fungeert het voorste transmembraandomein als signaalsequentie. Een transmembraandomein lijkt biochemisch gezien op een signaalpeptide, afgezien van het feit dat het niet wordt afgesplitst. Transmembraandomeinen zijn vergelijkbaar met specialistische peptiden die een eiwit naar zijn doel vervoeren (target peptides).
Functie
bewerkenSignaalpeptiden hebben als functie een cel ertoe aan te zetten een eiwit te transporteren (transloceren), naar een specifieke bestemming, gewoonlijk het celmembraan. In prokaryoten sturen signaalpeptiden het nieuw gesynthetiseerde eiwit naar het zogenaamde SecYEG-eiwitkanaal, dat aanwezig is in hun membraan. In eukaryoten bestaat een soortgelijk systeem, waarbij het signaalpeptide het nieuw gesynthetiseerde eiwit naar het Sec61-kanaal stuurt, dat qua structuur en sequentie homologie vertoont met een SecYEG-kanaal, maar in het endoplasmatisch reticulum aanwezig is.[3] Zowel de SecYEG- als de Sec61-kanalen worden gewoonlijk de translocon genoemd; de doorvoer via dit kanaal staat bekend als translocatie. Wanneer secretie-eiwitten zich door het kanaal verplaatsen, kunnen transmembraandomeinen via een doorgang (naast het translocon) diffunderen om zich in het omringende membraan te vestigen.
Signaalpeptidestructuur
bewerkenHet middengedeelte van een signaalpeptide bevat een lange reeks hydrofobe aminozuren (ongeveer 5-16 residuen lang).[b] Dit middengedeelte heeft de neiging om een alfa-helixstructuur aan te nemen en wordt de "h-regio" genoemd.[4] Het begin van een signaalpeptide bestaat vaak uit een aantal positief geladen aminozuren die een rol spelen bij het tot stand brengen van de juiste topologie van het polypeptide tijdens translocatie: de positive-inside rule.[5] Omdat dit gedeelte van het signaalpeptide dicht bij de N-terminus ligt, wordt dit de "n-regio" genoemd.
Aan het einde van het signaalpeptide bevindt zich een kenmerkend stuk vaak polaire aminozuren dat door signaalpeptidase wordt afgesplitst: de splitsingsplaats (cleavage site). Deze splitsingsplaats is afwezig in transmembraandomeinen die dienen als signaalpeptiden; deze worden soms signaalankersequenties genoemd. Signaalpeptidase kan tijdens of na voltooiing van translocatie het signaalpeptide afsplitsen van de rest van het eiwit.[6] De vrijgekomen signaalpeptide worden vervolgens door specifieke proteasen afgebroken.
Signaalpeptiden brengen het eiwit altijd naar een specifieke cellulaire bestemming. Een signaalpeptide dat gericht is naar het mitochondrium vertoont een alternerend patroon van kleine, positief geladen en hydrofobe stukken en verschilt in lengte en structuur van andere signaalpeptiden. Op de celkern gerichte signaalpeptiden kunnen op zowel de N-terminus als de C-terminus van een eiwit voorkomen en worden in de meeste gevallen niet van het eiwit afgesplitst.
Het is mogelijk om de aminozuurvolgorde van een N-terminale signaalpeptide te bepalen met behulp van Edman-degradatie, een iteratieve procedure waarin de aminozuren een voor een worden afgesplitst.[7] Uit dergelijke onderzoeken blijkt dat zowel de aminozuurvolgorde als de lengte van signaalpeptiden enorm variëren tussen soorten.
Co-translationele versus post-translationele translocatie
bewerkenIn zowel prokaryoten als eukaryoten kunnen signaalpeptiden co-translationeel of post-translationeel werken, respectievelijk tijdens of na de translatie.
De co-translationele route wordt in gang gezet wanneer het signaalpeptide uit het ribosoom vrijkomt en wordt herkend door het signaalherkenningsdeeltje (signal-recognition particle, SRP).[8] SRP stopt dan verdere translatie (deze translationele stopprocedure treedt alleen op bij eukaryoten) en stuurt het signaalsequentie-ribosoom-mRNA-complex naar de SRP-receptor, die aanwezig is op het oppervlak van ofwel het plasmamembraan (in prokaryoten) of het ER (in eukaryoten).[9] Zodra het complex aan een specifieke receptor op het membraanoppervlak bindt, wordt het signaalpeptide in het naastgelegen translocon ingevoegd. Ribosomen worden dan fysiek gekoppeld aan de intracellulaire kant van het translocon en de eiwitsynthese wordt vervolgd.[10]
Het post-translationele traject wordt pas ingezet nadat de eiwitsynthese is voltooid. In prokaryoten wordt de signaalsequentie van post-translationele substraten herkend door het SecB chaperone-eiwit dat het eiwit naar het SecA ATPase overbrengt, dat op zijn beurt het eiwit door de translocon pompt. Hoewel bekend is dat posttranslationele translocatie ook plaatsvindt in eukaryoten, wordt dit proces nog slecht begrepen. Het is echter wel bekend dat posttranslationele translocatie in gist naast de translocon 2 extra membraangebonden eiwitten vereist: Sec62 en Sec63.[11]
Signaalpeptiden bepalen de efficiëntie van de secretie
bewerkenSignaalpeptiden zijn extreem heterogeen en veel prokaryotische en eukaryote signaalpeptiden zijn functioneel uitwisselbaar, zelfs tussen verschillende soorten. De efficiëntie van eiwitsecretie wordt sterk bepaald door de aanwezigheid van een signaalpeptide.[12][13]
Functies op nucleotide-niveau
bewerkenIn gewervelden kan het gedeelte van het mRNA dat codeert voor het signaalpeptide (de signaalsequentie coderende regio of SSCR) functioneren als een RNA-fragment met specifieke activiteiten. SSCR's bevorderen enerzijds de export van mRNA vanuit de celkern en anderzijds de juiste lokalisatie naar het oppervlak van het endoplasmatisch reticulum. Bovendien hebben SSCR's specifieke sequentiekenmerken: ze bevatten nauwelijks adenine-nucleotiden, zijn verrijkt met bepaalde sequentiemotieven en zijn onverwacht vaak aanwezig in het eerste exon (het coderende deel van een gen).[14][15]
Signaalpeptide minder secretie
bewerkenEiwitten zonder signaalpeptiden kunnen ook worden afgescheiden, namelijk via ongebruikelijke mechanismen; voorbeelden daarvan zijn Interleukine en Galectine.[16] Het secretieproces waarmee dergelijke secretoire eiwitten worden afgescheiden naar de extracellulaire vloeistof, heeft men unconvetional protein secretion (onconventionele eiwitsecretie) (UPS) genoemd. In planten kan zelfs 50% van de uitgescheiden eiwitten UPS-afhankelijk zijn.[17]
Zie ook
bewerkenNoten
- ↑ Deze eiwitten zijn dusdanig in het membraan ingepast, dat zij een transportkanaal vormen voor bepaalde stoffen die deze membraan moeten passeren (zie de figuur). Het verschil tussen de typeringen I, II en III betreft de manier waarop de signaalpeptiden gebonden zijn aan een membraan.
- ↑ Met een residu wordt een aminozuur-eenheid binnen de keten bedoeld.
Bronnen
- ↑ (en) Kapp, K (1 januari 2013). Post-Targeting Functions of Signal Peptides. Landes Bioscience.
- ↑ (en) Blobel G, Dobberstein B (dec 1975). Transfer of proteins across membranes. I. Presence of proteolytically processed and unprocessed nascent immunoglobulin light chains on membrane-bound ribosomes of murine myeloma.. J Cell Biol 67 (3): 835–51. PMID 811671. PMC 2111658. DOI: 10.1083/jcb.67.3.835.
- ↑ (en) Rapoport T. (2007). Protein translocation across the eukaryotic endoplasmic reticulum and bacterial plasma membranes.. Nature 450 (7170): 663–669. PMID 18046402. DOI: 10.1038/nature06384.
- ↑ (en) Käll, Lukas (2004). A Combined Transmembrane Topology and Signal Peptide Prediction Method. J. Mol. Biol. 338 (5): 1027–1036. PMID 15111065. DOI: 10.1016/j.jmb.2004.03.016.
- ↑ (en) von Heijne, G. (1988). Topogenic signals in integral membrane proteins. Eur J Biochem 174 (4): 671–678. PMID 3134198. DOI: 10.1111/j.1432-1033.1988.tb14150.x.
- ↑ (en) Neil Campbell, Jane Reece en Lawrence Mitchell (1999). Biology. Fifth edition. Benjamin Cummings, p.310. ISBN 0805365737.
- ↑ (en) "26.6 Peptide Sequencing: The Edman Degradation", Chemistry LibreTexts, 26 augustus 2015. Geraadpleegd op 27 september 2018.
- ↑ (en) Walter, P, Ibrahimi I, Blobel, G (1981). Translocation of proteins across the endoplasmic reticulum. I. Signal recognition protein (SRP) binds to in-vitro-assembled polysomes synthesizing secretory protein. JCB 91 (2 Pt1): 545–50. PMID: 7309795. PMC: 2111968. DOI:10.1083/jcb.91.2.545.
- ↑ (en) Gilmore, R, Blobel G, Walter, P (1982). Protein translocation across the endoplasmic reticulum. I. Detection in the microsomal membrane of a receptor for the signal recognition particle. JCB 95 (2 Pt1): 463–9. PMID: 6292235. PMC: 2112970. DOI:10.1083/jcb.95.2.463.
- ↑ (en) Görlich, D, Prehn S, Hartmann E, Kalies KU, Rapoport TA (1992). A mammalian homolog of SEC61p and SECYp is associated with ribosomes and nascent polypeptides during translocation. Cell 71 (3): 489–503. PMID: 1423609. DOI:10.1016/0092-8674(92)90517-G.
- ↑ (en) Panzner, S (1995). Posttranslational protein transport in yeast reconstituted with a purified complex of Sec proteins and Kar2p. Cell 81 (4): 561–570. ISSN:0092-8674. PMID: 7758110. DOI:10.1016/0092-8674(95)90077-2.
- ↑ (en) Kober, L, Zehe C, Bode J (april 2013). Optimized signal peptides for the development of high expressing CHO cell lines. Biotechnol. Bioeng. 110 (4): 1164–73. PMID: 23124363. DOI:10.1002/bit.24776.
- ↑ (en) von Heijne, G (1985). Signal sequences: The limits of variation. Journal Mol. Biol. 184 (1): 99–105. PMID: 4032478. DOI:10.1016/0022-2836(85)90046-4.
- ↑ (en) Palazzo, Alexander F. (2007). The Signal Sequence Coding Region Promotes Nuclear Export of mRNA. PLoS Biology 5 (12): e322. ISSN: 1544-9173. PMID 18052610. PMC 2100149. DOI: 10.1371/journal.pbio.0050322.
- ↑ (en) Cenik, Can et al. (2011). Genome Analysis Reveals Interplay between 5′UTR Introns and Nuclear mRNA Export for Secretory and Mitochondrial Genes. PLoS Genetics 7 (4): e1001366. ISSN: 1553-7404. PMID 21533221. PMC 3077370. DOI: 10.1371/journal.pgen.1001366.
- ↑ (en) Nickel, W (2008). Unconventional mechanisms of protein transport to the cell surface of eukaryotic cells.. Annual Review of Cell and Developmental Biology 24: 287–308. PMID 18590485. DOI: 10.1146/annurev.cellbio.24.110707.175320.
- ↑ (en) Agrawal, GK (2010). Plant secretome: unlocking secrets of the secreted proteins.. Proteomics 10 (4): 799–827. PMID 19953550. DOI: 10.1002/pmic.200900514.
- Dit artikel of een eerdere versie ervan is een (gedeeltelijke) vertaling van het artikel Signal peptide op de Engelstalige Wikipedia, dat onder de licentie Creative Commons Naamsvermelding/Gelijk delen valt. Zie de bewerkingsgeschiedenis aldaar.
Externe links
- MeSH Signal+Peptide
- SignalP — voorspelt de aanwezigheid en locatie van signaalpeptidesplitsingsplaatsen in aminozuursequenties van verschillende organismen.