Bakteri

Bakteri (nama ilmiah: Bacteria) adalah kelompok mikroorganisme bersel satu yang diklasifikasikan pada tingkat domain. Bersama dengan domain Arkea, bakteri digolongkan sebagai prokariota[4]. Sel bakteri memiliki bentuk tertentu, misalnya menyerupai bola, batang, atau spiral, yang biasanya berukuran beberapa mikrometer. Bakteri merupakan salah satu bentuk kehidupan pertama yang muncul dan saat ini menghuni sebagian besar habitat di Bumi. Bakteri dapat hidup di tanah, air, mata air panas yang asam, limbah radioaktif, hingga kerak Bumi. Bakteri juga menjalin hubungan simbiosis dengan tumbuhan dan hewan. Sebagian besar bakteri belum diketahui karakternya, dan hanya sekitar 27 persen filum bakteri yang memiliki spesies yang dapat ditumbuhkan di laboratorium. Studi tentang bakteri disebut bakteriologi, salah satu cabang mikrobiologi.
Hampir semua hewan bergantung pada bakteri agar mereka dapat bertahan hidup karena hanya bakteri dan sejumlah arkea yang memiliki gen dan enzim yang diperlukan untuk menyintesis vitamin B12. Vitamin ini diperoleh hewan melalui rantai makanan atau dihasilkan oleh mikroorganisme yang hidup dalam sistem pencernaan mereka. Terdapat sekitar 40 juta sel bakteri dalam satu gram tanah dan satu juta sel bakteri dalam satu mililiter air tawar. Secara keseluruhan, ada sekitar 4–6 x 1030 bakteri dan arkea di Bumi, yang membentuk biomassa yang hanya dilampaui oleh tumbuhan. Bakteri sangat berperan dalam siklus nutrisi, misalnya dalam proses pengikatan nitrogen dari atmosfer dan dekomposisi mayat. Pada komunitas organisme di sekitar ventilasi hidrotermal dan ventilasi dingin, bakteri ekstremofil menyediakan nutrisi yang dibutuhkan untuk menopang kehidupan dengan mengubah senyawa terlarut, seperti hidrogen sulfida dan metana, menjadi energi.
Pada manusia dan sebagian besar hewan, bakteri paling banyak berada di saluran pencernaan. Kulit juga dihuni bakteri dalam jumlah besar. Mayoritas bakteri dalam tubuh tidak berbahaya karena tubuh dilindungi sistem imun. Di samping itu, banyak bakteri yang bermanfaat, terutama sebagai flora usus. Namun, beberapa spesies bakteri bersifat patogenik dan menyebabkan penyakit menular, antara lain kolera, sifilis, gonore, antraks, kusta, dan pes. Penyakit bakterial mematikan yang paling banyak ditemukan adalah infeksi saluran pernapasan. Tuberkulosis membunuh sekitar dua juta orang per tahun, yang kebanyakan terjadi di Afrika Sub-Sahara. Antibiotik digunakan untuk mengobati infeksi bakteri dan juga digunakan dalam pertanian, yang membuat resistansi antibiotik menjadi masalah yang terus berkembang. Di bidang perindustrian, bakteri berperan penting dalam pengolahan limbah dan penguraian tumpahan minyak, produksi keju dan yoghurt melalui fermentasi, pemurnian emas, paladium, tembaga, dan logam lainnya pada sektor pertambangan, serta dalam bioteknologi seperti pembuatan antibiotik dan bahan kimia lainnya.
Sejarah penemuan
[sunting | sunting sumber]
Bakteri merupakan organisme mikroskopik sehingga sulit dideteksi, terutama sebelum ditemukannya mikroskop. Organisme ini pertama kali diamati pada tahun 1676 oleh Antony van Leeuwenhoek, pedagang dan ilmuwan Belanda. Ia menggunakan mikroskop berlensa tunggal yang dirancangnya sendiri.[5] Leeuwenhoek lalu menerbitkan pengamatannya dalam serangkaian surat kepada Royal Society of London,[6][7][8] yang kemudian dipublikasikan dalam bahasa Inggris pada 1684.[9] Bakteri merupakan objek yang berada dalam batas yang bisa dilihat oleh lensa sederhana Leeuwenhoek dan tak ada orang lain yang bisa melihatnya selama lebih dari satu abad.[10] Leeuwenhoek juga mengamati protozoa, yang kesemuanya ia sebut sebagai "hewan kecil".[11]
Christian Gottfried Ehrenberg, ilmuwan Jerman, memperkenalkan kata bacterium pada tahun 1838.[12][13] Kata ini berasal dari romanisasi bahasa Yunani βακτηριον (bakterion),[14] bentuk diminutif dari βακτηρία (bakteria) yang memiliki arti "batang" atau "tongkat" karena bakteri pertama yang ditemukan berbentuk batang.[15][16]
Pada pertengahan abad ke-19, Ferdinand Cohn, seorang ahli botani asal Breslau, Prusia (sekarang bagian dari Polandia), tertarik pada bakteri yang tahan panas. Ia menemukan bahwa sejumlah bakteri membentuk endospora yang resistan terhadap suhu tinggi, termasuk Bacillus yang mampu beralih dari bentuk vegetatif menjadi endospora dan sebaliknya. Cohn juga menginisiasi pengelompokan bakteri berdasarkan bentuknya (bulat, batang, filamen, dan spiral) serta mengembangkan beberapa metode untuk mencegah kontaminasi pada kultur bakteri, seperti penggunaan kapas sebagai penutup pada tabung reaksi.[9][17]
Louis Pasteur, ahli kimia Prancis, menemukan bahwa pemanasan dapat membunuh atau menonaktifkan bakteri dan mikroorganisme lain pada anggur sehingga anggur tersebut tidak mudah rusak dan memiliki umur simpan yang lebih panjang.[18] Metode ini kemudian disebut pasteurisasi. Pada periode 1859 hingga 1864, Pasteur membantah konsep pembentukan spontan melalui eksperimen-eksperimennya yang kemudian diterima secara luas.[19] Bersama Robert Koch yang hidup sezaman dengannya, Pasteur adalah pendukung awal teori kuman penyakit.[20]

Pada masa itu, mikroorganisme telah diketahui menyebabkan penyakit menular. Namun, belum ada bukti definitif yang mendukung teori ini sampai Robert Koch, dokter berkebangsaan Jerman, berhasil mengisolasi dan membuat biakan murni bakteri, serta menumbuhkannya di laboratorium. Bacillus anthracis dan Mycobacterium tuberculosis adalah bakteri yang digunakan Koch untuk membuktikan teori kuman penyakit hingga ia diberikan penghargaan Nobel Fisiologi atau Kedokteran pada 1905.[21][22] Postulat Koch yang dirumuskannya untuk menentukan kausalitas antara patogen dan penyakit infeksi masih dipakai hingga saat ini.[23]
Meskipun berbagai penyakit bakterial telah diketahui, tetapi saat itu belum ada pengobatan yang memadai.[24] Sekitar tahun 1910, Paul Ehrlich bersama rekan-rekannya mengembangkan antibiotik sintetis pertama, yaitu Salvarsan (yang kemudian dikenal sebagai Arsfenamina) untuk mengobati sifilis yang diakibatkan oleh Treponema pallidum.[25] Ehrlich menerima penghargaan Nobel pada 1908 atas karyanya di bidang imunologi.[26] Ia juga memelopori penggunaan bahan pewarna untuk mendeteksi dan mengidentifikasi bakteri, yang menjadi dasar berbagai teknik pewarnaan seperti Ziehl–Neelsen.[27]
Perkembangan besar pada bakteriologi terjadi pada tahun 1977 ketika Carl Woese memublikasikan bahwa arkea memiliki garis keturunan evolusioner yang terpisah dari bakteri.[3] Taksonomi filogenetik ini bergantung pada pengurutan RNA ribosomal 16S dan membagi prokariota menjadi dua domain, sebagai bagian dari sistem tiga domain.[1]
Asal-usul dan evolusi awal
[sunting | sunting sumber]
Nenek moyang bakteri masa kini adalah mikroorganisme uniseluler yang merupakan bentuk kehidupan pertama di Bumi sekitar 4 miliar tahun yang lalu. Selama sekitar 3 miliar tahun, mayoritas organisme berukuran mikroskopis, yang didominasi oleh bakteri dan arkea.[28][29] Walaupun fosil bakteri ditemukan, misalnya dalam bentuk stromatolit, morfologinya yang tidak terlalu khas mengakibatkan mereka tak bisa digunakan untuk mengetahui riwayat evolusi bakteri atau waktu munculnya spesies bakteri tertentu. Meskipun demikian, urutan gen dapat digunakan untuk merekonstruksi filogeni bakteri, yang menunjukkan bahwa bakterilah yang pertama kali membentuk cabang dan keluar dari garis keturunan arkea/eukariota.[30] Nenek moyang bersama paling terkini dari bakteri dan arkea mungkin adalah hipertermofil yang hidup sekitar 2,5 hingga 3,2 miliar tahun yang lalu.[31][32] Bentuk kehidupan paling awal di darat mungkin berupa bakteri yang hidup sekitar 3,22 miliar tahun yang lalu.[33]
Bakteri juga terlibat dalam divergensi evolusioner besar kedua yang menciptakan percabangan arkea dan eukariota. Saat itu, eukariota terbentuk dari peristiwa masuknya bakteri purba ke dalam nenek moyang sel eukariota (yang mungkin masih berhubungan dekat dengan arkea) melalui asosiasi endosimbiotik.[34][35] Secara lebih spesifik, sel-sel proto-eukariota “menelan” Alphaproteobacteria sebagai simbion sehingga terbentuk salah satu dari mitokondria atau hidrogenosom, yang masih ditemukan di semua sel eukariota yang diketahui (kadang-kadang dalam bentuk yang sangat tereduksi, misalnya dalam protozoa kuno tanpa mitokondria). Belakangan, beberapa eukariota yang sudah memiliki mitokondria juga menelan organisme mirip sianobakteri yang pada akhirnya membentuk kloroplas pada alga dan tumbuhan. Hal ini dikenal sebagai endosimbiosis primer.[36][37]
Fosil
[sunting | sunting sumber]Satu kelompok bakteri, sianobakteri atau "blue green algae," telah meninggalkan fosil jauh dari zaman Prakambrium. Fosil cyanobacteria tertua yang diketahui hingga saat ini berusia hampir 3,5 miliar tahun. Sianobakteri lebih besar dari bakteri pada umumnya, dan dapat mengeluarkan dinding sel yang tebal. Selain itu, sianobakteri juga dapat membentuk struktur berlapis besar, yang disebut stromatolit (jika berbentuk kubah) atau onkolit (jika bulat). Struktur ini terbentuk sebagai alas sianobakteri yang tumbuh di lingkungan akuatik, membentuk sedimen dan terkadang mengeluarkan kalsium karbonat. Ketika dipotong sangat tipis, sianobakteri dan alga fosil yang terawetkan dengan baik dapat ditemukan pada fosil stromatolit.
Selain sianobakteri , tidak banyak fosil bakteri yang dipublikasikan. Dalam kondisi tertentu, sel bakteri dapat di salah pahamkan dengan mineral, terutama dengan pirit atau siderit (besi karbonat), mineral yang dapat membentuk replika dari sel hidup atau pseudomorphs. Beberapa bakteri juga mengeluarkan selubung berlapis besi yang terkadang memfosil. Adapula bakteri yang masuk ke dalam cangkang atau batu dan membentuk saluran mikroskopis di dalam cangkang; bakteri tersebut disebut sebagai endolitik. Bakteri juga telah ditemukan dalam damar, fosil resin pohon, dan dalam jaringan mumi.[38]
Morfologi
[sunting | sunting sumber]
Bakteri memiliki berbagai macam bentuk dan ukuran. Sel bakteri besarnya sekitar sepersepuluh sel eukariota dan biasanya berukuran 0,5 hingga 5 mikrometer. Namun, beberapa spesies bisa dilihat dengan mata telanjang, misalnya Thiomargarita namibiensis yang panjangnya mencapai setengah milimeter[39] dan Epulopiscium fishelsoni yang mencapai 0,7 mm.[40] Contoh bakteri terkecil adalah anggota genus Mycoplasma yang berukuran 0,3 mikrometer, kurang lebih sama dengan ukuran virus terbesar.[41] Beberapa bakteri bahkan mungkin lebih kecil, tetapi jenis-jenis bakteri ultramikro ini belum dipahami dengan baik.[42]
Sebagian besar spesies bakteri berbentuk bulat (disebut kokus; dari bahasa Yunani kókkos yang artinya butir atau biji) atau berbentuk batang (disebut basilus, dari bahasa Latin baculus yang artinya tongkat).[43] Beberapa jenis bakteri berbentuk seperti batang yang agak melengkung atau berbentuk koma (disebut vibrio); bakteri-bakteri lainnya bisa berbentuk spiral (disebut spirillum) atau melingkar rapat (disebut spiroket). Bentuk yang tidak umum juga telah dijumpai, misalnya bakteri berbentuk bintang.[44] Berbagai macam bentuk ini ditentukan oleh dinding sel bakteri dan sitoskeleton, yang berperan penting karena dapat memengaruhi kemampuan bakteri dalam memperoleh nutrisi, menempel pada permukaan, berenang dalam cairan, dan melarikan diri dari predator.[45][46]

Banyak spesies bakteri hanya berupa sel tunggal, sementara bakteri yang lain berkelompok dalam pola yang khas: Neisseria berbentuk diploid (berpasangan), Streptococcus membentuk rantai, sedangkan Staphylococcus bergerombol bersama-sama menyerupai sekumpulan anggur. Bakteri juga dapat berkelompok membentuk struktur multiseluler yang lebih besar, seperti Actinobacteria dengan filamen yang memanjang, miksobakteri yang membentuk agregat, dan Streptomyces yang mempunyai hifa kompleks.[47] Struktur-struktur multiseluler ini sering kali hanya terlihat pada kondisi tertentu. Sebagai contoh, ketika kekurangan asam amino, miksobakteri mendeteksi sel-sel di sekitarnya melalui proses yang dikenal sebagai pengindraan kuorum untuk bermigrasi menuju satu sama lain dan berkumpul membentuk tubuh buah dengan panjang hingga 500 mikrometer dan mengandung sekitar 100.000 sel bakteri.[48] Dalam tubuh buah ini, bakteri-bakteri melakukan tugas terpisah; misalnya, sekitar satu dari sepuluh sel bermigrasi ke bagian atas tubuh buah dan berdiferensiasi menjadi bentuk dorman khusus yang disebut miksospora yang lebih tahan terhadap kondisi kering dan keadaan lingkungan yang merugikan.[49]
Bakteri sering kali menempel pada suatu permukaan dan membentuk agregasi padat yang disebut biofilm, sementara formasi yang lebih besar dikenal sebagai tikar mikrob. Ketebalan biofilm dan tikar ini sekitar beberapa mikrometer sedangkan kedalamannya dapat mencapai setengah meter, dan mungkin mengandung banyak spesies bakteri, protista, dan arkea. Bakteri yang hidup dalam biofilm menampilkan susunan sel dan komponen ekstraseluler yang kompleks, serta membentuk struktur sekunder, seperti mikrokoloni, yang di dalamnya terdapat jejaring saluran untuk memungkinkan difusi nutrisi yang lebih baik.[50][51] Di lingkungan alami, seperti tanah atau permukaan tumbuhan, sebagian besar bakteri terikat dalam bentuk biofilm.[52] Biofilm merupakan hal penting dalam kedokteran karena struktur ini sering kali muncul saat infeksi bakteri berlangsung kronis atau saat terjadi infeksi pada implan peralatan medis. Bakteri yang terlindung dalam biofilm jauh lebih sulit dibunuh dibandingkan bakteri yang hidup sendiri-sendiri.[53]
Struktur sel
[sunting | sunting sumber]
Struktur intraseluler
[sunting | sunting sumber]Sel bakteri dikelilingi oleh membran sel, yang terutama terbuat dari fosfolipid. Membran ini membungkus isi sel dan menjadi pembatas bagi nutrien, protein, dan komponen-komponen penting lainnya di sitoplasma agar mereka tetap berada di dalam sel.[54] Tidak seperti eukariota, sel bakteri biasanya tidak memiliki struktur besar yang terbungkus membran di dalam sitoplasma mereka, seperti nukleus, mitokondria, kloroplas, dan organel-organel lainnya.[55] Meskipun demikian, sejumlah bakteri mempunyai organel yang berikatan dengan protein, contohnya karboksisom,[56] yang menciptakan kompartemen untuk memisahkan aspek-aspek metabolisme bakteri.[57][58] Selain itu, bakteri memiliki sitoskeleton multikomponen untuk mengatur lokalisasi protein dan asam nukleat di dalam sel, serta untuk mengelola proses pembelahan sel.[59][60][61]
Banyak reaksi biokimia esensial, seperti pembangkitan energi, terjadi karena adanya gradien konsentrasi lintas membran. Akibatnya, tercipta perbedaan potensial yang serupa dengan baterai. Secara umum, kurangnya jumlah membran internal pada bakteri mengakibatkan reaksi-reaksi ini, misalnya rantai transpor elektron, berlangsung melintasi membran sel, baik antara sitoplasma (di bagian dalam sel) dengan bagian luar sel ataupun dengan periplasma.[62] Namun, pada banyak bakteri fotosintetik, membran plasma sangat terlipat dan mengisi sebagian besar sel dengan lapisan-lapisan membran pengumpul cahaya.[63] Kompleks pengumpul cahaya ini dapat membentuk struktur yang ditutupi lipid yang disebut klorosom pada bakteri belerang hijau.[64]
Bakteri tidak memiliki nukleus yang terbungkus membran. Materi genetiknya biasanya berupa nukleoid, yaitu DNA yang terletak di sitoplasma secara ireguler yang membentuk kromosom melingkar tunggal.[65] Nukleoid mengandung kromosom yang lengkap dengan struktur protein dan RNA-nya. Seperti semua organisme lain, bakteri memiliki ribosom untuk menghasilkan protein, tetapi struktur ribosom bakteri berbeda dari ribosom pada eukariota dan arkea.[66]
Sejumlah bakteri menghasilkan butiran penyimpanan nutrisi di dalam selnya, seperti glikogen,[67] polifosfat,[68] belerang,[69] atau polihidroksi alkanoat.[70] Beberapa bakteri, seperti sianobakteri fotosintetik, mempunyai vakuola gas internal yang mereka gunakan untuk mengatur daya apung sehingga mereka dapat berpindah untuk naik atau turun di dalam lap air yang memiliki intensitas cahaya dan tingkat nutrisi yang berbeda.[71]
Struktur ekstraseluler
[sunting | sunting sumber]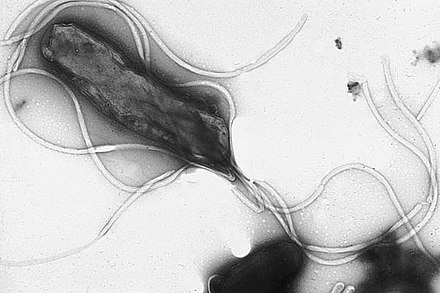
Lapisan yang mengelilingi bagian luar membran sel adalah dinding sel. Dinding sel bakteri terbuat dari peptidoglikan (disebut juga murein), yang disusun oleh rantai polisakarida yang terhubung secara silang dengan peptida yang mengandung asam amino-D.[72] Dinding sel bakteri berbeda dari dinding sel tumbuhan dan fungi, yang masing-masing terbuat dari selulosa dan kitin.[73] Dinding sel bakteri juga berbeda dengan arkea yang tidak mengandung peptidoglikan. Bagi banyak bakteri, dinding sel sangat penting untuk kelangsungan hidup mereka karena beberapa zat, misalnya penisilin (antibiotik yang diproduksi oleh jamur Penicillium), mampu membunuh bakteri dengan menghalangi satu langkah reaksi dalam sintesis peptidoglikan.[73]
Secara garis besar, ada dua jenis dinding sel pada bakteri, yang mengelompokkan bakteri menjadi bakteri Gram-positif dan bakteri Gram-negatif. Penamaan tersebut didasarkan dari reaksi sel terhadap pewarnaan Gram, suatu metode yang telah lama dilakukan untuk mengklasifikasikan jenis bakteri.[74]
Bakteri Gram-positif mempunyai dinding sel tebal yang mengandung banyak lapisan peptidoglikan dan asam teikoat. Sebaliknya, bakteri Gram-negatif memiliki dinding sel yang relatif tipis yang terdiri atas beberapa lapisan peptidoglikan yang dikelilingi oleh membran lipida dwilapis yang mengandung lipopolisakarida dan lipoprotein. Sebagian besar bakteri memiliki dinding sel bertipe Gram-negatif, dan hanya filum Firmicutes dan Actinobacteria (sebelumnya masing-masing dikenal sebagai bakteri Gram-positif dengan G+C rendah dan G+C tinggi) yang memiliki susunan Gram-positif alternatif.[75] Perbedaan struktur ini dapat menghasilkan perbedaan kerentanan terhadap antibiotik; misalnya, vankomisin hanya dapat membunuh bakteri Gram-positif dan tidak efektif melawan patogen Gram-negatif, seperti Haemophilus influenzae atau Pseudomonas aeruginosa.[76] Sebagian bakteri mempunyai struktur dinding sel yang tidak tergolong Gram-positif atau Gram-negatif, termasuk bakteri yang penting secara klinis seperti Mycobacterium yang mempunyai dinding sel dengan peptidoglikan tebal seperti bakteri Gram-positif, tetapi juga memiliki lapisan lipid kedua di bagian luarnya.[77]
Pada banyak bakteri, lapisan-S berupa molekul protein yang tersusun secara kaku menutupi bagian luar sel.[78] Lapisan ini melindungi permukaan sel secara fisik dan kimiawi dan dapat bertindak sebagai penghalang difusi makromolekul. Lapisan-S memiliki fungsi yang beragam, tetapi sebagian besar fungsinya kurang dipahami. Sejauh ini, lapisan-S diketahui bertindak sebagai faktor virulensi pada Campylobacter dan mengandung enzim permukaan pada Bacillus stearothermophilus.[79]

Banyak bakteri memiliki struktur ekstrasel lainnya seperti flagela, fimbria, dan pili yang digunakan untuk bergerak, melekat, dan berkonjugasi.[80] Flagela merupakan struktur protein kaku yang digunakan untuk motilitas. Diameter flagela sekitar 20 nanometer dan panjangnya mencapai 20 mikrometer. Flagela digerakkan oleh energi yang dilepaskan oleh transfer ion, yang terjadi karena gradien elektrokimia lintas membran sel.[81] Fimbria (kadang-kadang disebut "pili yang melekat") adalah filamen protein yang halus, dengan diameter sekitar 2–10 nanometer dan panjang beberapa mikrometer. Mereka tersebar di permukaan sel dan terlihat seperti rambut halus bila diamati melalui mikroskop elektron. Fimbria diyakini terlibat dalam perlekatan bakteri ke permukaan padat atau ke sel lain, dan berperan dalam virulensi beberapa bakteri patogen.[82] Sementara itu, pili adalah struktur pelengkap yang sedikit lebih besar dari fimbria. Struktur ini disebut sebagai pili konjugasi atau pili kelamin saat menjadi sarana transfer materi genetik antarsel bakteri dalam proses yang disebut konjugasi (lihat genetika bakteri di bawah).[83] Mereka juga dapat menghasilkan gerakan yang disebut pili tipe IV.[84]

Banyak bakteri memproduksi glikokaliks untuk mengelilingi sel mereka. Kompleksitas struktur glikokaliks bervariasi, mulai dari lapisan lendir tak teratur yang terbuat dari zat polimer ekstraseluler hingga kapsul yang sangat terstruktur. Struktur-struktur ini dapat melindungi sel bakteri dari sel eukariota, misalnya makrofag (bagian dari sistem imun manusia), yang hendak menelan mereka.[85] Glikokaliks juga memiliki beberapa peran lain: bertindak sebagai antigen, terlibat dalam pengenalan sel, serta membantu perlekatan ke suatu permukaan dan pembentukan biofilm.[86]
Perakitan struktur-struktur ekstraseluler bergantung pada sistem sekresi bakteri, yang mentransfer protein dari sitoplasma ke periplasma atau ke lingkungan di sekitar sel. Para ilmuwan telah mengetahui bermacam-macam sistem sekresi bakteri dan menemukan bahwa struktur-struktur ekstrasel yang dihasilkannya sering kali berperan penting dalam menentukan virulensi patogen. Oleh karenanya, mereka dipelajari secara intensif.[87]
Endospora
[sunting | sunting sumber]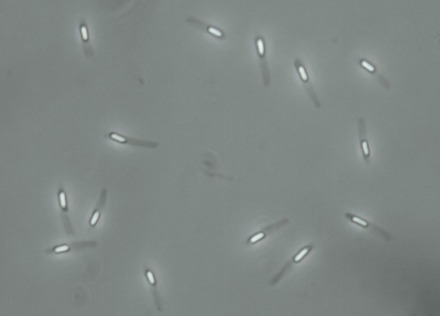
Beberapa genus bakteri Gram-positif, seperti Bacillus, Clostridium, Sporohalobacter, Anaerobacter, dan Heliobacterium, dapat membentuk struktur yang sangat resistan yang disebut endospora.[88] Endospora berkembang di dalam sitoplasma dan umumnya ada satu endospora yang berkembang di setiap sel. Setiap endospora mengandung DNA dan ribosom yang dikelilingi oleh lapisan korteks dan dilindungi oleh berlapis-lapis selubung kaku yang terdiri dari peptidoglikan dan berbagai protein.[89]
Endospora tidak menunjukkan tanda-tanda metabolisme dan dapat bertahan dari tekanan fisik dan kimia, seperti sinar ultraungu, radiasi gama, detergen, disinfektan, panas, pembekuan, tekanan, dan pengeringan, dalam tingkatan yang ekstrem.[90] Dalam keadaan yang tidak aktif ini, suatu organisme dapat tetap hidup selama jutaan tahun,[91][92] dan endospora bahkan memungkinkan bakteri bertahan hidup pada kondisi hampa udara dan radiasi di ruang angkasa sehingga mungkin bakteri dapat didistribusikan ke seluruh Alam semesta melalui debu kosmik, meteoroid, asteroid, komet, planetoid, atau melalui panspermia terarah.[93] Bakteri pembentuk endospora juga dapat menyebabkan penyakit. Sebagai contoh, antraks dapat ditularkan dengan menghirup endospora Bacillus anthracis, sementara luka-tusuk dalam yang terkontaminasi endospora Clostridium tetani dapat menyebabkan tetanus.[94] Selain itu, endospora Clostridium botulinum membuatnya terlindung dari suhu dan tekanan tinggi pada pemrosesan makanan kaleng sehingga dapat mengakibatkan keracunan saat dikonsumsi.[95]
Metabolisme
[sunting | sunting sumber]Bakteri menunjukkan tipe metabolisme yang sangat beragam.[96] Perbedaan sifat metabolik dalam suatu kelompok bakteri awalnya digunakan untuk menentukan taksonomi mereka, tetapi sifat-sifat ini sering kali tidak selaras dengan klasifikasi modern berbasis genetik.[97] Metabolisme bakteri dibagi menjadi beberapa kelompok nutrisi berdasarkan tiga kriteria utama: sumber energi, donor elektron yang digunakan, dan sumber karbon yang digunakan untuk pertumbuhan.[98]

Bakteri memperoleh energi dengan salah satu dari dua cara: berfotosintesis untuk mengubah energi dari cahaya (mereka disebut fototrof) atau dengan memecah senyawa kimia menggunakan oksidasi (disebut kemotrof).[99] Bakteri kemotrof menggunakan senyawa kimia sebagai sumber energi dengan mentransfer elektron dari donor ke akseptor terminal dalam reaksi redoks. Reaksi ini melepaskan energi yang dapat digunakan untuk bermetabolisme. Kemotrof selanjutnya dibagi berdasarkan jenis senyawa yang mereka gunakan untuk mentransfer elektron. Bakteri yang menggunakan senyawa anorganik seperti hidrogen, karbon monoksida, atau amonia sebagai sumber elektron disebut litotrof, sedangkan yang menggunakan senyawa organik disebut organotrof. Senyawa yang digunakan untuk menerima elektron juga digunakan untuk mengklasifikasikan bakteri: organisme aerob menggunakan oksigen sebagai akseptor elektron terminal, sedangkan organisme anaerob menggunakan senyawa lain seperti nitrat, sulfat, atau karbon dioksida.[99]
Banyak bakteri mendapatkan karbon untuk selnya dari karbon organik lain; mereka disebut heterotrof. Bakteri lainnya seperti sianobakteri dan beberapa bakteri ungu merupakan autotrof, artinya mereka memperoleh karbon dengan memfiksasi karbon dioksida.[100] Dalam situasi tertentu, gas metana dapat digunakan oleh bakteri metanotrof sebagai sumber elektron dan sebagai substrat untuk anabolisme karbon.[101]
| Tipe nutrisi | Sumber energi | Sumber karbon | Contoh |
|---|---|---|---|
| Fototrof | Cahaya matahari | Senyawa organik (fotoheterotrof) atau fiksasi karbon (fotoautotrof) | Sianobakteri, bakteri belerang hijau, Chloroflexi, dan bakteri ungu |
| Litotrof | Senyawa anorganik | Senyawa organik (litoheterotrof) atau fiksasi karbon (litoautotrof) | Thermodesulfobacteriaceae, Hydrogenophilaceae, dan Nitrospiraceae |
| Organotrof | Senyawa organik | Senyawa organik (kemoheterotrof) atau fiksasi karbon (kemoautotrof) | Bacillus, Clostridium, dan Enterobacteriaceae |
Dalam banyak hal, metabolisme bakteri memberi manfaat bagi stabilitas ekologi dan kehidupan manusia. Sebagai contoh, beberapa bakteri mampu memfiksasi gas nitrogen menggunakan enzim nitrogenase. Sifat ini penting bagi lingkungan dan dapat ditemukan pada sebagian besar tipe metabolisme bakteri yang disebutkan di atas,[102] yang mengarah pada proses denitrifikasi, reduksi sulfat, dan asetogenesis, yang semuanya penting secara ekologis.[103][104] Proses metabolisme bakteri juga berperan penting dalam pencemaran; misalnya, bakteri pereduksi sulfat sangat bertanggung jawab atas produksi bentuk merkuri yang sangat beracun (metilmerkuri dan dimetilmerkuri) di lingkungan.[105] Bakteri anaerob nonrespiratori menggunakan fermentasi untuk menghasilkan energi dan mengurangi daya, serta mengeluarkan produk sampingan metabolik (seperti etanol dalam pembuatan bir) sebagai limbah. Bakteri anaerob fakultatif dapat beralih antara fermentasi dan beberapa bentuk akseptor elektron terminal yang berbeda, tergantung pada kondisi lingkungan tempat mereka berada.[106]
Habitat
[sunting | sunting sumber]
Bakteri hidup di mana-mana dengan jumlah berlimpah. Ekosistem tempat bakteri hidup mencakup ekosistem terestrial, ekosistem akuatik, di dalam tubuh makhluk hidup lainnya, dan di struktur buatan manusia.[107] Menurut sebuah penelitian tahun 1998, jumlah bakteri dan arkea yang ada di Bumi diperkirakan sebanyak 4–6 x 1030, yang mayoritas hidup di biosfer dalam. Sekitar 3,5 x 1030 prokariota hidup di biosfer dalam di laut (lapisan dasar laut yang lebih dalam dari 10 cm) dan antara 0,25–2,5 x 1030 di biosfer dalam di terestrial (lapisan bawah tanah yang lebih dalam dari 8 m). Jumlah yang lebih rendah ditemukan di habitat-habitat terestrial, yaitu 2,6 x 1029 sel, dan di habitat-habitat akuatik, yaitu 1,2 x 1029 sel. Proporsi yang jauh lebih kecil ditemukan di dalam tubuh hewan (termasuk manusia), di daun dan bagian tumbuhan lain, serta di udara.[108] Kesemuanya membentuk biomassa terbesar di Bumi yang hanya dilampaui oleh tumbuhan.[109]
Di tanah, yang merupakan habitat penting bagi berbagai organisme, jumlah dan kepadatan bakteri berbeda-beda tergantung tipe ekosistemnya. Secara umum, jumlah bakteri tanah di ekosistem hutan lebih rendah dibandingkan tipe ekosistem lainnya, seperti gurun bersemak, sabana, dan lahan pertanian. Hingga kedalaman satu meter, jumlah prokariota diperkirakan 40 juta sel per gram tanah hutan, sedangkan di tipe ekosistem terestrial lainnya mencapai 2 miliar sel per gram tanah. Di lingkungan akuatik, kepadatan bakteri tertinggi ditemukan di sedimen pada ketebalan 0 hingga 10 cm di dasar laut, yakni 460 juta sel per ml.[108] Di sedimen dasar laut ini, bakteri mempunyai peran penting dalam siklus biogeokimia, misalnya pada siklus belerang yang diperankan oleh Desulfobulbaceae.[110] Di sekitar ventilasi hidrotermal dan ventilasi dingin, bakteri ekstremofil menyediakan nutrisi yang dibutuhkan untuk menopang kehidupan dengan mengubah senyawa terlarut, seperti hidrogen sulfida dan metana, menjadi energi melalui kemosintesis.[111][112] Sementara itu, konsentrasi prokariota yang lebih rendah ditemukan di ekosistem sungai, danau air tawar, dan danau garam yaitu 10 juta sel per ml, sedangkan kepadatan 5 juta sel per ml ada di perairan landas benua dan di laut lepas hingga kedalaman 200 m.[108]

Bakteri dapat ditemukan di dalam tubuh manusia, terutama di dalam saluran pencernaan. Banyak media populer dan tulisan ilmiah menyebutkan bahwa jumlah sel bakteri yang menghuni tubuh manusia sekitar 10 kali lipat lebih banyak dibandingkan jumlah sel manusianya sendiri, dengan perkiraan 100 triliun sel bakteri dan 10 triliun sel manusia.[114] Meskipun demikian, jumlah mereka sangat bergantung pada banyak hal, seperti usia, ukuran tubuh, lingkungan, hingga pangan yang dikonsumsi.[115] Di sisi lain, sebuah studi yang diterbitkan pada 2016 menemukan bahwa perbandingannya sekitar 1,3 sel bakteri untuk setiap sel manusia.[116]
Jumlah bakteri di saluran pencernaan manusia cukup bervariasi. Mereka paling banyak ditemukan di usus besar, yaitu sekitar 1011 sel per gram isi usus, sedangkan di usus halus sebanyak 108 sel per gram, dan di lambung yang asam jumlahnya 104 sel mikrob per gram.[117] Dari segi variasi, hanya dua filum utama yang menghuni tubuh manusia, yaitu Firmicutes dan Bacteroidetes. Selain mereka, tercatat pula filum Actinobacteria, Proteobacteria, Fusobacteria, dan Verrucomicrobia.[113][118] Contoh yang biasa ditemukan adalah bakteri asam laktat Lactobacillus acidophilus.[119] Jenis ini tergolong probiotik, yang dilaporkan bermanfaat bagi tubuh dan mencegah gangguan kesehatan seperti diare dan penyakit degeneratif.[120][121] Bakteri juga dapat ditemukan di permukaan kulit dan mulut. Di dalam mulut, bakteri dapat membentuk biofilm berupa plak yang mengakibatkan bau mulut.[122]
Pengaruh lingkungan
[sunting | sunting sumber]
Kondisi lingkungan dapat memacu maupun menghambat pertumbuhan dan reproduksi bakteri. Faktor-faktor lingkungan yang sangat memengaruhi kehidupan bakteri adalah suhu, pH, ketersediaan air, dan oksigen.[123] Meskipun demikian, serupa dengan arkea, bakteri mampu hidup di lingkungan yang tidak memungkinkan organisme lain untuk hidup, misalnya lingkungan yang terlalu panas atau terlalu dingin, terlalu asam atau basa, dan terlalu bergaram. Habitat-habitat yang ekstrem menuntut mikroorganisme mengembangkan kemampuan untuk bertahan hidup. Kelompok mikroorganisme ini disebut sebagai ekstremofil.[124]
Suhu sangat berkaitan dengan metabolisme. Seiring dengan naiknya suhu, reaksi enzimatik akan berlangsung lebih cepat hingga mencapai suhu optimum yang memungkinkan sel bakteri tumbuh dengan kecepatan tertinggi. Akan tetapi, setelah melewati suhu tertentu, protein dan komponen-komponen sel lainnya akan mengalami denaturasi sehingga sel akan mati.[123] Demikian pula bila suhu lingkungan berada di bawah batas toleransi sel, membran sel akan kehilangan wujud semicairnya sehingga tak bisa menjalankan fungsinya. Berdasarkan kisaran suhu aktivitasnya, bakteri dibagi menjadi empat golongan: psikrofili yang hidup di lingkungan bersuhu rendah dengan suhu optimum sekitar 4 °C, mesofili yang hidup di lingkungan bersuhu sedang dengan suhu optimum sekitar 40 °C, termofili yang hidup di lingkungan bersuhu tinggi dengan suhu optimum sekitar 60 °C, dan hipertermofili yang hidupnya di lingkungan bersuhu sangat tinggi dengan suhu optimum 88 °C dan bahkan di atasnya.[125] Sebagai contoh, Thermus aquaticus merupakan salah satu spesies bakteri termofili yang hidup di mata air panas.[126] Di Antarktika, sejumlah bakteri melakukan vetrifikasi untuk bertahan hidup pada suhu hingga −20 °C.[127]
Selain suhu, pertumbuhan dan reproduksi bakteri juga ditentukan oleh derajat keasaman (pH). Sebagian besar lingkungan alam memiliki pH antara 3 hingga 9 sehingga hampir semua bakteri hidup pada kisaran pH ini. Tiap jenis mikroorganisme memiliki kisaran pH optimum sendiri-sendiri, dengan rentang 2–3 unit pH.[128] Oleh karena itu, bakteri juga bisa dikelompokkan berdasarkan pH lingkungannya. Organisme neutrofili tumbuh di pH netral, sedangkan asidofili tumbuh di lingkungan asam dan alkalifili tumbuh di lingkungan basa.[129]
Pada umumnya bakteri memerlukan kelembaban relatif yang cukup tinggi, kira-kira 85%.[130] Kelembaban relatif dapat didefinisikan sebagai kandungan air yang terdapat di udara.[130] Pengurangan kadar air dari protoplasma menyebabkan kegiatan metabolisme terhenti, misalnya pada proses pembekuan dan pengeringan.[130] Sebagai contoh, bakteri Escherichia coli akan mengalami penurunan daya tahan dan elastisitas dinding selnya saat RH lingkungan kurang dari 84%.[131] Bakteri gram positif cenderung hidup pada kelembaban udara yang lebih tinggi dibandingkan dengan bakteri gram negatif terkait dengan perubahan struktur membran selnya yang mengandung lipid bilayer.[132]
Cahaya merupakan salah satu faktor yang mempengaruhi pertumbuhan bakteri.[133] Secara umum, bakteri dan mikroorganisme lainnya dapat hidup dengan baik pada paparan cahaya normal.[133] Akan tetapi, paparan cahaya dengan intensitas sinar ultraviolet (UV) tinggi dapat berakibat fatal bagi pertumbuhan bakteri.[133] Teknik penggunaan sinar UV, sinar x, dan sinar gamma untuk mensterilkan suatu lingkungan dari bakteri dan mikroorganisme lainnya dikenal dengan teknik iradiasi yang mulai berkembang sejak awal abad ke-20.[133][134] Metode ini telah diaplikasikan secara luas untuk berbagai keperluan, terutama pada sterilisasi makanan untuk meningkatkan masa simpan dan daya tahan.[134] Beberapa contoh bakteri patogen yang mampu dihambat ataupun dihilangkan antara lain Escherichia coli 0157:H7 dan Salmonella.[134]
Radiasi pada kekuatan tertentu dapat menyebabkan kelainan dan bahkan dapat bersifat letal bagi makhluk hidup, terutama bakteri.[135] Sebagai contoh pada manusia, radiasi dapat menyebabkan penyakit hati akut, katarak, hipertensi, dan bahkan kanker.[135] Akan tetapi, terdapat kelompok bakteri tertentu yang mampu bertahan dari paparan radiasi yang sangat tinggi, bahkan ratusan kali lebih besar dari daya tahan manusia tehadap radiasi, yaitu kelompok Deinococcaceae.[136] Sebagai perbandingan, manusia pada umumnya tidak dapat bertahan pada paparan radiasi lebih dari 10 Gray (Gy, 1 Gy = 100 rad), sedangkan bakteri yang termasuk dalam kelompok ini dapat bertahan hingga 5.000 Gy.[136][137]
Pada umumnya, paparan energi radiasi dapat menyebabkan mutasi gen dan putusnya rantai DNA.[138] Apabila terjadi pada intensitas yang tinggi, bakteri dapat mengalami kematian.[138] Deinococcus radiodurans memiliki kemampuan untuk bertahan terhadap mekanisme perusakan materi genetik tersebut melalui sistem adaptasi dan adanya proses perbaikan rantai DNA yang sangat efisien.[138]
Ada pula bakteri halofili yang dapat hidup di lingkungan dengan kadar garam yang sangat tinggi, seperti Salinibacter ruber yang tumbuh optimal pada konsentrasi garam antara 20 hingga 30%.[139] Selain itu, sejumlah bakteri lain yang mampu hidup pada kadar gula tinggi (kelompok osmofil), kadar air rendah (kelompok xerofil), serta derajat keasaman yang sangat tinggi dan rendah.[130]
Beberapa komunitas bakteri dapat bertahan hidup di dalam awan dengan ketingian hingga 10 kilometer. Sebuah tim peneliti menggunakan pesawat tua DC-8 yang dimodifikasi sebagai laboratorium terbang berhasil menggambil sampel sejumlah bakteri di awan dalam kondisi badai. Bakteri yang hidup dalam nukleasi es terbawa badai dan bertahan dalam ionisasi awan.[140]
Pertumbuhan dan reproduksi
[sunting | sunting sumber]
Pada organisme uniseluler, penambahan ukuran sel (pertumbuhan sel) dan reproduksi (melalui pembelahan sel) merupakan dua hal yang terkait erat. Bakteri tumbuh hingga mencapai ukuran yang tetap dan kemudian berkembang biak melalui pembelahan biner, salah satu bentuk reproduksi aseksual.[141] Dalam kondisi optimal, bakteri dapat tumbuh dan membelah dengan sangat cepat; populasi bakteri dapat bertambah dua kali lipat setiap 9,8 menit.[142] Pembelahan sel menghasilkan dua sel anakan yang identik. Meskipun masih bereproduksi secara aseksual, beberapa bakteri membentuk struktur reproduksi yang lebih kompleks untuk membantu menyebarkan sel anak yang baru terbentuk. Contohnya pembentukan tubuh buah oleh miksobakteri, pembentukan hifa oleh Streptomyces, serta pembentukan tunas. Pertunasan terjadi saat sebuah sel membentuk tonjolan yang kemudian memisahkan diri dari sel induk menjadi sel anak.
Di laboratorium, bakteri biasanya ditumbuhkan dengan menggunakan media padat atau media cair. Media pertumbuhan padat, seperti lempeng agar, digunakan untuk mengisolasi kultur murni suatu galur bakteri. Sementara itu, media pertumbuhan cair digunakan saat ilmuwan ingin mengukur pertumbuhan bakteri atau memerlukan sejumlah besar volume sel bakteri. Media cair dapat diaduk sampai terbentuk suspensi sel yang merata sehingga kultur bakteri mudah dibagi-bagi dan dipindahkan ke wadah lainnya. Penggunaan media selektif (media dengan penambahan atau pengurangan nutrisi tertentu atau dengan penambahan antibiotik) dapat membantu mengidentifikasi organisme spesifik.[144]
Sebagian besar teknik untuk menumbuhkan bakteri di laboratorium menggunakan nutrisi yang tinggi untuk menghasilkan sel dalam jumlah besar dengan murah dan cepat. Namun, di lingkungan alami, jumlah nutrisi terbatas. Artinya, bakteri tidak dapat terus berkembang biak selamanya. Keterbatasan nutrisi menyebabkan bakteri berevolusi dengan strategi pertumbuhan yang berbeda (lihat teori pemilihan r/K). Beberapa organisme dapat tumbuh sangat cepat ketika tersedia cukup nutrisi, seperti ledakan populasi alga (dan sianobakteri) yang sering terjadi di danau selama musim panas.[145] Bakteri lainnya beradaptasi terhadap lingkungan yang keras, seperti Streptomyces yang menghasilkan beberapa antibiotik untuk menghambat pertumbuhan mikroorganisme pesaing.[146] Di alam, banyak organisme yang hidup dalam komunitas (misalnya biofilm), yang memungkinkan peningkatan pasokan nutrisi dan perlindungan dari tekanan lingkungan.[52] Hubungan-hubungan ini menjadi penting bagi pertumbuhan organisme tertentu atau kelompok organisme tertentu (disebut sebagai sintrofi).[147]

Pertumbuhan bakteri terdiri atas empat fase. Ketika populasi bakteri pertama kali memasuki lingkungan bernutrisi tinggi yang memungkinkan pertumbuhan, mereka perlu beradaptasi dengan lingkungan barunya. Fase pertama adalah fase lamban. Pertumbuhan terjadi secara perlahan saat sel-sel bakteri beradaptasi dengan lingkungan kaya nutrisi dan bersiap untuk pertumbuhan cepat. Fase lamban memiliki tingkat biosintesis yang tinggi karena protein yang diperlukan untuk pertumbuhan diproduksi dengan cepat.[148][149] Fase pertumbuhan yang kedua adalah fase logaritmik, yang juga dikenal sebagai fase eksponensial. Fase log ditandai dengan pertumbuhan eksponensial yang cepat. Laju peningkatan jumlah sel selama fase ini dikenal sebagai laju pertumbuhan (k), sementara waktu yang dibutuhkan sel untuk menggandakan diri dikenal sebagai waktu pembentukan (g). Selama fase log, nutrisi dimetabolisme dengan kecepatan maksimum hingga salah satu nutrisi habis sehingga pertumbuhan mulai terbatas. Fase ketiga adalah fase stasioner atau fase diam akibat nutrisi yang terkuras. Sel-sel mengurangi aktivitas metaboliknya dan mengonsumsi protein internal sel yang nonesensial. Fase diam merupakan transisi dari kondisi pertumbuhan cepat ke kondisi yang menanggapi stres. Ada peningkatan ekspresi gen yang terlibat dalam perbaikan DNA, metabolisme antioksidan, dan transportasi nutrisi.[150] Fase terakhir adalah fase kematian saat bakteri kehabisan nutrisi dan mati.
Genetika
[sunting | sunting sumber]
Sebagian besar bakteri memiliki kromosom melingkar tunggal yang panjangnya dapat berkisar dari 160.000 pasangan basa (bp) pada bakteri endosimbiotik Carsonella ruddii,[151] hingga 12.200.000 pasangan basa (12,2 Mbp) pada bakteri penghuni tanah Sorangium cellulosum.[152] Ada banyak variasi bentuk kromosom bakteri, misalnya beberapa spesies Streptomyces dan Borrelia yang mempunyai satu kromosom linier,[153][154] sementara beberapa spesies Vibrio memiliki lebih dari satu kromosom.[155] Bakteri juga dapat mengandung plasmid, molekul DNA ekstrakromosomal kecil yang mungkin mengandung gen-gen yang berguna mengembangkan berbagai fungsi, seperti resistansi antibiotik, kemampuan metabolisme, atau faktor virulensi.[156]
Genom bakteri biasanya menyandi beberapa ratus hingga beberapa ribu gen. Gen-gen tersebut umumnya tersusun saling menyambung dalam satu bentangan DNA dan meskipun beberapa jenis intron ditemukan dalam genom bakteri, frekuensinya jauh lebih sedikit dibandingan dengan intron pada eukariota.[157]
Sebagai organisme aseksual, bakteri mewarisi salinan genom yang identik dengan sel induknya dan bersifat klonal. Walaupun begitu, semua bakteri dapat berevolusi dan mengalami perubahan DNA akibat rekombinasi genetik atau mutasi. Mutasi disebabkan oleh kesalahan yang terjadi selama replikasi DNA atau oleh paparan mutagen. Laju mutasi sangat bervariasi di antara berbagai spesies bakteri dan bahkan di antara klon yang berasal dari satu spesies bakteri.[158] Mutasi bisa muncul secara acak selama replikasi atau akibat “dorongan stres”, saat terjadi peningkatan laju mutasi pada gen-gen tertentu yang terlibat dalam situasi yang membatasi pertumbuhan.[159]

Sejumlah bakteri bisa melakukan transfer materi genetik antarsel melalui tiga cara utama. Pertama, bakteri dapat mengambil DNA eksogen dari lingkungan sekitarnya melalui proses yang disebut transformasi.[160] Secara alami, banyak bakteri yang memiliki kompetensi untuk mengambil DNA dari lingkungan, sementara bakteri lain harus dimodifikasi secara kimiawi untuk menginduksi mereka agar mengambil DNA.[161] Kemampuan ini dapat dikembangkan secara alami dan biasanya dikaitkan dengan kondisi lingkungan yang penuh tekanan, sebagai adaptasi bakteri untuk memperbaiki DNA yang rusak. Cara kedua untuk mentransfer materi genetik adalah dengan transduksi, ketika bakteriofag (suatu jenis virus) memasukkan DNA asing ke dalam kromosom bakteri. Ada banyak jenis bakteriofag; beberapa di antaranya hanya menginfeksi dan melisiskan bakteri inangnya, sementara yang lain masuk ke dalam kromosom bakteri.[162] Bakteri melawan infeksi fag melalui sistem modifikasi restriksi yang mendegradasi DNA asing[163] dan sistem yang menggunakan CRISPR yang memungkinkan mereka untuk memblokir replikasi virus melalui interferensi RNA.[164][165] Metode ketiga untuk mentransfer gen adalah konjugasi, saat DNA ditransfer melalui kontak antarsel secara langsung. Dalam keadaan biasa, transfer DNA melalui transduksi, konjugasi, dan transformasi melibatkan individu bakteri dari spesies yang sama, tetapi terkadang transfer DNA dapat terjadi antara individu bakteri yang spesiesnya berbeda dan hal ini dapat menimbulkan efek yang signifikan, seperti transfer kemampuan resistansi antibiotik.[166][167]
Perilaku
[sunting | sunting sumber]Pergerakan
[sunting | sunting sumber]
Banyak bakteri bersifat motil (dapat bergerak sendiri) dengan menggunakan berbagai mekanisme. Alat gerak yang paling dipelajari dengan baik adalah flagela, filamen panjang menyerupai cambuk yang pangkalnya menempel pada rotor yang berputar untuk menghasilkan gerakan seperti baling-baling.[168] Arah putaran flagela bersifat reversibel, yang menggunakan gradien elektrokimia lintas membran untuk menciptakan daya.[169]

Spesies bakteri yang berbeda memiliki jumlah dan susunan flagela yang berbeda. Ada spesies yang mempunyai flagela tunggal (disebut bakteri monotrik), ada juga yang memiliki flagela di setiap ujungnya (amfitrik), memiliki kelompok flagela di kutub sel (lofotrik), atau flagela yang terdistribusi di seluruh permukaan sel (peritrik). Flagela pada spiroket ditemukan di tempat yang unik, yaitu antara dua membran di ruang periplasmik. Kelompok bakteri ini memiliki tubuh heliks khas yang ikut berputar saat mereka bergerak.[168]
Bakteri motil mendekati atau menjauhi rangsangan tertentu. Perilaku ini disebut taksis, yang mencakup kemotaksis, fototaksis, energitaksis, dan magnetotaksis.[170][171][172] Pada satu kelompok khusus, miksobakteri, individu bakteri bergerak bersama-sama untuk membentuk gelombang sel yang kemudian berdiferensiasi membentuk tubuh buah yang mengandung spora.[49] Miksobakteri hanya bergerak pada permukaan padat, tidak seperti E. coli, yang bergerak dalam media cair atau padat.[173]
Sebagai patogen intraseluler, beberapa spesies Listeria dan Shigella bergerak di dalam sel inang dengan mengambil alih sitoskeleton, yang biasanya digunakan untuk memindahkan organel di dalam sel inang. Dengan mendorong polimerisasi aktin di salah satu kutub selnya, mereka dapat membentuk semacam ekor agar dapat bergerak di dalam sitoplasma sel inang.[174]
Komunikasi
[sunting | sunting sumber]Bentuk komunikasi antarbakteri dijumpai pada biofilm. Sel-sel bakteri yang membentuk agregat saling bertukar sinyal molekuler untuk berkomunikasi dan terlibat dalam perilaku multiseluler yang terkoordinasi. Manfaat dari kerja sama multiseluler di antaranya mengatur pembagian kerja seluler, mengakses sumber daya yang tidak dapat digunakan secara efektif oleh sel tunggal, secara kolektif bertahan melawan antagonis, dan mengoptimalkan kelangsungan hidup populasi dengan berdiferensiasi menjadi sel-sel yang berbeda jenisnya.[175] Sebagai contoh, sekumpulan bakteri dalam biofilm dapat meningkatkan resistansi terhadap agen antibakteri hingga lebih dari 500 kali lipat dibandingkan bakteri-bakteri planktonik (bakteri yang cenderung hidup bebas) yang spesiesnya seragam.[176]
Salah satu jenis komunikasi antarsel yang menggunakan sinyal molekuler adalah pengindraan kuorum. Mekanisme ini berfungsi untuk menentukan kepadatan lokal bakteri sejenis. Jika kepadatannya cukup tinggi, bakteri-bakteri tersebut akan bersama-sama melakukan hal yang sama, misalnya mengeluarkan enzim pencernaan atau memancarkan cahaya.[177][178]
Pada pengindraan kuorum, bakteri akan menghasilkan, melepaskan, dan mendeteksi sinyal molekuler. Jika sinyal molekulernya banyak (sebagai akibat dari banyaknya jumlah bakteri sejenis), mereka kemudian melakukan ekspresi gen yang terkoordinasi, yang akan menghasilkan protein dan menginisiasi perilaku yang serupa.[179]
Klasifikasi
[sunting | sunting sumber]
Taksonomi bertujuan menguraikan keanekaragaman spesies bakteri dengan cara memberi nama dan mengelompokkan bakteri berdasarkan kesamaan mereka. Bakteri dapat diklasifikasikan berdasarkan struktur sel, metabolisme sel, atau perbedaan komponen selnya, seperti DNA, asam lemak, pigmen, antigen, dan kuinon.[144] Meskipun metode ini memungkinkan identifikasi dan klasifikasi hingga galur bakteri, tetapi perbedaan yang ditemukan masih tidak jelas, apakah mewakili variasi di antara spesies yang berbeda atau di antara galur pada spesies yang sama. Ketidakpastian ini disebabkan oleh kurangnya struktur pembeda pada sebagian besar bakteri serta adanya transfer gen horizontal di antara spesies yang tidak terkait.[181] Akibat transfer gen horizontal, sejumlah bakteri yang berkerabat dekat dapat memiliki morfologi dan metabolisme yang sangat berbeda. Untuk mengatasi ketidakpastian ini, klasifikasi bakteri modern menggunakan filogenetika molekuler, yang memakai teknik genetik seperti penentuan rasio sitosina-guanina, hibridisasi genom-genom, serta pengurutan gen yang belum mengalami transfer gen horizontal secara ekstensif, seperti gen rRNA.[182] Klasifikasi bakteri ditentukan oleh publikasi dalam International Journal of Systematic and Evolutionary Microbiology dan Bergey's Manual.[183][184] Komite Internasional Sistematika Prokariota (ICSP) menetapkan aturan internasional mengenai penamaan bakteri, kategori taksonomi, dan peringkat mereka dalam Peraturan Internasional bagi Nomenklatur Prokariota.[185]
Pada awalnya, istilah "bakteri" digunakan untuk menyebut semua prokariota mikroskopis bersel tunggal. Namun, filogenetika molekuler menunjukkan bahwa prokariota terdiri dari dua domain terpisah, yang awalnya disebut Eubacteria dan Archaebacteria, tetapi sekarang disebut Bacteria dan Archaea; keduanya berevolusi secara independen dari satu nenek moyang bersama.[1] Hubungan kekerabatan antara arkea dan eukariota lebih dekat dibandingkan antara mereka dengan bakteri. Kesemuanya, yaitu Bacteria, Archaea, dan Eukarya, membentuk sistem tiga domain yang saat ini menjadi sistem klasifikasi yang paling banyak digunakan.[186] Namun, karena penerapan filogenetika molekuler masih relatif baru dan jumlah urutan genom yang diketahui meningkat dengan cepat, klasifikasi bakteri tetap menjadi bidang yang berubah dan berkembang.[187][188] Sebagai contoh, Cavalier-Smith, ahli biologi evolusioner, berpendapat bahwa arkea dan eukariota berevolusi dari bakteri Gram-positif.[189]
Identifikasi bakteri di laboratorium sangat relevan dalam ilmu kedokteran karena terapi yang tepat ditentukan oleh keakuratan identifikasi bakteri penyebab infeksi. Akibatnya, kebutuhan untuk mengidentifikasi patogen merupakan dorongan utama untuk mengembangkan teknik identifikasi bakteri.

Pewarnaan Gram, yang dikembangkan pada tahun 1884 oleh Hans Christian Gram, mengelompokkan bakteri berdasarkan struktur dinding selnya. Lapisan tebal peptidoglikan di dinding sel bakteri "Gram-positif" menunjukkan warna ungu, sedangkan dinding sel bakteri "Gram-negatif" yang tipis tampak merah muda. Dengan menggabungkan morfologi dan pewarnaan Gram, sebagian besar bakteri dapat diklasifikasikan sebagai salah satu dari empat kelompok: kokus Gram-positif, basil Gram-positif, kokus Gram-negatif, dan basil Gram-negatif. Sejumlah bakteri lebih baik diidentifikasi dengan pengecatan selain pewarnaan Gram, terutama Mycobacterium atau Nocardia, yang menunjukkan sifat tahan asam pada pewarnaan Ziehl–Neelsen atau pewarnaan serupa.[190] Bakteri lainnya mungkin perlu diidentifikasi dengan ditumbuhkan di media khusus, atau dengan teknik lain, seperti serologi.[191]

Kultur bakteri dirancang untuk mendorong pertumbuhan dan mengidentifikasi bakteri spesifik, sekaligus membatasi pertumbuhan bakteri lain dalam sampel. Jenis spesimen tertentu biasanya diperlakukan dengan teknik tertentu; misalnya, spesimen dahak akan ditangani untuk mengidentifikasi organisme penyebab pneumonia, sedangkan spesimen feses dikultur pada media selektif yang dapat mengidentifikasi organisme penyebab diare, sekaligus mencegah pertumbuhan bakteri nonpatogenik. Spesimen yang biasanya steril, seperti darah, urine, atau cairan serebrospinal, dikultur dalam kondisi yang dirancang untuk menumbuhkan semua kemungkinan mikroorganisme.[144][192] Setelah bakteri patogenik berhasil diisolasi, mereka dapat dikarakterisasi lebih lanjut berdasarkan morfologi, pola pertumbuhan (seperti pertumbuhan aerobik atau anaerobik), pola hemolisis, dan respons mereka terhadap pewarnaan.[193]
Seperti klasifikasi, teknik identifikasi bakteri juga semakin meningkat dengan adanya metode molekuler. Diagnosis menggunakan metode berbasis DNA, seperti reaksi berantai polimerase, semakin populer karena kecepatan dan spesifisitasnya dibandingkan metode berbasis kultur.[194] Metode molekuler juga memungkinkan deteksi dan identifikasi sel yang "dapat hidup tetapi tidak dapat dibiakkan", yaitu bakteri yang aktif secara metabolik tetapi tidak membelah.[195] Akan tetapi, bahkan dengan menggunakan metode-metode modern, jumlah spesies bakteri belum diketahui dan bahkan tidak dapat diperkirakan dengan pasti. Berdasarkan klasifikasi saat ini, jumlah spesies prokariota (bakteri dan arkea) yang diketahui hampir mencapai 9.300. Di sisi lain, studi yang memperkirakan keseluruhan jumlah keanekaragaman bakteri berkisar dari 107 hingga 109 spesies—dan bahkan perkiraan berinterval besar ini mungkin saja meleset jauh.[196][197]
Pohon filogenetika
[sunting | sunting sumber]Bagian ini memerlukan pengembangan. Anda dapat membantu dengan mengembangkannya. |
Interaksi dengan organisme lain
[sunting | sunting sumber]
Meskipun terlihat sederhana, bakteri dapat membentuk asosiasi yang kompleks dengan organisme lain. Asosiasi berupa simbiosis ini dapat dibedakan menjadi parasitisme, mutualisme, dan komensalisme. Karena ukurannya yang kecil, bakteri komensal ada di mana-mana, baik di permukaan maupun di dalam tubuh hewan dan tumbuhan, serta di permukaan benda lainnya. Pertumbuhan bakteri dapat ditingkatkan oleh suhu hangat dan keringat; pada manusia, populasi bakteri yang besar merupakan penyebab bau badan.[198]
Predator
[sunting | sunting sumber]Bakteri predator yaitu spesies bakteri yang membunuh dan kemudian memakan mikroorganisme lain.[199] Contohnya adalah Myxococcus xanthus yang membentuk kawanan sel yang membunuh dan mencerna bakteri yang mereka temui.[200] Predator bakteri lain mampu menempel pada mangsanya untuk mencerna dan menyerap nutrisi, seperti Vampirovibrio chlorellavorus,[201] atau menginvasi sel lain dan berkembang biak di dalam sitosolnya, seperti Daptobacter.[202] Bakteri predator diperkirakan telah berevolusi dari saprofag, yang memakan mikroorganisme mati, melalui adaptasi yang memungkinkan mereka untuk menjebak dan membunuh organisme lain.[203]
Mutualis
[sunting | sunting sumber]
Beberapa jenis bakteri membentuk hubungan erat dengan organisme lain untuk menjaga kelangsungan hidupnya. Salah satu simbiosis mutualisme ini disebut transfer hidrogen antarspesies, yang terjadi antara kelompok bakteri anaerob yang mengonsumsi asam organik (seperti asam butirat atau asam propionat) dan menghasilkan hidrogen dengan arkea metanogenik yang mengonsumsi hidrogen.[204] Bakteri-bakteri ini tidak dapat mengonsumsi asam organik dalam jumlah banyak karena reaksinya akan menghasilkan hidrogen yang terakumulasi di sekitarnya. Hanya dengan berhubungan dekat dengan arkea pengonsumsi hidrogen, konsentrasi hidrogen terjaga cukup rendah sehingga bakteri bisa tetap tumbuh.[205]
Di dalam tanah, mikroorganisme yang berada di rizosfer (zona yang meliputi permukaan akar dan tanah yang menempel pada akar tersebut setelah diguncang perlahan) melakukan fiksasi nitrogen, yang mengubah gas nitrogen menjadi senyawa yang mengandung nitrogen.[206] Reaksi ini berfungsi untuk menyediakan bentuk nitrogen yang mudah diserap bagi banyak tumbuhan yang tidak dapat mengikat nitrogen sendiri. Sementara itu, banyak bakteri lain ditemukan sebagai simbion pada manusia dan organisme lain. Misalnya, ada lebih dari 1.000 spesies bakteri sebagai flora normal pada usus manusia yang berperan dalam membentuk kekebalan usus, menyintesis vitamin seperti vitamin B7, vitamin B9, dan vitamin K, mengubah gula menjadi asam laktat (disebut bakteri asam laktat seperti Lactobacillus), serta memfermentasi karbohidrat kompleks yang tidak dapat dicerna manusia.[207][208][209] Kehadiran flora usus ini juga menghambat pertumbuhan bakteri yang berpotensi menjadi patogen (biasanya melalui eksklusi kompetitif) sehingga bakteri-bakteri bermanfaat ini dijual sebagai suplemen makanan dalam bentuk probiotik.[210]
Patogen
[sunting | sunting sumber]
Jika bakteri membentuk simbiosis parasitisme dengan organisme lain, mereka digolongkan sebagai patogen. Bakteri patogenik merupakan salah satu penyebab penyakit infeksi seperti tetanus (disebabkan oleh Clostridium tetani), demam tifoid (Salmonella serovar Typhi), difteri (Corynebacterium diphtheriae), sifilis (Treponema pallidum), kolera (Vibrio cholerae), kusta (Mycobacterium leprae), dan tuberkulosis (Mycobacterium tuberculosis), hingga beragam penyakit bawaan makanan. Suatu penyakit mungkin saja diketahui patogen penyebabnya setelah bertahun-tahun kemudian, seperti kasus penyakit tukak lambung dan Helicobacter pylori. Penyakit bakterial juga penting dalam pertanian dan peternakan, misalnya bakteri penyebab penyakit bercak daun, hawar api, dan kelayuan pada tumbuhan, serta paratuberkulosis, mastitis, dan antraks pada hewan ternak.[212]
Setiap spesies patogen memiliki spektrum interaksi yang khas dengan inangnya. Bakteri dalam genus Staphylococcus dan Streptococcus dapat mengakibatkan infeksi kulit, pneumonia, meningitis, dan sepsis, yaitu respons peradangan sistemik yang dapat berujung pada syok, vasodilasi masif, dan kematian.[213] Namun, mereka juga merupakan bagian dari flora normal manusia dan biasanya ada di kulit atau di hidung tanpa menyebabkan penyakit sama sekali. Bakteri lain selalu menyebabkan penyakit pada manusia, seperti Rickettsia yang merupakan parasit intraseluler obligat yang mampu tumbuh dan berkembang biak hanya di dalam sel organisme lain. Satu spesies Rickettsia menyebabkan tifus, sementara spesies yang lain menyebabkan demam berbintik Pegunungan Rocky. Chlamydia, patogen intraseluler obligat lainnya, mengandung spesies yang dapat menyebabkan pneumonia dan infeksi saluran kemih, serta mungkin terlibat dalam penyakit jantung koroner.[214] Beberapa spesies lainnya, seperti Pseudomonas aeruginosa, Burkholderia cenocepacia, dan Mycobacterium avium, merupakan patogen oportunistik yang menyebabkan penyakit terutama pada orang yang menderita imunosupresi atau fibrosis sistik.[215][216]
Infeksi bakteri dapat diobati dengan antibiotik, yang digolongkan sebagai bakterisida jika mereka membunuh bakteri atau bakteriostatik jika mereka hanya mencegah pertumbuhan bakteri. Ada banyak jenis antibiotik yang dikelompokkan menjadi beberapa golongan. Setiap golongan antibiotik mempunyai aksi yang berbeda dalam mencegah proses kimiawi tertentu dalam sel bakteri yang ditemukan dalam inang. Contoh antibiotik yang menghasilkan toksisitas selektif adalah kloramfenikol dan puromisina. Kedua golongan antibiotik ini menghambat ribosom bakteri, tetapi bukan ribosom eukariota yang strukturnya berbeda.[217] Antibiotik digunakan untuk mengobati penyakit manusia dan untuk mempercepat pertumbuhan hewan di peternakan intensif, yang mungkin berkontribusi pada cepatnya perkembangan resistansi antibiotik dalam populasi bakteri.[218] Infeksi bakteri dapat dicegah dengan tindakan antiseptik, misalnya mensterilkan kulit sebelum menusuknya dengan jarum suntik dan menjaga sterilitas kateter yang akan dimasukkan ke dalam tubuh. Peralatan bedah dan kedokteran gigi juga disterilkan untuk mencegah kontaminasi bakteri. Disinfektan seperti zat pemutih digunakan untuk membunuh bakteri atau patogen lain pada permukaan benda untuk mencegah kontaminasi dan mengurangi risiko infeksi.[219]
Peranan
[sunting | sunting sumber]Bidang lingkungan
[sunting | sunting sumber]Keanekaragaman bakteri dan jalur metabolismenya menyebabkan bakteri memiliki peranan yang besar bagi lingkungan.[134] Sebagai contoh, bakteri saprofit menguraikan tumbuhan atau hewan yang telah mati dan sisa-sisa atau kotoran organisme.[134] Bakteri tersebut menguraikan protein, karbohidrat dan senyawa organik lain menjadi CO2, gas amoniak, dan senyawa-senyawa lain yang lebih sederhana.[134] Contoh bakteri saprofit antara lain Proteus dan Clostridium.[134] Tidak hanya berperan sebagai pengurai senyawa organik, beberapa kelompok bakteri saprofit juga merupakan patogen oportunis.[134]

Kelompok bakteri lainnya berperan dalam siklus nitrogen, seperti bakteri nitrifikasi.[130] Bakteri nitrifikasi adalah kelompok bakteri yang mampu menyusun senyawa nitrat dari senyawa amonia yang pada umumnya berlangsung secara aerob di dalam tanah.[220] Kelompok bakteri ini bersifat kemolitotrof.[220] Nitrifikasi terdiri atas dua tahap yaitu nitritasi (oksidasi amonia (NH4) menjadi nitrit (NO2-)) dan nitratasi (oksidasi senyawa nitrit menjadi nitrat (NO3)).[220] Dalam bidang pertanian, nitrifikasi sangat menguntungkan karena menghasilkan senyawa yang diperlukan oleh tanaman yaitu nitrat.[220] Setelah reaksi nitrifikasi selesai, akan terjadi proses dinitrifikasi yang dilakukan oleh bakteri denitrifikasi.[220] Denitrifikasi sendiri merupakan reduksi anaerobik senyawa nitrat menjadi nitrogen bebas (N2) yang lebih mudah diserap dan dimetabolisme oleh berbagai makhluk hidup.[130] Contoh bakteri yang mampu melakukan metabolisme ini adalah Pseudomonas stutzeri, Pseudomonas aeruginosa, and Paracoccus denitrificans.[221] Di samping itu, reaksi ini juga menghasilkan nitrogen dalam bentuk lain, seperti dinitrogen oksida (N2O).[130] Senyawa tersebut tidak hanya dapat berperan penting bagi hidup berbagai organisme, tetapi juga dapat berperan dalam fenomena hujan asam dan rusaknya ozon.[130] Senyawa N2O akan dioksidasi menjadi senyawa NO dan selanjutnya bereaksi dengan ozon (O3) membentuk NO2- yang akan kembali ke bumi dalam bentuk hujan asam (HNO2).[130]
Di bidang pertanian dikenal adanya suatu kelompok bakteri yang mampu bersimbiosis dengan akar tanaman atau hidup bebas di tanah untuk membantu penyuburan tanah.[134] Kelompok bakteri ini dikenal dengan istilah bakteri pengikat nitrogen atau singkatnya bakteri nitrogen. Bakteri nitrogen adalah kelompok bakteri yang mampu mengikat nitrogen (terutaman N2) bebas di udara dan mereduksinya menjadi senyawa amonia (NH4) dan ion nitrat (NO3-) oleh bantuan enzim nitrogenase.[222][223] Kelompok bakteri ini biasanya bersimbiosis dengan tanaman kacang-kacangan dan polong untuk membentuk suatu simbiosis mutualisme berupa nodul atau bintil akar untuk mengikat nitrogen bebas di udara yang pada umumnya tidak dapat digunakan secara langsung oleh kebanyakan organisme.[130][223] Secara umum, kelompok bakteri ini dikenal dengan istilah rhizobia, termasuk di dalamnya genus bakteri Rhizobium, Bradyrhizobium, Mesorhizobium, Photorhizobium, dan Sinorhizobium.[130] Contoh bakteri nitrogen yang hidup bersimbiosis dengan tanaman polong-polongan yaitu Rhizobium leguminosarum, yang hidup di akar membentuk nodul atau bintil-bintil akar.[130]
Bidang pangan
[sunting | sunting sumber]Terdapat beberapa kelompok bakteri yang mampu melakukan proses fermentasi dan hal ini telah banyak diterapkan pada pengolahan berbagi jenis makanan.[134] Bahan pangan yang telah difermentasi pada umumnya akan memiliki masa simpan yang lebih lama, juga dapat meningkatkan atau bahkan memberikan cita rasa baru dan unik pada makanan tersebut.[134] Beberapa makanan hasil fermentasi dan mikroorganisme yang berperan:
| No. | Nama produk atau makanan | Bahan baku | Bakteri yang berperan |
|---|---|---|---|
| 1. | Yoghurt | susu | Lactobacillus bulgaricus dan Streptococcus thermophilus |
| 2. | Mentega | susu | Streptococcus lactis |
| 3. | Terasi | ikan | Lactobacillus sp. |
| 4. | Asinan buah-buahan | buah-buahan | Lactobacillus sp. |
| 5. | Sosis | daging | Pediococcus cerevisiae |
| 6. | Kefir | susu | Lactobacillus bulgaricus dan Streptococcus lactis |
Beberapa spesies bakteri pengurai dan patogen dapat tumbuh di dalam makanan.[224] Kelompok bakteri ini mampu memetabolisme berbagai komponen di dalam makanan dan kemudian menghasilkan metabolit sampingan yang bersifat racun.[224] Clostridium botulinum, menghasilkan racun botulinin, sering kali terdapat pada makanan kalengan dan kini senyawa tersebut dipakai sebagai bahan dasar botox.[224] Beberapa contoh bakteri perusak makanan:
- Burkholderia gladioli (sin. Pseudomonas cocovenenans), menghasilkan asam bongkrek, terdapat pada tempe bongkrek[225]
- Leuconostoc mesenteroides, penyebab pelendiran makanan, penurunan pH, dan pembentukkan gas.[226]
Bakteri juga dapat menyebabkan penyakit pada tanaman. Ralstonia solanacearum merupakan salah satu bakteri penyebab layu pada tanaman tomat. Tanaman yang terserang menunjukkan gejala layu mendadak bahkan dapat menimbulkan kematian. [227] Salah satu penyakit yang menyerang tanaman anggrek yaitu busuk busuk lunak yang disebabkan oleh bakteri Erwinia carotovora. Dalam perkembangan patogennya, gejala yang ditimbulkan akan cepat meluas dan dapat mematikan titik tumbuh tanaman. [228] Xanthomonas oryzae pv. oryzae (Swings et al. 1990) adalah bakteri patogen tanaman yang menyebabkan penyakit hawar daun pada padi, yang juga dikenal dengan sebutan penyakit kresek. [229] Penyakit busuk pangkal batang pada tanaman kedelai oleh Sclerotium rolsfii dapat menyebabkan rendahnya produksi kedelai. Penyakit ini sering ditemukan pada tanaman kedelai baik lahan kering, tadah hujan maupun pasang surut dengan intensitas serangan sebesar 5 - 55%. Tingkat serangan lebih dari 5% di lapang sudah dapat merugikan secara ekonomi. [230] Fusarium oxysporum f.sp. cubense (Foc) menyebabkan layu fusarium pada tanaman pisang. Infeksinya akan mengganggu proses penyerapan, transportasi air dan zat makanan di dalam tanah, sehingga tanaman menjadi layu dan akhirnya mati. [231]
Bidang kesehatan
[sunting | sunting sumber]Tidak hanya di bidang lingkungan dan pangan, bakteri juga dapat memberikan manfaat dibidang kesehatan. Antibiotik merupakan zat yang dihasilkan oleh mikroorganisme dan mempunyai daya hambat terhadap kegiatan mikroorganisme lain dan senyawa ini banyak digunakan dalam menyembuhkan suatu penyakit.[134] Beberapa bakteri yang menghasilkan antibiotik adalah:
- Streptomyces griseus, menghasilkan antibiotik streptomisin[130]
- Streptomyces aureofaciens, menghasilkan antibiotik tetrasiklin[130]
- Streptomyces venezuelae, menghasilkan antibiotik kloramfenikol[130]
- Penicillium, menghasilkan antibiotik penisilin[134]
- Bacillus polymyxa, menghasilkan antibiotik polimiksin.[134]
Terlepas dari peranannya dalam menghasilkan antibiotik, banyak jenis bakteri yang justru bersifat patogen.[232] Pada manusia, beberapa jenis bakteri yang sering kali menjadi agen penyebab penyakit adalah Salmonella enterica subspesies I serovar Typhi yang menyebabkan penyakit tifus, Mycobacterium tuberculosis yang menyebabkan penyakit TBC, dan Clostridium tetani yang menyebabkan penyakit tetanus.[232][233] Bakteri patogen juga dapat menyerang hewan ternak, seperti Brucella abortus yang menyebabkan brucellosis pada sapi dan Bacillus anthracis yang menyebabkan antraks.[234] Untuk infeksi pada tanaman yang umum dikenal adalah Xanthomonas oryzae yang menyerang pucuk batang padi dan Erwinia amylovora yang menyebabkan busuk pada buah-buahan.[235]
Dekomposisi
[sunting | sunting sumber]Proses degradasi jasad makhluk hidup dilakukan oleh banyak organisme, salah satunya adalah bakteri. Beberapa jenis bakteri, terutama bakteri heterotrof, mampu mendegradasi senyawa organik dan menggunakannya untuk menunjang pertumbuhannya.[236] Proses dekomposisi ini dibantu oleh beberapa jenis enzim untuk memecah makromolekul, seperti karbohidrat, protein, dan lemak, untuk dipecah menjadi senyawa yang lebih sederhana. Sebagai contoh, enzim protease digunakan untuk memecah protein menjadi senyawa lebih sederhana, seperti asam amino.[236] Proses dekomposisi ini juga berperan dalam pengembalian unsur-unsur, terutama karbon dan nitrogen, ke alam untuk masuk ke dalam siklus lagi.[237]
Dekomposisi jasad makhluk hidup dimulai oleh bakteri yang hidup di dalam tubuh manusia, dimulai dari jaringan-jaringan otot.[237] Proses ini dipercepat saat tubuh telah dikuburkan. Reaksi pertama dalam dekomposisi ini adalah hidrolisis protein oleh protease membentuk asam amino.[237] Selanjutnya, asam amino akan diubah menjadi asam asetat, gas hidrogen, gas nitrogen, dan karbon dioksida sehingga pH lingkungan akan turun menjadi 4-5.[237] Reaksi ini dilakukan oleh bakteri acetogen. Pada tahap akhir, semua senyawa tersebut diubah menjadi gas metana oleh metanogen.[237]
Bidang penelitian
[sunting | sunting sumber]Karena tingkat pertumbuhannya yang cepat dan manipulasi yang relatif mudah, bakteri memainkan peran penting dalam bidang biologi molekuler, genetika, dan biokimia. Dengan memperkenalkan mutasi ke dalam DNA bakteri dan memeriksa perubahan fenotip yang dihasilkan, para ilmuwan dapat mengungkap fungsi gen, enzim, dan jalur metabolisme pada bakteri. Pengetahuan ini kemudian dapat diekstrapolasi ke organisme yang lebih rumit.
Tujuan akhir dari memahami biokimia seluler dicontohkan dengan pembuatan model matematika yang komprehensif untuk seluruh organisme, yang menggabungkan data ekstensif tentang kinetika enzim dan ekspresi gen. Dalam kasus bakteri yang dipelajari dengan baik, seperti Escherichia coli, model-model ini saat ini sedang dalam pengembangan dan eksperimen.
Pemahaman mendalam tentang genetika dan metabolisme bakteri ini mendasari bidang bioteknologi, yang memungkinkan bioteknologi bakteri untuk memproduksi protein terapeutik seperti insulin, faktor pertumbuhan, dan antibodi.
Referensi
[sunting | sunting sumber]- ^ a b c Woese, C.R.; Kandler, O.; Wheelis, M.L. (1 Juni 1990). "Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya". Proceedings of the National Academy of Sciences. 87 (12): 4576–4579. doi:10.1073/pnas.87.12.4576. ISSN 0027-8424. PMC 54159
. PMID 2112744.
- ^ Pavan, María Elisa; Pavan, Esteban E.; Glaeser, Stefanie P.; Etchebehere, Claudia; Kämpfer, Peter; Pettinari, María Julia; López, Nancy I. (1 Mei 2018). "Proposal for a new classification of a deep branching bacterial phylogenetic lineage: transfer of Coprothermobacter proteolyticus and Coprothermobacter platensis to Coprothermobacteraceae fam. nov., within Coprothermobacterales ord. nov., Coprothermobacteria classis nov. and Coprothermobacterota phyl. nov. and emended description of the family Thermodesulfobiaceae". International Journal of Systematic and Evolutionary Microbiology. 68 (5): 1627–1632. doi:10.1099/ijsem.0.002720. ISSN 1466-5026. Diarsipkan dari versi asli tanggal 2021-05-24. Diakses tanggal 2021-05-24.
- ^ a b Woese, C.R.; Fox, G.E. (1 November 1977). "Phylogenetic structure of the prokaryotic domain: The primary kingdoms". Proceedings of the National Academy of Sciences. 74 (11): 5088–5090. doi:10.1073/pnas.74.11.5088. ISSN 0027-8424. PMC 432104
- ^ Parker, Sybil, P (1984). McGraw-Hill Dictionary of Biology. McGraw-Hill Company.
- ^ Porter, J.R. (Juni 1976). "Antony van Leeuwenhoek: tercentenary of his discovery of bacteria". Bacteriological Reviews. 40 (2): 260–269. doi:10.1128/mmbr.40.2.260-269.1976. PMC 413956
- ^ van Leeuwenhoek, A. (1684). "An abstract of a letter from Mr. Anthony Leevvenhoek at Delft, dated Sep. 17, 1683, Containing Some Microscopical Observations, about Animals in the Scurf of the Teeth, the Substance Call'd Worms in the Nose, the Cuticula Consisting of Scales". Philosophical Transactions. 14 (155–166): 568–574. Bibcode:1684RSPT...14..568L. doi:10.1098/rstl.1684.0030
- ^ van Leeuwenhoek, A. (1700). "Part of a Letter from Mr Antony van Leeuwenhoek, concerning the Worms in Sheeps Livers, Gnats, and Animalcula in the Excrements of Frogs". Philosophical Transactions. 22 (260–276): 509–18. Bibcode:1700RSPT...22..509V. doi:10.1098/rstl.1700.0013
- ^ van Leeuwenhoek, A. (1702). "Part of a Letter from Mr Antony van Leeuwenhoek, F.R.S. concerning Green Weeds Growing in Water, and Some Animalcula Found about Them". Philosophical Transactions. 23 (277–288): 1304–11. Bibcode:1702RSPT...23.1304V. doi:10.1098/rstl.1702.0042.
- ^ a b Madigan dkk. 2015, hlm. 13.
- ^ Asimov, Isaac (1982). Asimov's Biographical Encyclopedia of Science and Technology (edisi ke-2). Garden City, NY: Doubleday and Company. hlm. 143.
- ^ Lane, Nick (19 April 2015). "The unseen world: reflections on Leeuwenhoek (1677) 'Concerning little animals'". Philosophical Transactions of the Royal Society B: Biological Sciences. 370 (1666): 20140344. doi:10.1098/rstb.2014.0344. ISSN 0962-8436. PMC 4360124
- ^ Ehrenberg, Christian Gottfried; Carlgren, Oskar; Hemprich, W. F.; Hilgendorf, Franz; Klug, Fr.; Klug, Fr.; Martens, Eduard von; Matschie, Paul; Schumann, K. (1828). "Symbolae Physicae: Animalia Evertebrata Exclusis Insectis". Symbolae physicae, seu, Icones et descriptiones corporum naturalium novorum aut minus cognitorum : quae ex itineribus per Libyam Aegyptum Nubiam Dongalam Syriam Arabiam et Habessiniam publico institutis sumptu Friderici Guilelmi Hemprich et Christiani Godofredi Ehrenberg : studio annis MDCCCXX-MDCCCXXV redierunt. Berolini: Ex Officina Academica, venditur a Mittlero. hlm. 16. doi:10.5962/bhl.title.107403. Diarsipkan dari versi asli tanggal 2021-02-27. Diakses tanggal 2021-05-15.
- ^ Breed, Robert S.; Conn, H. J. (1936). "The Status of the Generic Term Bacterium Ehrenberg 1828*". Journal of Bacteriology. 31 (5): 517–518. doi:10.1128/JB.31.5.517-518.1936. ISSN 0021-9193. Diarsipkan dari versi asli tanggal 2021-05-15. Diakses tanggal 2021-05-15.
- ^ Liddell, Henry George; Scott, Robert (1940). "βακτήριον". A Greek–English Lexicon. Perseus Project. Oxford: Clarendon Press. Diarsipkan dari versi asli tanggal 2021-05-13. Diakses tanggal 2021-05-15.
- ^ "Bacterium". Lexico. Diarsipkan dari versi asli tanggal 2021-05-15. Diakses tanggal 15 Mei 2021.
- ^ Harper, Douglas. "Bacteria". Online Etymology Dictionary.
- ^ Drews, Gerhart (1999). "Ferdinand Cohn, a founder of modern microbiology" (PDF). ASM News. 65 (8): 547–552. Diarsipkan dari versi asli (PDF) tanggal 13 Juli 2017.
- ^ Vallery-Radot, René (1919). The Life of Pasteur. Diterjemahkan oleh Devonshire, R.L. London: Constable & Company. hlm. 113.
- ^ Roll-Hansen, Nils (1979). "Experimental Method and Spontaneous Generation: The Controversy between Pasteur and Pouchet, 1859–64" (PDF). Journal of the History of Medicine and Allied Sciences. XXXIV (3): 273–292. doi:10.1093/jhmas/XXXIV.3.273. PMID 383780. Diarsipkan dari versi asli (PDF) tanggal 3 Maret 2019.
- ^ "Pasteur's Papers on the Germ Theory". Louisiana State University Law Center's Medical and Public Health Law Site, Historic Public Health Articles. Diarsipkan dari versi asli tanggal 18 Desember 2006. Diakses tanggal 23 November 2006.
- ^ Madigan dkk. 2015, hlm. 16–20.
- ^ "The Nobel Prize in Physiology or Medicine 1905". Nobel Prize. Diarsipkan dari versi asli tanggal 10 Desember 2006. Diakses tanggal 22 November 2006.
- ^ Inglis TJ (November 2007). "Principia aetiologica: taking causality beyond Koch's postulates". Journal of Medical Microbiology. 56 (Pt 11): 1419–1422. doi:10.1099/jmm.0.47179-0
- ^ Thurston AJ (Desember 2000). "Of blood, inflammation and gunshot wounds: the history of the control of sepsis". The Australian and New Zealand Journal of Surgery. 70 (12): 855–861. doi:10.1046/j.1440-1622.2000.01983.x. PMID 11167573.
- ^ Schwartz RS (Maret 2004). "Paul Ehrlich's magic bullets". The New England Journal of Medicine. 350 (11): 1079–1080. doi:10.1056/NEJMp048021. PMID 15014180.
- ^ "Biography of Paul Ehrlich". Nobel Prize. Diarsipkan dari versi asli tanggal 28 November 2006. Diakses tanggal 26 November 2006.
- ^ Titford, Michael (Agustus 2010). "Paul Ehrlich: Histological Staining, Immunology, Chemotherapy". Laboratory Medicine. 41 (8): 497–498. doi:10.1309/LMHJS86N5ICBIBWM. ISSN 0007-5027.
- ^ Schopf, J.W. (Juli 1994). "Disparate rates, differing fates: tempo and mode of evolution changed from the Precambrian to the Phanerozoic". Proceedings of the National Academy of Sciences of the United States of America. 91 (15): 6735–6742. Bibcode:1994PNAS...91.6735S. doi:10.1073/pnas.91.15.6735. PMC 44277
- ^ DeLong, E.F.; Pace, N.R. (Agustus 2001). "Environmental diversity of bacteria and archaea". Systematic Biology. 50 (4): 470–478. CiteSeerX 10.1.1.321.8828
- ^ Brown, J.R.; Doolittle, W.F. (Desember 1997). "Archaea and the prokaryote-to-eukaryote transition". Microbiology and Molecular Biology Reviews. 61 (4): 456–502. doi:10.1128/.61.4.456-502.1997. PMC 232621
- ^ Di Giulio, M. (Desember 2003). "The universal ancestor and the ancestor of bacteria were hyperthermophiles". Journal of Molecular Evolution. 57 (6): 721–730. Bibcode:2003JMolE..57..721D. doi:10.1007/s00239-003-2522-6. PMID 14745541.
- ^ Battistuzzi, F.U.; Feijao, A.; Hedges, S.B. (November 2004). "A genomic timescale of prokaryote evolution: insights into the origin of methanogenesis, phototrophy, and the colonization of land". BMC Evolutionary Biology. 4: 44. doi:10.1186/1471-2148-4-44. PMC 533871
- ^ Homann, Martin; dkk. (23 Juli 2018). "Microbial life and biogeochemical cycling on land 3,220 million years ago" (PDF). Nature Geoscience. 11 (9): 665–671. Bibcode:2018NatGe..11..665H. doi:10.1038/s41561-018-0190-9. Diarsipkan (PDF) dari versi asli tanggal 2021-05-09. Diakses tanggal 2021-05-17.
- ^ Poole, A.M.; Penny, D. (Januari 2007). "Evaluating hypotheses for the origin of eukaryotes". BioEssays. 29 (1): 74–84. doi:10.1002/bies.20516. PMID 17187354.
- ^ Dyall, S.D.; Brown, M.T.; Johnson, P.J. (April 2004). "Ancient invasions: from endosymbionts to organelles". Science. 304 (5668): 253–257. Bibcode:2004Sci...304..253D. doi:10.1126/science.1094884. PMID 15073369.
- ^ Lang, B.F.; Gray, M.W.; Burger, G. (1999). "Mitochondrial genome evolution and the origin of eukaryotes". Annual Review of Genetics. 33: 351–397. doi:10.1146/annurev.genet.33.1.351. PMID 10690412.
- ^ McFadden, G.I. (Desember 1999). "Endosymbiosis and evolution of the plant cell". Current Opinion in Plant Biology. 2 (6): 513–519. doi:10.1016/S1369-5266(99)00025-4. PMID 10607659.
- ^ "Fossil Record of the Bacteria". ucmp.berkeley.edu. Diarsipkan dari versi asli tanggal 2022-01-17. Diakses tanggal 2022-02-21.
- ^ Schulz, Heide N.; Jørgensen, Bo Barker (Oktober 2001). "Big Bacteria". Annual Review of Microbiology. 55 (1): 105–137. doi:10.1146/annurev.micro.55.1.105. ISSN 0066-4227. Diarsipkan dari versi asli tanggal 2020-02-22. Diakses tanggal 2021-05-18.
- ^ Williams, Caroline (2011). "Who are you calling simple?". New Scientist. 211 (2821): 38–41. doi:10.1016/S0262-4079(11)61709-0.
- ^ Robertson, J.; Gomersall, M.; Gill, P. (November 1975). "Mycoplasma hominis: growth, reproduction, and isolation of small viable cells". Journal of Bacteriology. 124 (2): 1007–1018. doi:10.1128/JB.124.2.1007-1018.1975. PMC 235991
- ^ Velimirov, Branko (2001). "Nanobacteria, Ultramicrobacteria and Starvation Forms: A Search for the Smallest Metabolizing Bacterium". Microbes and environments. 16 (2): 67–77. doi:10.1264/jsme2.2001.67. ISSN 1342-6311. Diarsipkan dari versi asli tanggal 2021-05-18. Diakses tanggal 2021-05-18.
- ^ Dusenbery, David B. (2009). Living at Micro Scale: The Unexpected Physics of Being Small. Cambridge, Mass.: Harvard University Press. hlm. 20–25. ISBN 978-0-674-03116-6. OCLC 225874255. Diarsipkan dari versi asli tanggal 2020-06-09. Diakses tanggal 2021-05-18.
- ^ Yang, D.C.; Blair, K.M.; Salama, N.R. (Maret 2016). "Staying in Shape: the Impact of Cell Shape on Bacterial Survival in Diverse Environments". Microbiology and Molecular Biology Reviews. 80 (1): 187–203. doi:10.1128/MMBR.00031-15. PMC 4771367
- ^ Cabeen, Matthew T.; Jacobs-Wagner, Christine (Agustus 2005). "Bacterial cell shape". Nature Reviews Microbiology. 3 (8): 601–610. doi:10.1038/nrmicro1205. ISSN 1740-1526. Diarsipkan dari versi asli tanggal 2021-03-16. Diakses tanggal 2021-05-18.
- ^ Young, K.D. (September 2006). "The selective value of bacterial shape". Microbiology and Molecular Biology Reviews. 70 (3): 660–703. doi:10.1128/MMBR.00001-06. PMC 1594593
- ^ Claessen, Dennis; Rozen, Daniel E.; Kuipers, Oscar P.; Søgaard-Andersen, Lotte; van Wezel, Gilles P. (Februari 2014). "Bacterial solutions to multicellularity: a tale of biofilms, filaments and fruiting bodies". Nature Reviews Microbiology. 12 (2): 115–124. doi:10.1038/nrmicro3178. ISSN 1740-1526. Diarsipkan dari versi asli tanggal 2021-03-28. Diakses tanggal 2021-05-18.
- ^ Shimkets, Lawrence J. (Oktober 1999). "Intercellular Signaling During Fruiting-Body Development of Myxococcus xanthus". Annual Review of Microbiology. 53 (1): 525–549. doi:10.1146/annurev.micro.53.1.525. ISSN 0066-4227. Diarsipkan dari versi asli tanggal 2020-03-17. Diakses tanggal 2021-05-18.
- ^ a b Kaiser, Dale (Oktober 2004). "Signaling in myxobacteria". Annual Review of Microbiology. 58 (1): 75–98. doi:10.1146/annurev.micro.58.030603.123620. ISSN 0066-4227. Diarsipkan dari versi asli tanggal 2020-03-10. Diakses tanggal 2021-05-18.
- ^ Donlan, Rodney M. (September 2002). "Biofilms: Microbial Life on Surfaces". Emerging Infectious Diseases. 8 (9): 881–890. doi:10.3201/eid0809.020063. ISSN 1080-6040. PMC 2732559
- ^ Branda, Steven S.; Vik, Åshild; Friedman, Lisa; Kolter, Roberto (Januari 2005). "Biofilms: the matrix revisited". Trends in Microbiology. 13 (1): 20–26. doi:10.1016/j.tim.2004.11.006. Diarsipkan dari versi asli tanggal 2021-05-25. Diakses tanggal 2021-05-18.
- ^ a b Davey, Mary Ellen; O'toole, George A. (1 Desember 2000). "Microbial Biofilms: from Ecology to Molecular Genetics". Microbiology and Molecular Biology Reviews. 64 (4): 847–867. doi:10.1128/MMBR.64.4.847-867.2000. ISSN 1098-5557. PMC 99016
- ^ Donlan, Rodney M.; Costerton, J. William (April 2002). "Biofilms: Survival Mechanisms of Clinically Relevant Microorganisms". Clinical Microbiology Reviews. 15 (2): 167–193. doi:10.1128/CMR.15.2.167-193.2002. ISSN 0893-8512. PMC 118068
- ^ Slonczewski, Joan; Foster, John Watkins (2014). Microbiology: An Evolving Science (edisi ke-3). New York: John Watkins Foster. hlm. 82. ISBN 978-0-393-91929-5. OCLC 881060733.
- ^ Lodish, Harvey F.; Berk, Arnold; Kaiser, Chris A. (2013). Molecular Cell Biology (edisi ke-7). New York: W.H. Freeman and Co. hlm. 13. ISBN 978-1-4292-3413-9. OCLC 171110915. Diarsipkan dari versi asli tanggal 2019-12-18. Diakses tanggal 2021-05-21.
- ^ Kerfeld, C. A. (5 Agustus 2005). "Protein Structures Forming the Shell of Primitive Bacterial Organelles". Science. 309 (5736): 936–938. doi:10.1126/science.1113397. ISSN 0036-8075.
- ^ Bobik, Thomas A. (Mei 2006). "Polyhedral organelles compartmenting bacterial metabolic processes". Applied Microbiology and Biotechnology. 70 (5): 517–525. doi:10.1007/s00253-005-0295-0. ISSN 0175-7598.
- ^ Yeates, Todd O.; Kerfeld, Cheryl A.; Heinhorst, Sabine; Cannon, Gordon C.; Shively, Jessup M. (September 2008). "Protein-based organelles in bacteria: carboxysomes and related microcompartments". Nature Reviews Microbiology. 6 (9): 681–691. doi:10.1038/nrmicro1913. ISSN 1740-1526. Diarsipkan dari versi asli tanggal 2021-05-18. Diakses tanggal 2021-05-21.
- ^ Gitai, Zemer (Maret 2005). "The New Bacterial Cell Biology: Moving Parts and Subcellular Architecture". Cell. 120 (5): 577–586. doi:10.1016/j.cell.2005.02.026. Diarsipkan dari versi asli tanggal 2021-03-27. Diakses tanggal 2021-05-21.
- ^ Shih, Yu-Ling; Rothfield, Lawrence (September 2006). "The Bacterial Cytoskeleton". Microbiology and Molecular Biology Reviews. 70 (3): 729–754. doi:10.1128/MMBR.00017-06. ISSN 1092-2172. PMC 1594594
- ^ Norris, Vic; den Blaauwen, Tanneke; Cabin-Flaman, Armelle; Doi, Roy H.; Harshey, Rasika; Janniere, Laurent; Jimenez-Sanchez, Alfonso; Jin, Ding Jun; Levin, Petra Anne (Maret 2007). "Functional Taxonomy of Bacterial Hyperstructures". Microbiology and Molecular Biology Reviews. 71 (1): 230–253. doi:10.1128/MMBR.00035-06. ISSN 1092-2172. PMC 1847379
- ^ Harold, F M (1972). "Conservation and transformation of energy by bacterial membranes". Bacteriological Reviews. 36 (2): 172–230. doi:10.1128/BR.36.2.172-230.1972. ISSN 0005-3678. Diarsipkan dari versi asli tanggal 2021-05-21. Diakses tanggal 2021-05-21.
- ^ Bryant, Donald A.; Frigaard, Niels-Ulrik (November 2006). "Prokaryotic photosynthesis and phototrophy illuminated". Trends in Microbiology. 14 (11): 488–496. doi:10.1016/j.tim.2006.09.001. Diarsipkan dari versi asli tanggal 2021-06-03. Diakses tanggal 2021-05-21.
- ^ Pšenčík, J.; Ikonen, T.P.; Laurinmäki, P.; Merckel, M.C.; Butcher, S.J.; Serimaa, R.E.; Tuma, R. (Agustus 2004). "Lamellar Organization of Pigments in Chlorosomes, the Light Harvesting Complexes of Green Photosynthetic Bacteria". Biophysical Journal. 87 (2): 1165–1172. doi:10.1529/biophysj.104.040956. PMC 1304455
- ^ Thanbichler, Martin; Wang, Sherry C.; Shapiro, Lucy (15 Oktober 2005). "The bacterial nucleoid: A highly organized and dynamic structure". Journal of Cellular Biochemistry. 96 (3): 506–521. doi:10.1002/jcb.20519. ISSN 0730-2312.
- ^ Poehlsgaard, Jacob; Douthwaite, Stephen (November 2005). "The bacterial ribosome as a target for antibiotics". Nature Reviews Microbiology. 3 (11): 870–881. doi:10.1038/nrmicro1265. ISSN 1740-1526. Diarsipkan dari versi asli tanggal 2021-05-17. Diakses tanggal 2021-05-21.
- ^ Yeo, Marcus; Chater, Keith (1 Maret 2005). "The interplay of glycogen metabolism and differentiation provides an insight into the developmental biology of Streptomyces coelicolor". Microbiology. 151 (3): 855–861. doi:10.1099/mic.0.27428-0. ISSN 1350-0872. Diarsipkan dari versi asli tanggal 2021-04-15. Diakses tanggal 2021-05-21.
- ^ Shiba, T.; Tsutsumi, K.; Ishige, K.; Noguchi, T. (Maret 2000). "Inorganic polyphosphate and polyphosphate kinase: their novel biological functions and applications". Biochemistry. Biokhimiia. 65 (3): 315–323. ISSN 0006-2979. PMID 10739474. Diarsipkan dari versi asli tanggal 2021-03-04. Diakses tanggal 2021-05-21.
- ^ Brune, Daniel C. (Juni 1995). "Isolation and characterization of sulfur globule proteins from Chromatium vinosum and Thiocapsa roseopersicina". Archives of Microbiology. 163 (6): 391–399. doi:10.1007/BF00272127. ISSN 0302-8933.
- ^ Kadouri, Daniel; Jurkevitch, Edouard; Okon, Yaacov; Castro-Sowinski, Susana (Januari 2005). "Ecological and Agricultural Significance of Bacterial Polyhydroxyalkanoates". Critical Reviews in Microbiology. 31 (2): 55–67. doi:10.1080/10408410590899228. ISSN 1040-841X. Diarsipkan dari versi asli tanggal 2021-03-09. Diakses tanggal 2021-05-21.
- ^ Walsby, A E (1994). "Gas vesicles". Microbiological Reviews. 58 (1): 94–144. doi:10.1128/MR.58.1.94-144.1994. ISSN 0146-0749. Diarsipkan dari versi asli tanggal 2021-05-21. Diakses tanggal 2021-05-21.
- ^ Heijenoort, J. v. (1 Maret 2001). "Formation of the glycan chains in the synthesis of bacterial peptidoglycan". Glycobiology. 11 (3): 25R–36R. doi:10.1093/glycob/11.3.25R. ISSN 0959-6658. Diarsipkan dari versi asli tanggal 2022-05-11. Diakses tanggal 2021-05-21.
- ^ a b Koch, Arthur L. (Oktober 2003). "Bacterial Wall as Target for Attack". Clinical Microbiology Reviews. 16 (4): 673–687. doi:10.1128/CMR.16.4.673-687.2003. ISSN 0893-8512. PMC 207114
- ^ Gram, HC (1884). "Über die isolierte Färbung der Schizomyceten in Schnitt- und Trockenpräparaten". Fortschr. Med. 2: 185–189.
- ^ Hugenholtz, Philip (29 Januari 2002). "Exploring prokaryotic diversity in the genomic era". Genome Biology. 3 (2): reviews0003.1. doi:10.1186/gb-2002-3-2-reviews0003. PMC 139013
- ^ Walsh, Fiona M; Amyes, Sebastian GB (Oktober 2004). "Microbiology and drug resistance mechanisms of fully resistant pathogens". Current Opinion in Microbiology. 7 (5): 439–444. doi:10.1016/j.mib.2004.08.007. Diarsipkan dari versi asli tanggal 2021-05-31. Diakses tanggal 2021-05-21.
- ^ Alderwick, Luke J.; Harrison, James; Lloyd, Georgina S.; Birch, Helen L. (Agustus 2015). "The Mycobacterial Cell Wall—Peptidoglycan and Arabinogalactan". Cold Spring Harbor Perspectives in Medicine. 5 (8): a021113. doi:10.1101/cshperspect.a021113. ISSN 2157-1422. PMC 4526729
- ^ Engelhardt, Harald; Peters, Jürgen (Desember 1998). "Structural Research on Surface Layers: A Focus on Stability, Surface Layer Homology Domains, and Surface Layer–Cell Wall Interactions". Journal of Structural Biology. 124 (2-3): 276–302. doi:10.1006/jsbi.1998.4070. Diarsipkan dari versi asli tanggal 2021-02-25. Diakses tanggal 2021-05-21.
- ^ Beveridge, T. J.; Pouwels, P. H.; Sára, M.; Kotiranta, A.; Lounatmaa, K.; Kari, K.; Kerosuo, E.; Haapasalo, M.; Egelseer, E. M. (Juni 1997). "Functions of S-layers". FEMS microbiology reviews. 20 (1-2): 99–149. doi:10.1111/j.1574-6976.1997.tb00305.x. ISSN 0168-6445. PMID 9276929. Diarsipkan dari versi asli tanggal 2021-02-25. Diakses tanggal 2021-05-21.
- ^ Carl. The Bacteria Cell. http://www.lanesville.k12.in.us/lcsyellowpages/tickit/carl/bacteria.html[pranala nonaktif permanen]. Diakses pada 22 Juni 2011.
- ^ Kojima, Seiji; Blair, David F (2004). International Review of Cytology. 233. Elsevier. hlm. 93–134. doi:10.1016/s0074-7696(04)33003-2. ISBN 978-0-12-364637-8. Diarsipkan dari versi asli tanggal 2021-05-25. Diakses tanggal 2021-05-21.
- ^ Beachey, E. H. (1 Maret 1981). "Bacterial Adherence: Adhesin-Receptor Interactions Mediating the Attachment of Bacteria to Mucosal Surfaces". Journal of Infectious Diseases. 143 (3): 325–345. doi:10.1093/infdis/143.3.325. ISSN 0022-1899. Diarsipkan dari versi asli tanggal 2022-03-13. Diakses tanggal 2021-05-21.
- ^ Silverman, Philip M. (Januari 1997). "Towards a structural biology of bacterial conjugation". Molecular Microbiology. 23 (3): 423–429. doi:10.1046/j.1365-2958.1997.2411604.x.
- ^ Costa, Tiago R. D.; Felisberto-Rodrigues, Catarina; Meir, Amit; Prevost, Marie S.; Redzej, Adam; Trokter, Martina; Waksman, Gabriel (Juni 2015). "Secretion systems in Gram-negative bacteria: structural and mechanistic insights". Nature Reviews Microbiology. 13 (6): 343–359. doi:10.1038/nrmicro3456. ISSN 1740-1526. Diarsipkan dari versi asli tanggal 2021-03-10. Diakses tanggal 2021-05-21.
- ^ Stokes, Richard W.; Norris-Jones, Raymond; Brooks, Donald E.; Beveridge, Terry J.; Doxsee, Dan; Thorson, Lisa M. (Oktober 2004). "The Glycan-Rich Outer Layer of the Cell Wall of Mycobacterium tuberculosis Acts as an Antiphagocytic Capsule Limiting the Association of the Bacterium with Macrophages". Infection and Immunity. 72 (10): 5676–5686. doi:10.1128/IAI.72.10.5676-5686.2004. ISSN 0019-9567. PMC 517526
- ^ Daffé, M.; Etienne, G. (Juni 1999). "The capsule of Mycobacterium tuberculosis and its implications for pathogenicity". Tubercle and Lung Disease. 79 (3): 153–169. doi:10.1054/tuld.1998.0200. Diarsipkan dari versi asli tanggal 2021-03-31. Diakses tanggal 2021-05-23.
- ^ Finlay, B B; Falkow, S (1997). "Common themes in microbial pathogenicity revisited". Microbiology and molecular biology reviews : MMBR. 61 (2): 136–169. doi:10.1128/.61.2.136-169.1997. ISSN 1092-2172. Diarsipkan dari versi asli tanggal 2021-05-25. Diakses tanggal 2021-05-23.
- ^ Nicholson, Wayne L.; Munakata, Nobuo; Horneck, Gerda; Melosh, Henry J.; Setlow, Peter (1 September 2000). "Resistance of Bacillus Endospores to Extreme Terrestrial and Extraterrestrial Environments". Microbiology and Molecular Biology Reviews. 64 (3): 548–572. doi:10.1128/MMBR.64.3.548-572.2000. ISSN 1098-5557. PMC 99004
- ^ McKenney, Peter T.; Driks, Adam; Eichenberger, Patrick (Januari 2013). "The Bacillus subtilis endospore: assembly and functions of the multilayered coat". Nature Reviews Microbiology. 11 (1): 33–44. doi:10.1038/nrmicro2921. ISSN 1740-1526. Diarsipkan dari versi asli tanggal 2021-05-17. Diakses tanggal 2021-05-23.
- ^ Nicholson, Wayne L.; Fajardo-Cavazos, Patricia; Rebeil, Roberto; Slieman, Tony A.; Riesenman, Paul J.; Law, Jocelyn F.; Xue, Yaming (2002). "Bacterial endospores and their significance in stress resistance". Antonie van Leeuwenhoek. 81 (1/4): 27–32. doi:10.1023/A:1020561122764.
- ^ Vreeland, Russell H.; Rosenzweig, William D.; Powers, Dennis W. (Oktober 2000). "Isolation of a 250 million-year-old halotolerant bacterium from a primary salt crystal". Nature. 407 (6806): 897–900. doi:10.1038/35038060. ISSN 0028-0836. Diarsipkan dari versi asli tanggal 2021-06-05. Diakses tanggal 2021-05-23.
- ^ Cano, R.; Borucki, M. (19 Mei 1995). "Revival and identification of bacterial spores in 25- to 40-million-year-old Dominican amber". Science. 268 (5213): 1060–1064. doi:10.1126/science.7538699. ISSN 0036-8075.
- ^ Nicholson, Wayne L.; Schuerger, Andrew C.; Setlow, Peter (1 April 2005). "The solar UV environment and bacterial spore UV resistance: considerations for Earth-to-Mars transport by natural processes and human spaceflight". Mutation Research/Fundamental and Molecular Mechanisms of Mutagenesis. 571 (1-2): 249–264. doi:10.1016/j.mrfmmm.2004.10.012. Diarsipkan dari versi asli tanggal 2021-03-08. Diakses tanggal 2021-05-23.
- ^ Hatheway, C L (Januari 1990). "Toxigenic clostridia". Clinical Microbiology Reviews. 3 (1): 66–98. doi:10.1128/CMR.3.1.66. ISSN 0893-8512. PMC 358141
- ^ Margosch, Dirk; Ehrmann, Matthias A.; Buckow, Roman; Heinz, Volker; Vogel, Rudi F.; Ganzle, Michael G. (Mei 2006). "High-Pressure-Mediated Survival of Clostridium botulinum and Bacillus amyloliquefaciens Endospores at High Temperature". Applied and Environmental Microbiology. 72 (5): 3476–3481. doi:10.1128/AEM.72.5.3476-3481.2006. ISSN 0099-2240. PMC 1472378
- ^ Nealson, Kenneth H. (1999). "Post-Viking Microbiology: New Approaches, New Data, New Insights". Origins of Life and Evolution of the Biosphere. 29 (1): 73–93. doi:10.1023/A:1006515817767.
- ^ Xu, Jianping (8 Maret 2006). "Microbial ecology in the age of genomics and metagenomics: concepts, tools, and recent advances". Molecular Ecology. 15 (7): 1713–1731. doi:10.1111/j.1365-294X.2006.02882.x.
- ^ Zillig, Wolfram (Desember 1991). "Comparative biochemistry of Archaea and Bacteria". Current Opinion in Genetics & Development. 1 (4): 544–551. doi:10.1016/S0959-437X(05)80206-0. Diarsipkan dari versi asli tanggal 2021-04-04. Diakses tanggal 2021-05-24.
- ^ a b Slonczewski, Joan L.; Foster, John W. (2013). Microbiology: An Evolving Science. W.W. Norton & Company. hlm. 491. ISBN 0-393-12368-5. OCLC 956340090.
- ^ Hellingwerf, K. J.; Crielaard, W.; Hoff, W. D.; Matthijs, H. C. P.; Mur, L. R.; van Rotterdam, B. J. (Desember 1994). "Photobiology of Bacteria". Antonie van Leeuwenhoek. 65 (4): 331–347. doi:10.1007/BF00872217. ISSN 0003-6072.
- ^ Dalton, Howard (29 Juni 2005). "The Leeuwenhoek Lecture 2000 The natural and unnatural history of methane-oxidizing bacteria". Philosophical Transactions of the Royal Society B: Biological Sciences. 360 (1458): 1207–1222. doi:10.1098/rstb.2005.1657. ISSN 0962-8436. PMC 1569495
- ^ Zehr, Jonathan P.; Jenkins, Bethany D.; Short, Steven M.; Steward, Grieg F. (Juli 2003). "Nitrogenase gene diversity and microbial community structure: a cross-system comparison". Environmental Microbiology. 5 (7): 539–554. doi:10.1046/j.1462-2920.2003.00451.x. ISSN 1462-2912.
- ^ Zumft, W G (1997). "Cell biology and molecular basis of denitrification". Microbiology and molecular biology reviews. 61 (4): 533–616. doi:10.1128/61.4.533-616.1997. ISSN 1092-2172. Diarsipkan dari versi asli tanggal 2021-05-25. Diakses tanggal 2021-05-24.
- ^ Drake, Harold L.; Daniel, Steven L.; Küsel, Kirsten; Matthies, Carola; Kuhner, Carla; Braus-Stromeyer, Susanna (1997). "Acetogenic bacteria: what are the in situ consequences of their diverse metabolic versatilities?". BioFactors. 6 (1): 13–24. doi:10.1002/biof.5520060103.
- ^ Morel, François M. M.; Kraepiel, Anne M. L.; Amyot, Marc (November 1998). "The Chemical Cycle and Bioaccumulation of Mercury". Annual Review of Ecology and Systematics. 29 (1): 543–566. doi:10.1146/annurev.ecolsys.29.1.543. ISSN 0066-4162. Diarsipkan dari versi asli tanggal 2021-05-25. Diakses tanggal 2021-05-24.
- ^ Ślesak, Ireneusz; Kula, Monika; Ślesak, Halina; Miszalski, Zbigniew; Strzałka, Kazimierz (Agustus 2019). "How to define obligatory anaerobiosis? An evolutionary view on the antioxidant response system and the early stages of the evolution of life on Earth". Free Radical Biology and Medicine. 140: 61–73. doi:10.1016/j.freeradbiomed.2019.03.004. Diarsipkan dari versi asli tanggal 2021-03-08. Diakses tanggal 2021-05-24.
- ^ Madigan dkk. 2015, hlm. 6–7.
- ^ a b c Whitman, W.B.; Coleman, D.C.; Wiebe, W.J. (9 Juni 1998). "Prokaryotes: The unseen majority". Proceedings of the National Academy of Sciences. 95 (12): 6578–6583. doi:10.1073/pnas.95.12.6578. ISSN 0027-8424. PMC 33863
- ^ Bar-On, Yinon M.; Phillips, Rob; Milo, Ron (19 Mei 2018). "The biomass distribution on Earth". Proceedings of the National Academy of Sciences. 115 (25): 6506–6511. doi:10.1073/pnas.1711842115. ISSN 0027-8424. PMC 6016768
- ^ Jørgensen, Bo Barker; Findlay, Alyssa J.; Pellerin, André (24 April 2019). "The Biogeochemical Sulfur Cycle of Marine Sediments". Frontiers in Microbiology. 10: 849. doi:10.3389/fmicb.2019.00849. ISSN 1664-302X. PMC 6492693
- ^ Fry, Brian; Gest, Howard; Hayes, J.M. (November 1983). "Sulphur isotopic compositions of deep-sea hydrothermal vent animals". Nature. 306 (5938): 51–52. doi:10.1038/306051a0. ISSN 0028-0836. Diarsipkan dari versi asli tanggal 2022-03-07. Diakses tanggal 2021-05-30.
- ^ Wirsen, Carl O.; Jannasch, Holger W.; Molyneaux, Stephen J. (1993). "Chemosynthetic microbial activity at Mid-Atlantic Ridge hydrothermal vent sites". Journal of Geophysical Research. 98 (B6): 9693. doi:10.1029/92JB01556. ISSN 0148-0227.
- ^ a b Segre, Julie A. (18 Desember 2018). "Microbiome". National Human Genome Research Institute. Diarsipkan dari versi asli tanggal 2021-06-02. Diakses tanggal 31 Mei 2021.
- ^ Savage, D.C. (Oktober 1977). "Microbial Ecology of the Gastrointestinal Tract". Annual Review of Microbiology. 31 (1): 107–133. doi:10.1146/annurev.mi.31.100177.000543. ISSN 0066-4227. Diarsipkan dari versi asli tanggal 2021-06-03. Diakses tanggal 2021-05-30.
- ^ Rosner, Judah L. (1 Februari 2014). "Ten Times More Microbial Cells than Body Cells in Humans?". Microbe Magazine. 9 (2): 47–47. doi:10.1128/microbe.9.47.2. ISSN 1558-7452. Diarsipkan dari versi asli tanggal 2020-09-17. Diakses tanggal 2021-05-30.
- ^ Sender, Ron; Fuchs, Shai; Milo, Ron (Januari 2016). "Are We Really Vastly Outnumbered? Revisiting the Ratio of Bacterial to Host Cells in Humans". Cell. 164 (3): 337–340. doi:10.1016/j.cell.2016.01.013. Diarsipkan dari versi asli tanggal 2021-04-09. Diakses tanggal 2021-05-30.
- ^ Madigan dkk. 2015, hlm. 10.
- ^ Rinninella, Emanuele; Raoul, Pauline; Cintoni, Marco; Franceschi, Francesco; Miggiano, Giacinto; Gasbarrini, Antonio; Mele, Maria (10 Januari 2019). "What is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases". Microorganisms. 7 (1): 14. doi:10.3390/microorganisms7010014. ISSN 2076-2607. PMC 6351938
- ^ Heilig, Hans G.H.J.; Zoetendal, Erwin G.; Vaughan, Elaine E.; Marteau, Philippe; Akkermans, Antoon D.L.; de Vos, Willem M. (Januari 2002). "Molecular Diversity of Lactobacillus spp. and Other Lactic Acid Bacteria in the Human Intestine as Determined by Specific Amplification of 16S Ribosomal DNA". Applied and Environmental Microbiology. 68 (1): 114–123. doi:10.1128/AEM.68.1.114-123.2002. ISSN 0099-2240. PMC 126540
- ^ Guandalini, Stefano (November 2011). "Probiotics for Prevention and Treatment of Diarrhea". Journal of Clinical Gastroenterology. 45: S149–S153. doi:10.1097/MCG.0b013e3182257e98. ISSN 0192-0790.
- ^ Azad, Md. Abul Kalam; Sarker, Manobendro; Li, Tiejun; Yin, Jie (2018). "Probiotic Species in the Modulation of Gut Microbiota: An Overview". BioMed Research International. 2018: 1–8. doi:10.1155/2018/9478630. ISSN 2314-6133. PMC 5964481
- ^ Porter, S.R.; Scully, C (23 September 2006). "Oral malodour (halitosis)". BMJ. 333 (7569): 632–635. doi:10.1136/bmj.38954.631968.AE. ISSN 0959-8138.
- ^ a b Madigan dkk. 2015, hlm. 158.
- ^ Rothschild, Lynn J.; Mancinelli, Rocco L. (Februari 2001). "Life in extreme environments". Nature. 409 (6823): 1092–1101. doi:10.1038/35059215. ISSN 0028-0836. Diarsipkan dari versi asli tanggal 2021-05-25. Diakses tanggal 2021-06-25.
- ^ Madigan dkk. 2015, hlm. 160.
- ^ Brock, Thomas D.; Freeze, Hudson (April 1969). "Thermus aquaticus gen. n. and sp. n., a Nonsporulating Extreme Thermophile". Journal of Bacteriology. 98 (1): 289–297. doi:10.1128/jb.98.1.289-297.1969. ISSN 0021-9193. Diarsipkan dari versi asli tanggal 2021-06-24. Diakses tanggal 2021-06-24.
- ^ Clarke, Andrew; Morris, G. John; Fonseca, Fernanda; Murray, Benjamin J.; Acton, Elizabeth; Price, Hannah C. (19 Juni 2013). Neufeld, Josh, ed. "A Low Temperature Limit for Life on Earth". PLoS ONE. 8 (6): e66207. doi:10.1371/journal.pone.0066207. ISSN 1932-6203. PMC 3686811
- ^ Madigan dkk. 2015, hlm. 165.
- ^ Madigan dkk. 2015, hlm. 166.
- ^ a b c d e f g h i j k l m n o Madigan MT (2009). Brock Biology of Microorganisms Twelfth Edition. Pearson Benjammin Cummings.
- ^ Nikiyan H, Vasilchencko A, Deryabin D. 2010. Humidity-Dependent Bacterial Cells Functional Morphometry Investigations Using Atomic Force Microscope. Int J Microbiol. Vol 2010. doi:10.1155/2010/704170.
- ^ Maier RM, Pepper IL, Gerba CP (2009). Environmental Microbiology, 2nd Edition. Elsevier. hlm. 91. ISBN 978-0-12-370519-8.
- ^ a b c d Caldwell A. 2011. The Effects of Ultraviolet Light on Bacterial Growth. http://www.ehow.com/facts_5871403_effects-ultraviolet-light-bacterial-growth.html Diarsipkan 2012-02-15 di Wayback Machine.. Diakses pada 24 Juni 2011.
- ^ a b c d e f g h i j k l m n Todar, Kenneth Gregory. "Todar's Online Textbook of Bacteriology". Diarsipkan dari versi asli tanggal 2019-12-09. Diakses tanggal 2021-05-29.
- ^ a b Shrieve DC, Loeffler JS. 2010. Human Radiation Injury. Halaman 105. Philadelphia: Lippincott Williams & Wilkins. ISBN 978-1-60547-011-5
- ^ a b Mattimore V, Battista JR. 1995. Radioresistance of Deinococcus radiodurans: Functions Necessary To Survive Ionizing Radiation Are Also Necessary To Survive Prolonged Desiccation. J Bacteriol 178(3): 633-637.
- ^ Madigan MT (2009). Brock Biology of Microorganisms Twelfth Edition. Pearson Benjammin Cummings. hlm. 480–481.
- ^ a b c Battista JR, Cox MM. 2005. Deinococcus radiodurans — the consummate survivor. Nat Rev Microbiol 3:882-892. doi:10.1038/nrmicro1264
- ^ Antón, Josefa; Oren, Aharon; Benlloch, Susana; Rodríguez-Valera, Francisco; Amann, Rudolf; Rosselló-Mora, Ramón (1 Maret 2002). "Salinibacter ruber gen. nov., sp. nov., a novel, extremely halophilic member of the Bacteria from saltern crystallizer ponds". International Journal of Systematic and Evolutionary Microbiology. 52 (2): 485–491. doi:10.1099/00207713-52-2-485. ISSN 1466-5026. Diarsipkan dari versi asli tanggal 2021-06-24. Diakses tanggal 2021-06-24.
- ^ "Bakteri Hidup Tinggi di Awan Badai". Diarsipkan dari versi asli tanggal 2016-03-06. Diakses tanggal 29 Januari 2013. Teks "Jurnal KeSimpulan" akan diabaikan (bantuan)
- ^ Koch, Arthur L. (Januari 2002). "Control of the Bacterial Cell Cycle by Cytoplasmic Growth". Critical Reviews in Microbiology. 28 (1): 61–77. doi:10.1080/1040-840291046696. ISSN 1040-841X.
- ^ Eagon, R. G. (1962). "Pseudomonas natriegens, a marine bacterium with a generation time of less than 10 minutes". Journal of Bacteriology. 83 (4): 736–737. doi:10.1128/JB.83.4.736-737.1962. ISSN 0021-9193. Diarsipkan dari versi asli tanggal 2021-03-08. Diakses tanggal 2021-05-24.
- ^ Stewart, Eric J; Madden, Richard; Paul, Gregory; Taddei, François (1 Februari 2005). Kirkwood, Thomas, ed. "Aging and Death in an Organism That Reproduces by Morphologically Symmetric Division". PLoS Biology. 3 (2): e45. doi:10.1371/journal.pbio.0030045. ISSN 1545-7885. PMC 546039
- ^ a b c Thomson, Richard B.; Bertram, Heidi (Desember 2001). "Laboratory diagnosis of central nervous system infections". Infectious Disease Clinics of North America. 15 (4): 1047–1071. doi:10.1016/S0891-5520(05)70186-0. Diarsipkan dari versi asli tanggal 2021-03-08. Diakses tanggal 2021-05-24.
- ^ Paerl, Hans W.; Fulton, Rolland S.; Moisander, Pia H.; Dyble, Julianne (2001). "Harmful Freshwater Algal Blooms, With an Emphasis on Cyanobacteria". The Scientific World JOURNAL. 1: 76–113. doi:10.1100/tsw.2001.16. ISSN 1537-744X. PMC 6083932
- ^ Challis, G. L.; Hopwood, D. A. (25 November 2003). "Synergy and contingency as driving forces for the evolution of multiple secondary metabolite production by Streptomyces species". Proceedings of the National Academy of Sciences. 100 (Supplement 2): 14555–14561. doi:10.1073/pnas.1934677100. ISSN 0027-8424. PMC 304118
- ^ Kooijman, S. A. L. M.; Auger, P.; Poggiale, J. C.; Kooi, B. W. (Agustus 2003). "Quantitative steps in symbiogenesis and the evolution of homeostasis". Biological Reviews. 78 (3): 435–463. doi:10.1017/S1464793102006127.
- ^ Bertrand, Robert L. (14 Januari 2019). Margolin, William, ed. "Lag Phase Is a Dynamic, Organized, Adaptive, and Evolvable Period That Prepares Bacteria for Cell Division". Journal of Bacteriology. 201 (7): e00697–18, /jb/201/7/JB.00697–18.atom. doi:10.1128/JB.00697-18. ISSN 0021-9193. PMC 6416914
- ^ Prats, Clara; López, Daniel; Giró, Antoni; Ferrer, Jordi; Valls, Joaquim (Agustus 2006). "Individual-based modelling of bacterial cultures to study the microscopic causes of the lag phase". Journal of Theoretical Biology. 241 (4): 939–953. doi:10.1016/j.jtbi.2006.01.029. Diarsipkan dari versi asli tanggal 2021-03-08. Diakses tanggal 2021-05-24.
- ^ Hecker, Michael; Völker, Uwe (2001). General stress response of Bacillus subtilis and other bacteria. Advances in Microbial Physiology. 44. Elsevier. hlm. 35–91. doi:10.1016/s0065-2911(01)44011-2. ISBN 978-0-12-027744-5. Diarsipkan dari versi asli tanggal 2021-03-08. Diakses tanggal 2021-05-24.
- ^ Nakabachi, A.; Yamashita, A.; Toh, H.; Ishikawa, H.; Dunbar, H. E.; Moran, N. A.; Hattori, M. (13 Oktober 2006). "The 160-Kilobase Genome of the Bacterial Endosymbiont Carsonella". Science. 314 (5797): 267–267. doi:10.1126/science.1134196. ISSN 0036-8075.
- ^ Pradella, Silke; Hans, Astrid; Spröer, Cathrin; Reichenbach, Hans; Gerth, Klaus; Beyer, Stefan (1 Desember 2002). "Characterisation, genome size and genetic manipulation of the myxobacterium Sorangium cellulosum So ce56". Archives of Microbiology. 178 (6): 484–492. doi:10.1007/s00203-002-0479-2. ISSN 0302-8933.
- ^ Hinnebusch, Joe; Tilly, Kit (Desember 1993). "Linear plasmids and chromosomes in bacteria". Molecular Microbiology. 10 (5): 917–922. doi:10.1111/j.1365-2958.1993.tb00963.x. ISSN 0950-382X.
- ^ Lin, Yi-Shing; Kieser, Helen M.; Hopwood, David A.; Chen, Carton W. (Desember 1993). "The chromosomal DNA of Streptomyces lividans 66 is linear". Molecular Microbiology. 10 (5): 923–933. doi:10.1111/j.1365-2958.1993.tb00964.x. ISSN 0950-382X.
- ^ Val, Marie-Eve; Soler-Bistué, Alfonso; Bland, Michael J; Mazel, Didier (Desember 2014). "Management of multipartite genomes: the Vibrio cholerae model". Current Opinion in Microbiology. 22: 120–126. doi:10.1016/j.mib.2014.10.003. Diarsipkan dari versi asli tanggal 2021-03-08. Diakses tanggal 2021-05-25.
- ^ Kado, Clarence I. (3 Oktober 2014). "Historical Events That Spawned the Field of Plasmid Biology". Microbiology Spectrum. 2 (5). doi:10.1128/microbiolspec.PLAS-0019-2013. ISSN 2165-0497. Diarsipkan dari versi asli tanggal 2021-05-25. Diakses tanggal 2021-05-25.
- ^ Belfort, M; Reaban, M E; Coetzee, T; Dalgaard, J Z (1995). "Prokaryotic introns and inteins: a panoply of form and function". Journal of bacteriology. 177 (14): 3897–3903. doi:10.1128/JB.177.14.3897-3903.1995. ISSN 0021-9193. PMC 177115
- ^ Denamur, Erick; Matic, Ivan (Mei 2006). "Evolution of mutation rates in bacteria". Molecular Microbiology. 60 (4): 820–827. doi:10.1111/j.1365-2958.2006.05150.x. ISSN 0950-382X.
- ^ Wright, Barbara E. (29 Maret 2004). "Stress-directed adaptive mutations and evolution: Stress-directed mutations". Molecular Microbiology. 52 (3): 643–650. doi:10.1111/j.1365-2958.2004.04012.x.
- ^ Chen, Inês; Dubnau, David (Maret 2004). "DNA uptake during bacterial transformation". Nature Reviews Microbiology. 2 (3): 241–249. doi:10.1038/nrmicro844. ISSN 1740-1526. Diarsipkan dari versi asli tanggal 2021-05-26. Diakses tanggal 2021-05-25.
- ^ Johnsborg, Ola; Eldholm, Vegard; Håvarstein, Leiv Sigve (Desember 2007). "Natural genetic transformation: prevalence, mechanisms and function". Research in Microbiology. 158 (10): 767–778. doi:10.1016/j.resmic.2007.09.004. Diarsipkan dari versi asli tanggal 2021-05-25. Diakses tanggal 2021-05-25.
- ^ Brüssow, Harald; Canchaya, Carlos; Hardt, Wolf-Dietrich (1 September 2004). "Phages and the Evolution of Bacterial Pathogens: from Genomic Rearrangements to Lysogenic Conversion". Microbiology and Molecular Biology Reviews. 68 (3): 560–602. doi:10.1128/MMBR.68.3.560-602.2004. ISSN 1092-2172. PMC 515249
- ^ Bickle, T A; Krüger, D H (1993). "Biology of DNA restriction". Microbiological Reviews. 57 (2): 434–450. doi:10.1128/MR.57.2.434-450.1993. ISSN 0146-0749. Diarsipkan dari versi asli tanggal 2021-05-25. Diakses tanggal 2021-05-25.
- ^ Barrangou, R.; Fremaux, C.; Deveau, H.; Richards, M.; Boyaval, P.; Moineau, S.; Romero, D. A.; Horvath, P. (23 Maret 2007). "CRISPR Provides Acquired Resistance Against Viruses in Prokaryotes". Science. 315 (5819): 1709–1712. doi:10.1126/science.1138140. ISSN 0036-8075. Diarsipkan dari versi asli tanggal 2020-12-12. Diakses tanggal 2021-05-25.
- ^ Brouns, S. J. J.; Jore, M. M.; Lundgren, M.; Westra, E. R.; Slijkhuis, R. J. H.; Snijders, A. P. L.; Dickman, M. J.; Makarova, K. S.; Koonin, E. V. (15 Agustus 2008). "Small CRISPR RNAs Guide Antiviral Defense in Prokaryotes". Science. 321 (5891): 960–964. doi:10.1126/science.1159689. ISSN 0036-8075. PMC 5898235
- ^ Michod, Richard E.; Bernstein, Harris; Nedelcu, Aurora M. (Mei 2008). "Adaptive value of sex in microbial pathogens". Infection, Genetics and Evolution. 8 (3): 267–285. doi:10.1016/j.meegid.2008.01.002. Diarsipkan dari versi asli tanggal 2021-04-07. Diakses tanggal 2021-05-25.
- ^ Hastings, P; Rosenberg, S; Slack, A (September 2004). "Antibiotic-induced lateral transfer of antibiotic resistance". Trends in Microbiology. 12 (9): 401–404. doi:10.1016/j.tim.2004.07.003. Diarsipkan dari versi asli tanggal 2021-03-08. Diakses tanggal 2021-05-25.
- ^ a b Bardy, Sonia L.; Ng, Sandy Y. M.; Jarrell, Ken F. (1 Februari 2003). "Prokaryotic motility structures". Microbiology. 149 (2): 295–304. doi:10.1099/mic.0.25948-0. ISSN 1350-0872. Diarsipkan dari versi asli tanggal 2022-10-15. Diakses tanggal 2021-05-25.
- ^ Macnab, Robert M. (1 Desember 1999). "The Bacterial Flagellum: Reversible Rotary Propellor and Type III Export Apparatus". Journal of Bacteriology. 181 (23): 7149–7153. doi:10.1128/JB.181.23.7149-7153.1999. ISSN 1098-5530. Diarsipkan dari versi asli tanggal 2021-03-08. Diakses tanggal 2021-05-25.
- ^ Lux, Renate; Shi, Wenyuan (Juli 2004). "C hemotaxis-guided M ovements in B acteria". Critical Reviews in Oral Biology & Medicine. 15 (4): 207–220. doi:10.1177/154411130401500404. ISSN 1045-4411. Diarsipkan dari versi asli tanggal 2021-03-09. Diakses tanggal 2021-05-25.
- ^ Schweinitzer, Tobias; Josenhans, Christine (Juli 2010). "Bacterial energy taxis: a global strategy?". Archives of Microbiology. 192 (7): 507–520. doi:10.1007/s00203-010-0575-7. ISSN 0302-8933. PMC 2886117
- ^ Frankel, R.B.; Bazylinski, D.A.; Johnson, M.S.; Taylor, B.L. (Agustus 1997). "Magneto-aerotaxis in marine coccoid bacteria". Biophysical Journal. 73 (2): 994–1000. doi:10.1016/S0006-3495(97)78132-3. PMC 1180996
- ^ Nan, Beiyan; Zusman, David R. (15 Desember 2011). "Uncovering the Mystery of Gliding Motility in the Myxobacteria". Annual Review of Genetics. 45 (1): 21–39. doi:10.1146/annurev-genet-110410-132547. ISSN 0066-4197. PMC 3397683
- ^ Goldberg, Marcia B. (1 Desember 2001). "Actin-Based Motility of Intracellular Microbial Pathogens". Microbiology and Molecular Biology Reviews. 65 (4): 595–626. doi:10.1128/MMBR.65.4.595-626.2001. ISSN 1098-5557. PMC 99042
- ^ Shapiro, James A. (Oktober 1998). "Thinking about bacterial populations as multicellular organisms". Annual Review of Microbiology. 52 (1): 81–104. doi:10.1146/annurev.micro.52.1.81. ISSN 0066-4227. Diarsipkan dari versi asli tanggal 2021-05-25. Diakses tanggal 2021-05-27.
- ^ Costerton, J W; Lewandowski, Z; Caldwell, D E; Korber, D R; Lappin-Scott, H M (Oktober 1995). "Microbial Biofilms". Annual Review of Microbiology. 49 (1): 711–745. doi:10.1146/annurev.mi.49.100195.003431. ISSN 0066-4227. Diarsipkan dari versi asli tanggal 2019-04-24. Diakses tanggal 2021-05-27.
- ^ Miller, Daniel P.; Lamont, Richard J. (2019). Belibasakis, Georgios N.; Hajishengallis, George; Bostanci, Nagihan; Curtis, Michael A., ed. Oral Mucosal Immunity and Microbiome. 1197. Cham: Springer International Publishing. hlm. 27–43. doi:10.1007/978-3-030-28524-1_3. ISBN 978-3-030-28523-4.
- ^ Abisado, Rhea G.; Benomar, Saida; Klaus, Jennifer R.; Dandekar, Ajai A.; Chandler, Josephine R. (22 Mei 2018). Garsin, Danielle A., ed. "Bacterial Quorum Sensing and Microbial Community Interactions". mBio. 9 (3): e02331–17, /mbio/9/3/mBio.02331–17.atom. doi:10.1128/mBio.02331-17. ISSN 2150-7511. PMC 5964356
- ^ Miller, Melissa B.; Bassler, Bonnie L. (Oktober 2001). "Quorum Sensing in Bacteria". Annual Review of Microbiology. 55 (1): 165–199. doi:10.1146/annurev.micro.55.1.165. ISSN 0066-4227. Diarsipkan dari versi asli tanggal 2019-04-05. Diakses tanggal 2021-05-27.
- ^ Zhu, Qiyun; Mai, Uyen; Pfeiffer, Wayne; Janssen, Stefan; Asnicar, Francesco; Sanders, Jon G.; Belda-Ferre, Pedro; Al-Ghalith, Gabriel A.; Kopylova, Evguenia (Desember 2019). "Phylogenomics of 10,575 genomes reveals evolutionary proximity between domains Bacteria and Archaea". Nature Communications. 10 (1): 5477. doi:10.1038/s41467-019-13443-4. ISSN 2041-1723. PMC 6889312
- ^ Boucher, Yan; Douady, Christophe J.; Papke, R. Thane; Walsh, David A.; Boudreau, Mary Ellen R.; Nesbø, Camilla L.; Case, Rebecca J.; Doolittle, W. Ford (Desember 2003). "Lateral Gene Transfer and the Origins of Prokaryotic Groups". Annual Review of Genetics. 37 (1): 283–328. doi:10.1146/annurev.genet.37.050503.084247. ISSN 0066-4197. Diarsipkan dari versi asli tanggal 2019-02-16. Diakses tanggal 2021-05-27.
- ^ Olsen, G.J.; Woese, C.R.; Overbeek, R.A. (1 Maret 1996). "The winds of (evolutionary) change: Breathing new life into microbiology". Journal of Bacteriology. 176 (1): 1–6. doi:10.2172/205047. PMC 205007
- ^ "International Journal of Systematic and Evolutionary Microbiology". Microbiology Research. Diarsipkan dari versi asli tanggal 2021-04-16. Diakses tanggal 27 Mei 2021.
- ^ "Bergey's Manual Trust Publications". Bergey's Manual of Systematics of Archaea and Bacteria. Diarsipkan dari versi asli tanggal 2021-05-27. Diakses tanggal 27 Mei 2021.
- ^ Hedlund, Brian P.; Dodsworth, Jeremy A.; Staley, James T. (Juni 2015). "The changing landscape of microbial biodiversity exploration and its implications for systematics". Systematic and Applied Microbiology. 38 (4): 231–236. doi:10.1016/j.syapm.2015.03.003. Diarsipkan dari versi asli tanggal 2021-03-07. Diakses tanggal 2021-05-27.
- ^ Gupta, Radhey S. (Januari 2000). "The Natural Evolutionary Relationships among Prokaryotes". Critical Reviews in Microbiology. 26 (2): 111–131. doi:10.1080/10408410091154219. ISSN 1040-841X.
- ^ Rappé, Michael S.; Giovannoni, Stephen J. (Oktober 2003). "The Uncultured Microbial Majority". Annual Review of Microbiology. 57 (1): 369–394. doi:10.1146/annurev.micro.57.030502.090759. ISSN 0066-4227. Diarsipkan dari versi asli tanggal 2020-03-02. Diakses tanggal 2021-05-27.
- ^ Doolittle, Russell F (Juni 2005). "Evolutionary aspects of whole-genome biology". Current Opinion in Structural Biology. 15 (3): 248–253. doi:10.1016/j.sbi.2005.04.001. Diarsipkan dari versi asli tanggal 2021-06-01. Diakses tanggal 2021-05-27.
- ^ Cavalier-Smith, T (1 Januari 2002). "The neomuran origin of archaebacteria, the negibacterial root of the universal tree and bacterial megaclassification". International Journal of Systematic and Evolutionary Microbiology. 52 (1): 7–76. doi:10.1099/00207713-52-1-7. ISSN 1466-5026. Diarsipkan dari versi asli tanggal 2021-06-13. Diakses tanggal 2021-05-27.
- ^ Woods, G L; Walker, D H (Juli 1996). "Detection of infection or infectious agents by use of cytologic and histologic stains". Clinical Microbiology Reviews. 9 (3): 382–404. doi:10.1128/CMR.9.3.382. ISSN 0893-8512.
- ^ Tang, Silin; Orsi, Renato H.; Luo, Hao; Ge, Chongtao; Zhang, Guangtao; Baker, Robert C.; Stevenson, Abigail; Wiedmann, Martin (12 Juli 2019). "Assessment and Comparison of Molecular Subtyping and Characterization Methods for Salmonella". Frontiers in Microbiology. 10: 1591. doi:10.3389/fmicb.2019.01591. ISSN 1664-302X. PMC 6639432
- ^ Weinstein, Melvin P. (Maret 1994). "Clinical Importance of Blood Cultures". Clinics in Laboratory Medicine. 14 (1): 9–16. doi:10.1016/S0272-2712(18)30390-1. Diarsipkan dari versi asli tanggal 2021-03-07. Diakses tanggal 2021-05-27.
- ^ Riley, Lee W. (12 November 2018). "Laboratory Methods in Molecular Epidemiology: Bacterial Infections *". Microbiology Spectrum. 6 (6). doi:10.1128/microbiolspec.AME-0004-2018. ISSN 2165-0497. Diarsipkan dari versi asli tanggal 2021-04-20. Diakses tanggal 2021-05-27.
- ^ Lee, Ki-Soo; Oh, Chang-Gun; Yim, Jin-Heong; Ihm, Son-Ki (Oktober 2000). "Characteristics of zirconocene catalysts supported on Al-MCM-41 for ethylene polymerization". Journal of Molecular Catalysis A: Chemical. 159 (2): 301–308. doi:10.1016/S1381-1169(00)00220-X. PMC 80298
- ^ Oliver, James D. (Februari 2005). "The viable but nonculturable state in bacteria". Journal of Microbiology (Seoul, Korea). 43 Spec No: 93–100. ISSN 1225-8873. PMID 15765062. Diarsipkan dari versi asli tanggal 2021-02-24. Diakses tanggal 2021-05-27.
- ^ Curtis, T.P.; Sloan, W.T.; Scannell, J.W. (6 Agustus 2002). "Estimating prokaryotic diversity and its limits". Proceedings of the National Academy of Sciences. 99 (16): 10494–10499. doi:10.1073/pnas.142680199. ISSN 0027-8424. PMC 124953
- ^ Schloss, Patrick D.; Handelsman, Jo (Desember 2004). "Status of the Microbial Census". Microbiology and Molecular Biology Reviews. 68 (4): 686–691. doi:10.1128/MMBR.68.4.686-691.2004. ISSN 1092-2172. PMC 539005
- ^ Callewaert, Chris; Lambert, Jo; Van de Wiele, Tom (Mei 2017). "Towards a bacterial treatment for armpit malodour". Experimental Dermatology. 26 (5): 388–391. doi:10.1111/exd.13259.
- ^ Martin, Mark O. (September 2002). "Predatory prokaryotes: an emerging research opportunity". Journal of Molecular Microbiology and Biotechnology. 4 (5): 467–477. ISSN 1464-1801. PMID 12432957. Diarsipkan dari versi asli tanggal 2021-02-25. Diakses tanggal 2021-05-27.
- ^ Velicer, Gregory J.; Stredwick, Kristina L. (2002). "Experimental social evolution with Myxococcus xanthus". Antonie van Leeuwenhoek. 81: 155–164. doi:10.1023/A:1020546130033.
- ^ Gromov, B.V.; Mamkaeva, K. A. (Februari 1972). "Electron microscopic study of parasitism by Bdellovibrio chlorellavorus bacteria on cells of the green alga Chlorella vulgaris". Tsitologiia. 14 (2): 256–260. ISSN 0041-3771. PMID 5011884. Diarsipkan dari versi asli tanggal 2021-05-27. Diakses tanggal 2021-05-27.
- ^ Guerrero, R.; Pedros-Alio, C.; Esteve, I.; Mas, J.; Chase, D.; Margulis, L. (1 April 1986). "Predatory prokaryotes: Predation and primary consumption evolved in bacteria". Proceedings of the National Academy of Sciences. 83 (7): 2138–2142. doi:10.1073/pnas.83.7.2138. ISSN 0027-8424. PMC 323246
- ^ Velicer, Gregory J.; Mendes-Soares, Helena (Januari 2009). "Bacterial predators". Current Biology. 19 (2): R55–R56. doi:10.1016/j.cub.2008.10.043. Diarsipkan dari versi asli tanggal 2021-03-08. Diakses tanggal 2021-05-27.
- ^ Stams, Alfons J.M.; de Bok, Frank A.M.; Plugge, Caroline M.; van Eekert, Miriam H.A.; Dolfing, Jan; Schraa, Gosse (Maret 2006). "Exocellular electron transfer in anaerobic microbial communities". Environmental Microbiology. 8 (3): 371–382. doi:10.1111/j.1462-2920.2006.00989.x. ISSN 1462-2912.
- ^ Zinser, Erik R. (Agustus 2018). "Cross-protection from hydrogen peroxide by helper microbes: the impacts on the cyanobacterium Prochlorococcus and other beneficiaries in marine communities: HOOH and cross-protection". Environmental Microbiology Reports. 10 (4): 399–411. doi:10.1111/1758-2229.12625.
- ^ Barea, José-Miguel; Pozo, María José; Azcón, Rosario; Azcón-Aguilar, Concepción (1 Juli 2005). "Microbial co-operation in the rhizosphere". Journal of Experimental Botany. 56 (417): 1761–1778. doi:10.1093/jxb/eri197. ISSN 1460-2431. Diarsipkan dari versi asli tanggal 2021-12-31. Diakses tanggal 2021-05-27.
- ^ O'Hara, Ann M; Shanahan, Fergus (Juli 2006). "The gut flora as a forgotten organ". EMBO reports. 7 (7): 688–693. doi:10.1038/sj.embor.7400731. ISSN 1469-221X. PMC 1500832
- ^ Zoetendal, Erwin G.; Vaughan, Elaine E.; de Vos, Willem M. (Maret 2006). "A microbial world within us". Molecular Microbiology. 59 (6): 1639–1650. doi:10.1111/j.1365-2958.2006.05056.x. ISSN 0950-382X.
- ^ Gorbach, Sherwood L. (Januari 1990). "Lactic Acid Bacteria and Human Health". Annals of Medicine. 22 (1): 37–41. doi:10.3109/07853899009147239. ISSN 0785-3890.
- ^ Salminen, Seppo J.; Gueimonde, Miguel; Isolauri, Erika (1 Mei 2005). "Probiotics That Modify Disease Risk". The Journal of Nutrition. 135 (5): 1294–1298. doi:10.1093/jn/135.5.1294. ISSN 0022-3166. Diarsipkan dari versi asli tanggal 2021-05-25. Diakses tanggal 2021-05-27.
- ^ Harvey, Richard A.; Champe, Pamela C.; Fisher, Bruce D. (2007). Lippincott's Illustrated Reviews: Microbiology (edisi ke-2). Philadelphia: Lippincott Williams & Wilkins. hlm. 367–392. ISBN 0-7817-8215-5. OCLC 67817144.
- ^ Schwarz, Stefan; Enne, Virve I.; van Duijkeren, Engeline (Oktober 2016). "40 years of veterinary papers in JAC – what have we learnt?". Journal of Antimicrobial Chemotherapy. 71 (10): 2681–2690. doi:10.1093/jac/dkw363. ISSN 0305-7453. Diarsipkan dari versi asli tanggal 2022-01-15. Diakses tanggal 2021-05-27.
- ^ Fish, Douglas N. (1 Februari 2002). "Optimal antimicrobial therapy for sepsis". American Journal of Health-System Pharmacy. 59 (suppl_1): S13–S19. doi:10.1093/ajhp/59.suppl_1.S13. ISSN 1079-2082. Diarsipkan dari versi asli tanggal 2021-03-08. Diakses tanggal 2021-05-27.
- ^ Belland, Robert J.; Ouellette, Scot P.; Gieffers, Jens; Byrne, Gerald I. (Februari 2004). "Chlamydia pneumoniae and atherosclerosis". Cellular Microbiology. 6 (2): 117–127. doi:10.1046/j.1462-5822.2003.00352.x.
- ^ Heise, E.R. (Februari 1982). "Diseases associated with immunosuppression". Environmental Health Perspectives. 43: 9–19. doi:10.1289/ehp.82439. ISSN 0091-6765. Diarsipkan dari versi asli tanggal 2021-03-18. Diakses tanggal 2021-05-27.
- ^ Saiman, Lisa (Januari 2004). "Microbiology of early CF lung disease". Paediatric Respiratory Reviews. 5: S367–S369. doi:10.1016/S1526-0542(04)90065-6. Diarsipkan dari versi asli tanggal 2021-03-09. Diakses tanggal 2021-05-27.
- ^ Yonath, Ada; Bashan, Anat (Oktober 2004). "Ribosomal Crystallography: Initiation, Peptide Bond Formation, and Amino Acid Polymerization are Hampered by Antibiotics". Annual Review of Microbiology. 58 (1): 233–251. doi:10.1146/annurev.micro.58.030603.123822. ISSN 0066-4227. Diarsipkan dari versi asli tanggal 2020-03-12. Diakses tanggal 2021-05-27.
- ^ Khachatourians, G.G. (3 November 1998). "Agricultural use of antibiotics and the evolution and transfer of antibiotic-resistant bacteria". CMAJ: Canadian Medical Association journal. 159 (9): 1129–1136. ISSN 0820-3946. PMC 1229782
- ^ Kuo, Jeff (1 Oktober 2017). "Disinfection Processes". Water Environment Research. 89 (10): 1206–1244. doi:10.2175/106143017X15023776270278. ISSN 1061-4303.
- ^ a b c d e Madigan MT (2009). Brock Biology of Microorganisms Twelfth Edition. Pearson Benjammin Cummings. hlm. 403–404.
- ^ Carlson CA, Ingraham JL. 1983. Comparison of denitrification by Pseudomonas stutzeri, Pseudomonas aeruginosa, and Paracoccus denitrificans. Appl Environ Microbiol 45(4):1247–1253.
- ^ Nitrogen Fixing Bacteria Diarsipkan 2011-12-07 di Wayback Machine.. 2011. Diakses pada 26 Juli 2011.
- ^ a b Deacon J. The Microbial World: The Nitrogen cycle and Nitrogen fixation Diarsipkan 2011-08-07 di Wayback Machine. Diakases pada 26 Juli 2011.
- ^ a b c Marler B. 2010. Clostridium Botulinum (Botulism). http://www.foodborneillness.com/botulism_food_poisoning/ Diarsipkan 2011-06-12 di Wayback Machine.. Diakses pada 24 Juni 2011.
- ^ Welling W, Cohen JA, Berends W. 1960. Disturbance of oxidative phosphorylation by an antibioticum produced by pseudomonas cocovenenans. Biochem Pharmacol 3(2):122-135. doi:10.1016/0006-2952(60)90028-9.
- ^ "Fermented and vegetables. A global perspective. Chapter 5". www.fao.org. Diarsipkan dari versi asli tanggal 2018-10-09. Diakses tanggal 19 Apr 2020.
- ^ "Seleksi dan Karakterisasi Bakteri Endofit untuk Menekan Kejadian Penyakit Layu Bakteri (Ralstonia solanacearum) pada Tanaman Tomat". Diarsipkan dari versi asli tanggal 2013-06-12. Diakses tanggal 2013-06-07.
- ^ "Potensi Bacillus spp. dan Pseudomonas fluorescens sebagai Agens Pengendali Penyakit Busuk Lunak Bakteri (Erwinia carotovora) pada Anggrek Phalaenopsis". Diarsipkan dari versi asli tanggal 2013-07-07. Diakses tanggal 2013-06-07.
- ^ "Seleksi dan Identifikasi Aktinomiset sebagai Agens Hayati untuk Pengendalian Penyakit Kresek yang Diakibatkan oleh Xanthomonas oryzae pv. oryzae pada Padi". Diarsipkan dari versi asli tanggal 2016-03-04. Diakses tanggal 2013-06-07.
- ^ "Pengaruh Mulsa dan PGPR Terhadap Insidensi Penyakit Busuk Pangkal Batang (Sclerotium rolfsii Sacc.) pada Tanaman Kedelai (Glycine max (L) Merill)". Diarsipkan dari versi asli tanggal 2016-06-16. Diakses tanggal 2013-06-07.
- ^ "Eksplorasi Agens Antagonis yang Berpotensi Menekan Penyakit Fusarium pada Pisang". Diarsipkan dari versi asli tanggal 2016-06-16. Diakses tanggal 2013-06-07.
- ^ a b Parry CM, Hien TT, Dougan G, White NJ, Farrar JJ. 2002. Typhoid fever. N Engl J Med 347:1770–1782.
- ^ Medie FM, Salahi IB, Drancourt M, Henrissat B. 2010. Paradoxical conservation of a set of three cellulose-targeting genes in Mycobacterium tuberculosis complex organisms. Microbiol 156:1468-1475. doi: 10.1099/mic.0.037812-0.
- ^ Rodriguez MC, Froger A, Rolland JP, Thomas D, Aguerol J, Delamarche C, Garcia-Lobo JM. A functional water channel protein in the pathogenic bacterium Brucella abortus. Microbiol 146(12):3251-3257. doi: 3251-3257.
- ^ Feng JX, Song ZZ, Duan CJ, Zhao S, Wu YQ, Wang C, Dow JM, Tang JL. 2009. The xrvA gene of Xanthomonas oryzae pv. oryzae, encoding an H-NS-like protein, regulates virulence in rice. Microbiol 155(9):3033-44.
- ^ a b Decomposition by bacteria Diarsipkan 2013-06-01 di Wayback Machine.. Diakses pada 24 Juni 2011.
- ^ a b c d e Decomposition of Organic Matter[pranala nonaktif permanen]. Diakses pada 24 Juni 2011.
Bacaan lebih lanjut
[sunting | sunting sumber]- Alcamo IE (2001). Fundamentals of microbiology. Boston: Jones and Bartlett. ISBN 0-7637-1067-9.
- Atlas RM (1995). Principles of microbiology. St. Louis: Mosby. ISBN 0-8016-7790-4.
- Holt JC, Bergey DH (1994). Bergey's manual of determinative bacteriology (edisi ke-9th ed.). Baltimore: Williams & Wilkins. ISBN 0-683-00603-7.
- Madigan, Michael T.; Bender, Kelly S.; Buckley, Daniel Hezekiah; Stahl, David Allan (2015). Brock Biology of Microorganisms (edisi ke-14). Boston: Pearson. ISBN 978-0-321-89739-8. OCLC 857863493.
- Tortora, Gerard J.; Funke, Berdell L.; Case, Christine L. (2010). Microbiology: An Introduction (edisi ke-10). San Francisco, CA: Pearson Benjamin Cummings.
Pranala luar
[sunting | sunting sumber]| Cari tahu mengenai Bakteri pada proyek-proyek Wikimedia lainnya: | |
| Definisi dan terjemahan dari Wiktionary | |
| Gambar dan media dari Commons | |
| Berita dari Wikinews | |
| Kutipan dari Wikiquote | |
| Teks sumber dari Wikisource | |
| Buku dari Wikibuku | |
- MicrobeWiki, an extensive wiki about bacteria and viruses
- Bacteria that affect crops and other plants
- Bacterial Nomenclature Up-To-Date from DSMZ Diarsipkan 2007-09-29 di Wayback Machine.
- Genera of the domain Bacteria Diarsipkan 2008-09-17 di Wayback Machine. – list of Prokaryotic names with Standing in Nomenclature
- The largest bacteria Diarsipkan 2012-12-11 di Wayback Machine.
- Tree of Life: Eubacteria
- Videos Diarsipkan 2015-11-05 di Wayback Machine. of bacteria swimming and tumbling, use of optical tweezers and other videos.
- Planet of the Bacteria Diarsipkan 2019-06-22 di Wayback Machine. by Stephen Jay Gould
- On-line text book on bacteriology
- Animated guide to bacterial cell structure.
- Bacteria Make Major Evolutionary Shift in the Lab
- Online collaboration for bacterial taxonomy.
- PATRIC, a Bioinformatics Resource Center for bacterial pathogens, funded by NIAID
- Bacterial Chemotaxis Interactive Simulator – A web-app that uses several simple algorithms to simulate bacterial chemotaxis.
- Cell-Cell Communication in Bacteria Diarsipkan 2009-01-30 di Wayback Machine. on-line lecture by Bonnie Bassler, and TED: Discovering bacteria's amazing communication system Diarsipkan 2011-12-05 di Wayback Machine.
- Sulfur-cycling fossil bacteria from the 1.8-Ga Duck Creek Formation provide promising evidence of evolution's null hypothesis, Proceedings of the National Academy of Sciences of the United States of America. Summarized in: Scientists discover bacteria that haven't evolved in more than 2 billion years, LiveScience and BusinessInsider
| Umum | |
|---|---|
| Perpustakaan nasional | |
| Lain-lain | |