Sejtmag

A sejtmag (latinul nucleus) kettős membránnal körülvett organellum az eukarióta sejten belül. Elsődleges funkciója a sejt genetikai anyagának, a kromoszómáknak a tárolása és elkülönítése a citoplazmától, valamint a gének kifejeződésének szabályozása. Belső tartalmát a citoplazmától a maghártya – kettős lipidréteg – különíti el. Formáját a sejtmagváz, a nukleoszkeleton adja.
A fehérjék, nukleinsavak és egyéb molekulák a bonyolult szerkezetű magpórusokon áthaladva jutnak át a maghártyán, ez egy gondosan szabályozott folyamat. A sejtmagban nincsenek más, membránnal elkülönített organellumok, de belseje nem egyenletes, abban több, fehérjéből és ribonukleinsavból álló szubnukleáris test is megfigyelhető. Közülük legismertebb a riboszómákat összeállító sejtmagvacska (nucleolus).
A sejtmag volt a sejt először felfedezett szervecskéje (organelluma). Eddig ismert legrégebbi ábrázolását az első mikrobiológus, a mikroszkópot tökéletesítő 17. századi Anton van Leeuwenhoek vetette papírra. Ő a lazac vörösvérsejtjeiben figyelt meg belső „űrt” (az emlősök vörösvértesteitől eltérően a többi gerinces állat vérsejtjeinek van sejtmagjuk). 1804-ben Franz Bauer is leírta a sejtmagot, majd részletesebben 1831-ben a skót botanikus Robert Brown számolt be róla a Londoni Linné Társaság egyik ülésén. Brown orchideák virágainak külső sejtrétegében figyelt meg áttetsző, különálló részt, amelyet areolának vagy nucleusnak nevezett el, de azt nem kötötte valamilyen sejtfunkcióhoz. 1838-ban a német Matthias Schleiden felvetette, hogy a sejtmag az új sejtek létrehozásában játszhat szerepet, és erre alapozva a citoblaszt (sejtépítő) elnevezést javasolta. Azt állította, hogy sikerült megfigyelnie a „citoblasztok” körül csoportosuló kis sejteket. Franz Meyen határozottan ellenezte ezt az elképzelést, mert korábban leírta a sejtosztódást, és nézete szerint sok sejtnek egyáltalán nem volt magja. Az újonnan, semmiből létrejövő sejtek elmélete Robert Remak (1852) és Rudolf Virchow (1855) eredményeinek is ellentmondott, akik szerint sejt csak sejtből jöhet létre (Omnis cellula e cellula). A sejtmag funkciója ekkor még ismeretlen maradt.
1877–78-ban Oscar Hertwig több közleményben publikálta megfigyeléseit a tengerisünök petesejtjének megtermékenyüléséről. Kimutatta, hogy a spermium sejtmagja behatol a petesejtbe, és ott egyesül annak sejtmagjával; emellett bizonyította, hogy az élőlények egyetlen, nukleusszal rendelkező sejtből fejlődnek ki. Ezzel ellentmondott Ernst Haeckel korábbi elméletének, miszerint az egyedfejlődés teljesen megismétli a törzsfejlődés útját, beleértve azt is, hogy a kezdeti sejt egy strukturálatlan masszából jön létre. Az elméletét ért támadások után Hertwig bebizonyította, hogy a sejtmagra a puhatestűek és a kétéltűek megtermékenyülésénél is feltétlenül szükség van. 1884-ben ugyanerre jutott a növényeket tanulmányozó Eduard Strasburger. Fokozatosan felismerték, hogy a sejtmag az öröklődésben játszik fontos szerepet. 1873-ban August Weismann kimondta, hogy az öröklődésben az apai és anyai ivarsejtek egyformán fontosak. Miután megfigyelték a mitózist és a 20. század elején újra felfedezték Mendel törvényeit, egyértelművé vált, hogy a sejtmag a genetikai információk hordozója.
Felfedezése
[szerkesztés]

A sejtmag volt az első sejten belüli szervecske (organellum), amit felfedeztek. Az eddig ismert legrégebbi ábrázolása az első mikrobiológustól, a mikroszkóp tökéletesítőjétől, a 17. századi Anton van Leeuwenhoek-től származik, aki a lazac vörösvérsejtjeiben figyelt meg belső „űrt”[1] (az emlősök vörösvértestjeitől eltérően a többi gerinces állat vérsejtjeinek van sejtmagjuk). 1804-ben Franz Bauer is leírta a sejtmagot,[2] majd részletesebben 1831-ben a skót botanikus, Robert Brown számolt be róla a Londoni Linné Társaság egyik ülésén. Brown orchideák virágjának külső sejtrétegében figyelt meg áttetsző különálló részt, melyet ő areolának vagy nucleusnak nevezett el,[3] de nem kötötte azt valamilyen sejtfunkcióhoz. 1838-ban a német Matthias Schleiden felvetette, hogy a sejtmag az új sejtek létrehozásában játszik szerepet, és a citoblaszt (sejtépítő) elnevezést javasolta, és azt állította, hogy sikerült megfigyelnie a „citoblasztok” körül csoportosuló kis sejteket. Franz Meyen határozottan ellenezte ezt az elképzelést, mert korábban leírta a sejtosztódást, és nézete szerint sok sejtnek egyáltalán nem volt magja. Az újonnan, semmiből létrejövő sejtek elmélete Robert Remak (1852) és Rudolf Virchow (1855) eredményeinek is ellentmondott, akik szerint sejt csak sejtből jöhet létre (Omnis cellula e cellula). A sejtmag funkciója ekkor még ismeretlen maradt.[4]
1877–78-ban Oscar Hertwig több közleményben publikálta megfigyeléseit a tengeri sünök petesejtjének megtermékenyüléséről. Kimutatta, hogy a spermium sejtmagja behatol a petesejtbe és ott egyesül annak sejtmagjával; emellett bebizonyította, hogy az élőlények egyetlen, nukleusszal rendelkező sejtből fejlődnek ki. Ezzel ellentmondott Ernst Haeckel korábbi elméletének, miszerint az egyedfejlődés teljes mértékben megismétli a törzsfejlődési utat, beleértve azt, hogy a kiindulási sejt egy strukturálatlan masszából („Urschleim”) jön létre. Az elméletét ért támadások után Hertwig bebizonyította, hogy a sejtmagra a puhatestűek és a kétéltűek megtermékenyülésénél is feltétlenül szükség van. Eduard Strasburger 1884-ben ugyanerre jutott a növények tanulmányozása során. Fokozatosan felismerték, hogy a sejtmag az öröklődésben játszik fontos szerepet. 1873-ban August Weismann kimondta, hogy az öröklődés során az apai és anyai ivarsejtek egyenlő jelentőséggel bírnak. Miután megfigyelték a mitózist és a 20. század elején újra felfedezték Mendel törvényeit, egyértelművé vált, hogy a sejtmag a genetikai információk hordozója.[4]
Felépítése
[szerkesztés]A sejtmag az állati sejt legnagyobb organelluma.[5] Az emlőssejt nukleuszának átmérője kb. 6 mikrométer, és a sejt térfogatának 10%-át foglalja el.[6] Belsejét viszkózus folyadék, a magplazma (nukleoplazma vagy karioplazma) tölti ki, amelynek összetétele hasonló a citoszoléhoz.[7] Alakja többnyire gömbszerű, de számos szabálytalan sejtmagforma is ismert (korong, pálca, karéjozott stb.).
A maghártya és a magpórusok
[szerkesztés] |
A maghártya két, egymás mellett elhelyezkedő sejtmembránból, a külső és belső membránból áll, amelyeket egy 10–50 nanométeres rés választ el egymástól. A maghártya feladata, hogy elválassza a sejt genetikai anyagát, a kromoszómákat az őt körülvevő citoplazmától, és megakadályozza a makromolekulák szabad átjárását a citoplazma és a nukleoplazma között.[8] A külső membrán közvetlenül kapcsolódik a durva felszínű endoplazmatikus retikulumhoz, és hozzá hasonlóan riboszómákkal van teleszórva.[8] A két membrán közötti rés a perinukleáris űr, és közvetlenül folytonos az endoplazmatikus retikulum belső terével.
A maghártyán keresztül történő transzport a magpórusokon át zajlik. A pórusok bonyolult szerkezetű képződmények, amelyek 50 (élesztőben) vagy néhány száz (gerincesekben) fehérjéből tevődnek össze, és tömegük eléri a 125 millió daltont.[5] A pórus teljes átmérője 100 nm, de a tényleges tér, ahol a molekulák szabadon átdiffundálhatnak, alig 9 nm. Ezáltal a kis, vízoldékony vegyületek szabadon átjárnak, de a pórus meggátolja a nagy makromolekulák – fehérjék, nukleinsavak – irányítatlan közlekedését. A nagy molekulák aktív transzporttal kerülnek be a sejtmagba vagy ki a sejtmagból. Egy átlagos emlőssejtmag felületén mintegy 3–4 ezer pórus található.[9] A pórus egy nyolcszoros szimmetriájú, gyűrűszerű szerkezet, amely mellett a külső és belső membránok egybeolvadnak.[10] A gyűrűhöz belülről (a nukleoplazma felől) egy kosárnak nevezett szerkezet kapcsolódik, míg a másik oldalon szálszerű filamentumok nyúlnak a citoplazmába. Mindkét struktúra a transzportfehérjék kötődésére szolgál.[5]
A legtöbb fehérjét, riboszómaalegységet és egyes DNS-eket a karioferinnek nevezett transzportproteinek viszik át a pórusokon. A sejtmagba befelé történő szállítást szabályozó karioferineket importinnak, a kifelé történőt exportinnak nevezik. A legtöbb karioferin közvetlenül kapcsolódik a szállítmányához, bár bizonyos esetekben adaptor proteineket is használnak.[11] Az olyan lipidoldékony szteroidhormonok, mint a hidrokortizon, vagy az aldoszteron képesek átdiffundálni a sejthártyán, és a citoplazmában ún. nukleáris receptorok kötik meg őket, amelyek ezután aktív transzporttal a sejtmagba vándorolnak, ahol a génkifejeződést szabályozzák: a hormonnal kapcsolódva serkentik, anélkül gátolják az adott gén átírását.[5]
Nukleáris lamina
[szerkesztés]Az állati sejtekben két, intermedier filamentumokból álló állványzat biztosítja a sejtmag alakjának megtartását. A nukleáris lamina a maghártya belső felén alkot jól szervezett hálózatot, míg a hártya külső, citoplazma felőli oldalát egy kevésbé sűrű filamentumegyüttes támasztja meg. Mindkét rendszer segít a maghártya alakjának megőrzésében, és kapcsolódási pontokat nyújt a kromoszómáknak és a pórusoknak.[6]
A nukleáris lamina főleg a lamin fehérjéből áll. A többi proteinhez hasonlóan a lamin is a citoplazmában szintetizálódik, majd transzporttal bekerül a sejtmagba, ahol beépül a nukleáris lamina már meglévő hálózatába.[12][13] A maghártya külső oldalára irányított laminok (pl. az emerin és a nesprin) a citoszkeletonhoz kapcsolódva rögzítik a sejtmagot. Laminokat a nukleoplazmában is találhatunk, ahol az ún. nukleoplazmatikus fátylat hozzák létre,[14] amely fluoreszcenciamikroszkópban jól megfigyelhető. A fátyol funkciója ismeretlen, de a sejtmagvacskában nincs jelen, és a két sejtosztódás közötti interfázisban lehet megfigyelni.[15] A fátyol a kromoszómákhoz kapcsolódik, és ha ezt a kötődést meggátoljuk, akkor a gének átírása megszűnik.[16]
A többi intermedier filamentumhoz hasonlóan a lamin alapegysége, fehérjemonomerje alfa-hélix szerkezetű. Két ilyen alegység egymás köré tekeredve alkot egy dimert, majd két dimer egymás mellett, fej-láb elhelyezkedéssel a protofilamentumnak nevezett tetramert hozza létre. Nyolc egymáshoz kapcsolódott protofilamentum alkotja a csavart, kötélszerű filamentumot, melynek hossza a kapcsolódó, illetve leváló alegységek miatt folyamatosan változhat.[6]
A lamin egyes mutációi elégtelen szálösszeálláshoz vezethetnek, és a ritka, laminopátia nevű öröklődő betegségcsoportot okozzák. A legismertebb ilyen betegség a progeria, amely a betegek korai, akár kisgyerekkori testi öregedésével jár. Nem teljesen ismert, hogy a biokémiai változások hogyan vezetnek az öregedés külső szimptómáihoz.[17]
Kromoszómák
[szerkesztés]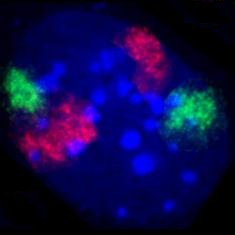
A sejtmag tartalmazza a sejt genetikai információjának döntő többségét (valamennyi a mitokondriumokban és a növényi kloroplasztiszokban is található). Az információ hosszú, lineáris DNS-molekulákban van kódolva, amelyek a kromoszómákat alkotják. Minden egyes kromoszóma egyetlen, hosszú DNS-szálat tartalmaz, illetve osztódáskor, amikor megduplázódik, akkor kettőt, ezek adják a kariotípusból ismert X-formát. Minden emberi sejt kb. két méternyi DNS-t tartalmaz. A kromoszóma anyaga a kromatin, a DNS és a hozzá kapcsolódó szabályozó és struktúrfehérjék (hisztonok) együttese.
A kromatinnak két fajtája ismert. Az eukromatin lazább szerkezetű, és azokat a géneket tartalmazza, amelyekre a sejtnek gyakran van szüksége, sokszor átír.[18] A heterokromatin viszont jóval kompaktabb, és a kevéssé használt DNS-szakaszok tartoznak hozzá. A heterokromatin további két részre osztható, a fakultatív heterokromatinra, amelynek génjeit időnként azért átírják (például a szervezet bizonyos fejlődési stádiumában), illetve az állandó heterokromatinra, amelyhez az olyan strukturális régiók tartoznak, mint a telomerek és centromerek.[19] Az interfázis alatt a fellazult, normál fénymikroszkópban nem látható kromoszómák adott „kromoszómaterületen” helyezkednek el.[20][21] Az aktív, eukromatikus gének ilyenkor többnyire a terület határán találhatók.[22]
Egyes autoimmun betegségekben, mint a szisztémás lupus erythematodesben, a kromatin alegységeihez, a nukleoszómákhoz kötődő ellenanyagok képződnek.[23] Hasonló nukleáris ellenanyagokat sclerosis multiplexben is megfigyeltek, és kapcsolatba hozták az immunrendszer általános zavarával.[24] A progeriához hasonlóan nem világos, hogyan vezet az ellenanyagok termelődése a betegségek konkrét tüneteihez. Bár korábban úgy gondolták, hogy az autoimmun betegségekben az autoantitestek nem képesek behatolni a sejtmagokba, újabban bizonyíték van ennek ellenkezőjére.[25]
Sejtmagvacska
[szerkesztés]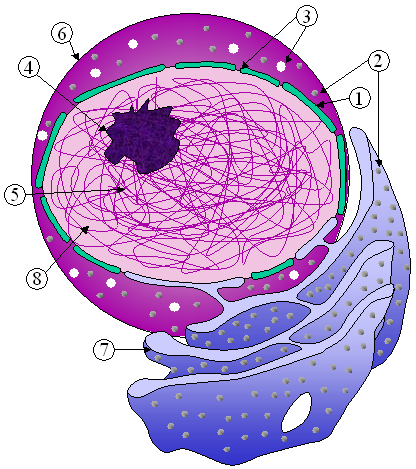
A sejtmagvacska vagy nukleólusz (nucleolus) egy különálló, sötéten festődő struktúra a sejtmagon belül. Nem veszi körül külön membrán, és néha szuborganellumként emlegetik. A sejtmagvacska a kromoszómák riboszomális RNS-t (rRNS) kódoló régiói körül formálódik; ezek a gének nagyon sok példányban vannak jelen egymás mellett. A nukleólusz fő funkciója a riboszomális RNS szintézise és a riboszómák összeállítása. Struktúrájának tömörsége aktivitásától függ, a riboszóma-összeszerelés fokozza a sejtmagvacska komponenseinek ideiglenes összekapcsolódását, amely még hatékonyabb riboszómakészítéshez vezet. Megfigyelték, hogy ha inaktiválják az rRNS-t, a sejtmagvacska struktúrái összekuszálódnak.[26]
A riboszóma-összeszerelés első lépéseként az RNS-polimeráz I a DNS-ről átírja az rRNS génjét, miáltal egy hosszú pre-rRNS keletkezik. Ennek feldarabolódásával jönnek létre az alegységek 5,8S, 18S és 28S rRNS-ei.[27] Az átírás (transzkripció), feldarabolás és a riboszóma-alegységek összeállítása a sejtmagvacskában történik a kis nukleoláris RNS (snoRNS) segítségével. Utóbbiak részben a riboszómák funkcióihoz kapcsolódó gének mRNS-einek kivágott intronjaiból származnak. Az összeállított riboszóma-alegységek a legnagyobb struktúrák, amelyek átférnek a maghártya pórusain.[5]
Elektronmikroszkóp alatt a neukleóluszban három régió különböztethető meg. A legbelső fibrilláris centrumokat (FC) az elektrondenz fibrilláris komponens (dense fibrillar component, DFC) veszi körül, amelyet viszont a granuláris komponens (granular component, GC) határol. Az rRNS transzkripciója az FC régióban vagy az FC-DFC határon zajlik, így ha a sejt igénye megnő a riboszómákra, a sejtmagvacskában több FC-t lehet megfigyelni. A pre-rRNS szétvágása és utólagos módosítása a DFC-ben, a fehérjék hozzákapcsolása az alegységek elkészítéséhez pedig a GC-ben történik.[27]
További struktúrák
[szerkesztés]| Név | Átmérő | |
|---|---|---|
| Cajal-testek | 0,2–2,0 µm | [28] |
| PIKA | 5 µm | [29] |
| PML-testek | 0,2–1,0 µm | [30] |
| Parafoltok | 0,2–1,0 µm | [31] |
| Foltok | 20–25 nm | [29] |
A sejtmagvacskán kívül a mag több kisebb, membránnal nem határolt testet tartalmaz. Ilyenek a Cajal-testek, az iker-spiráltestek, a polimorf interfázisbeli karioszomális halmaz (PIKA), a promielocitás leukémia (PML)-testek, a parafoltok és a splicing-foltok. Sokukról nagyon kevés információ áll rendelkezésre, de egyértelművé teszik, hogy a nukleoplazma egyáltalán nem homogén, hanem funkcionális kisebb terekre oszlik.[30]
Egyes szubnukleáris struktúrák bizonyos betegségekben tűnnek fel. Miopathia nemalinica esetén kis pálcikaszerű testek jelennek meg a sejtmagban, ami feltehetően egy mutáció miatt megváltozott szerkezetű aktin vagy a citoszkeleton más fehérjéje.[32]
- Cajal-testek. Egy sejtmag átlagosan 1–10 Cajal-testnek vagy spiráltestnek nevezett kompakt struktúrát tartalmaz, melyek átmérője 0,2–2 µm között változik, fajtól és sejttípustól függően.[28] Elektronmikroszkóp alatt gubancos fonalgombolyagnak látszanak[29] és nagy mennyiségű coilin fehérjét tartalmaznak.[33] A feladatuk a különböző RNS-ek (kis nukleoláris RNS, kis nukleáris RNS és a hiszton mRNS) utólagos módosítása.[28]
A Cajal-testek mellett helyezkednek el az iker-spiráltestek (innen kapták a nevüket is), amelyeket mikroszkóp alatt nem is lehet megkülönböztetni tőlük.[33] A Cajal-testektől eltérően azonban nem tartalmaznak kis nukleáris ribonukleoproteint (snRP, a hírvivő RNS utólagos módosítását végzi); funkciójuk feltehetően az snRP szintézisének segítése a Cajal-testekben,[34] de olyan vélemény is létezik, hogy a Cajal-testek és az ikertestek ugyanazon struktúra különböző formái.[33]
- PIKA. A polimorf interfázisbeli karioszomális halmazokat (PIKA, vagy más néven RAFA-domén) először 1991-ben figyelték meg mikroszkóppal. Feladatuk ismeretlen, de valószínűleg nincs közük a DNS másolásához, átírásához vagy az RNS utólagos módosításához.[35] Többnyire a PTF transzkripciós faktor (amely a kis nukleáris RNS, snRNS transzkripcióját segíti) halmazai közelében találhatók.[36]
- PML-testek. A promielocitás leukémia-testek (PML-testek, más néven nukleáris domén 10 vagy Kremer-testek) a nukleoplazmában elszórtan található kerek, 0,1–1 µm átmérőjű szerkezetek. Nevüket fő komponensükről, a promielocitás leukémia proteinről kapták. Többnyire a Cajal-testek vagy spliceoszómák és egyéb hasonló struktúrák közelében láthatók.[30] A PML-testek feltehetően a sejtmag sok fontos funkciójában (DNS-másolás, transzkripció, génműködés-gátlás) van koordináló szerepe, melyet főleg SUMO-val (kis ubiquitin-szerű módosító protein) való összekapcsolással ér el. A PML-hiányos sejtek (amelyekben a génjüket mesterségesen kikapcsolták) nem fejlődnek és funkcionálnak normálisan, azaz feladatuk biztosan alapvető fontosságú.
- Splicing-foltok (splicing speckle, más néven magfolt – nuclear speckle –, interkromatin granulumhalmaz, B-sznörposzóma).[37] A splicing-foltok az emlőssejtek kromatinközi régiójában találhatók, és az mRNS utólagos módosítását, splicingját (ejtsd: szplájszing) végző molekulák halmozódnak fel bennük. Fluoreszcenciamikroszkóp alatt változó méretű és alakú, szabálytalan, pettyezett foltok; elektronmikroszkóppal szemcsés halmaznak látszanak. Szerkezetük és összetételük változó, RNS és fehérjekomponenseik gyakran más helyre vándorolnak, például aktívan átíródó génekhez. A foltok összetétele és elhelyezkedése a génexpresszió változásával összhangban változik.[38]
- Parafoltok (paraspeckle). A 2002-ben felfedezett parafoltok a kromatinközi térben található szabálytalan formájú alakzatok.[39] Először HeLa sejtekben mutatták ki őket, de azután észlelték valamennyi emberi sejttípusban, beleértve a laboratóriumban tenyésztett immortalizált sejtvonalakat.[40] Egy sejtmagban 10–30 van belőlük. Nevüket onnan kapták, hogy a splicing-foltok közelében, azokkal paralel módon helyezkednek el.[41] Nagyságuk és elhelyezkedésük a sejt génexpressziójának függvényében változik. Létük az aktív génátírástól függ, ha a fehérjekódoló géneket átíró RNS-polimeráz II inaktív, akkor valamennyi parafolt eltűnik és fehérjekomponenseik a sejtmagvacskában tárolódnak. Ez a folyamat a sejtosztódás telofázisában is lezajlik, a génátírás ilyenkor is szünetel.[40]
Funkciója
[szerkesztés]A sejtmag elválasztja a gének transzkripciójának helyét a fehérjeszintézistől, a transzlációtól, és ezáltal a génszabályozásnak olyan lehetőségeit nyújtja, amire a prokarióta sejtek nem képesek. A sejtmag fő feladata a génkifejeződés és a DNS-replikáció szabályozása a sejtciklus során.
Génkifejeződés
[szerkesztés]A gének kifejeződése mRNS-sé való átírásukkal indul, majd az mRNS sokféleképpen módosul, és végül a riboszómákhoz kapcsolódva a transzláció révén fehérjék képződnek róla. Az utóbbi folyamat kivételével a többi tevékenység a sejtmagban zajlik.[42] A sejtmagban találhatók a transzkripcióhoz közvetlenül és közvetve szükséges enzimek és az ezeket szabályzó transzkripciós faktorok is. Az enzimek közé tartoznak a helikázok, amelyek szétcsavarják a kettős szálú DNS-t, az RNS-polimerázok, amelyek az mRNS szintézisét végzik és a topoizomerázok, amelyek a DNS széttekeredéséből származó fizikai feszültséget tüntetik el.[43]
Az átírt mRNS-t többféleképpen módosítani kell, mielőtt „érett”, transzlációra alkalmas lesz. Ezek közül a legfontosabbak az 5’cap („sapka”) és az adenozinokból álló poli(A) farok elhelyezése rajta, valamint a splicing. Az 5’cap-et a transzkripciós komplexhez tartozó enzim köti az mRNS-hez, a poli(A) farkat pedig közvetlenül az átírás után kapja meg. A splicing a spliceoszóma nevű komplexen belül zajlik, és hosszú, sok intront tartalmazó mRNS-ek esetében már azelőtt elkezdődhet, hogy a transzkripció befejeződött.[5] Sok esetben a pre-mRNS-t többféleképpen állítják össze érett mRNS-sé, ez az ún alternatív splicing; így keletkeznek például a rendkívül változatos ellenanyagok is.
Kompartmentalizáció
[szerkesztés]A kompartmentalizáció a sejten belüli folyamatok fizikai elválasztását, különböző térrészekbe való elhelyezését jelenti. A maghártya elválasztja a genetikai anyagot a citoplazma többi részétől, és lehetővé teszi, hogy attól elkülönülve kontrollálja a gének átírását. Az egyik általánosan elterjedt módszer szerint a megállítandó folyamat egyik közbenső molekuláját a sejtmagba transzportálják, ahol az transzkripciós faktorokhoz kötődve gátolja az adott reakciót katalizáló enzim génexpresszióját. Ilyen például a sejt energia-háztartásában alapvető glikolízis. Ennek első lépésében a hexokináz a glükózból glukóz-6-foszfátot generál, amiből pedig fruktóz-6-foszfát lesz. Ha az utóbbi koncentrációja túl magas, egy regulátor fehérje a sejtmagba viszi a hexokinázt,[44] ahol az más proteinekkel olyan komplexumot formál, amely meggátolja a glikolízis enzimjeinek kifejeződését.[45]
A génkifejeződést szabályozó transzkripciós faktorokat a sejt sokszor fizikailag elszeparálja a kromoszómáktól, hogy a minimális génátírást is megakadályozza. Ezek a faktorok csak akkor fejthetik ki működésüket, ha a megfelelő szignál aktiválja őket. Például a gyulladási folyamatokban szerepet játszó géneket aktiváló NF-κB faktor a citoplazmában található, amíg a sejt külső membránján lévő receptorokhoz nem csatlakozik valamilyen szignálmolekula (például a TNF-α), és a receptor aktiválja az NF-κB-t, amely ezután a rajta található nukleáris lokalizációs szignál révén a sejtmagba vándorol és megindítja a megfelelő gének transzkripcióját.[6]
A kompartmentalizáció tudja megakadályozni, hogy a génekről frissen átírt, módosítatlan pre-mRNS transzlációja azonnal megkezdődjék.[46] Az eukarióták génjeiben közbeszúrt szakaszok, intronok vannak, amelyeket el kell távolítani, mielőtt fehérjévé fordítanák át. A baktériumoknál a transzláció sokszor már ezelőtt elkezdődik, hogy a hírvivő RNS transzkripciója teljesen befejeződne, eukarióták esetében ez az intronok miatt hibás, használhatatlan fehérjék termelődéséhez vezetne.
Nukleáris transzport
[szerkesztés]
A nagy molekulák mozgása a sejtmagba befelé és onnan kifelé szigorúan szabályozott, és a pórusokon keresztül történik. Míg a kis méretű molekulák szabadon átjárhatnak,[47] a fehérjék és nukleinsavak csak a karioferinnek nevezett proteinhez kapcsolódva haladhatnak át a póruson. A befelé szállító karioferinek az importinok, a sejtmagból kifelé vivők az exportinok. A szállítandó fehérjén rövid aminosav-szekvencia, ún. nukleáris lokalizációs szignál jelzi, hogy végcélja a sejtmag; ugyanez igaz a sejtmagból kiszállítandó polipeptidekre is. A póruson való átszállítás energiája guanozin-trifoszfát (GTP) hasításából származik, amit GTPázok végeznek. A nukleáris transzport legfontosabb GTPáza a Ran nevű protein. Az importinoknak a RanGTP komplexre ahhoz van szükségük, hogy elengedjék szállítmányukat, míg az exportinok esetében a kapcsolódás miatt elengedhetetlen a Ran jelenléte.[11]
Nukleáris import esetén a citoplazmában található fehérjeszállítmányhoz odakötődik az importin és átviszi a póruson keresztül a magba. Odabent az importin a RanGTP segítségével leválasztja magáról a szállítmányt és visszatér a citoplazmába. Az exportálandó fehérjéhez az exportin köt hozzá a RanGTP energiája felhasználásával, átszállítja a póruson, leválik róla és visszatér a sejtmagba.
Az érett mRNS és transzfer RNS (tRNS) kiszállítására külön exportproteinek specializálódtak. Ezek a sejten belüli „minőségellenőrző” rendszer részei, amely megakadályozza hogy a nem vagy csak részben érett mRNS a riboszómákhoz jusson. A rendszer a citoplazmába kijutó, nem teljesen módosított mRNS-t még azelőtt lebontja, hogy elkezdődhetne rajta a transzláció.[5]
A sejtmag lebomlása és újjászerveződése
[szerkesztés]
A sejt életciklusának bizonyos szakaszaiban – a sejtosztódás és a programozott sejthalál, az apoptózis során - a sejtmag megszűnik, és az elkülönülését biztosító struktúrák, a maghártya és a nukleáris lamina tervezetten lebomlik.
A legtöbb sejtben a mitózis profázisának végén a mag membránjai szétesnek. Vannak kivételek, például egyes egysejtű eukarióták (élesztő) ún. zárt mitózis révén osztódnak, amikor maghártyájuk épen marad. Ilyenkor a leánykromoszómák a sejtmag két ellenkező végére vándorolnak, majd a mag osztódik. A fejlettebb eukariótákra azonban a maghártya lebomlásával járó nyílt mitózis jellemző, ahol a leánykromoszómák a mitotikus orsó pólusaihoz mozognak, és új sejtmag jön létre körülöttük.
A sejtosztódás után mindkét leánysejtnek teljes kromoszómakészletre van szüksége, ezért a mitózis előtt a kromoszómák megkettőződnek, majd az osztódás során a mitotikus orsó fonalai (mikrotubulusai) hozzájuk kapcsolódnak, és a leánykromoszómákat a sejt ellentétes végeibe, az orsó pólusához, a centroszómához húzzák. A legtöbb sejtben a centroszómák a magon kívül, a citoplazmában helyezkednek el, így ép maghártya esetén a mikrotubulusok nem férnének hozzá a kromoszómákhoz.[48] Emiatt a mitózis korai szakaszában, a profázisban kezdve és a prometafázisban befejeződve, a sejtmagot körülvevő membránokat szétbontják.[14] Ugyanekkor szétesik a nukleáris lamina is; ez a folyamat a laminfehérjék foszforilálásával jár.[49] A sejtosztódás végén a maghártya újból összeáll, akárcsak az immár defoszforilált laminokból álló lamina.[49]
A dinoflagellátáknál a centroszóma szintén a citoplazmában van, azonban a maghártya ép marad, és a kromoszómák centromerjei beépülnek a sejtmag membránjába, hogy a mikrotubulusok hozzájuk kapcsolódhassanak (ún. extranukleáris orsó). Sok más egysejtűnél (csillósok, sporozoák, egyes gombák) a centroszómák a magon belül találhatók, és a mag nem bomlik fel a sejtosztódáskor.
Az apoptózis a sejt ellenőrzött, előre programozott pusztulása. Ennek során mélyreható változások történnek, a kromatin kondenzálódik, a maghártya és a lamina szétesik. A lamina hálózatának szétbontását a kaszpáz nevű proteázok (fehérjebontó enzimek) végzik, amelyek szétvágják a laminokat. A kaszpázaktikivitást laboratóriumi tesztekben a korai apoptózis kimutatására használják.[14] Léteznek mutáns sejtvonalak, melyek laminja ellenálló a kaszpázokkal szemben, ezekben nem zajlik le a sejtmag apoptotikus bomlása.[14]
A maghártya a vírusok behatolását is meggátolja. Egyes vírusok szaporodásához a sejtmagban található proteinekre van szükség. Egyes DNS-vírusok (például a herpeszvírusok) a sejtmagban szaporodnak és állítják össze új virionjaikat. Ezek úgy szabadulnak ki, hogy lebontják a belső laminát és a maghártyán át bimbózással távoznak, miközben annak egy darabját burokként magukkal viszik.[14]
Mag nélküli és többmagvú sejtek
[szerkesztés]A sejteknek általában egy nukleuszuk van, de egyes esetekben több is lehet, esetleg egy sem. Ez lehet előre tervezett folyamat eredménye (például az emlősök vörösvértestjei) vagy hibás sejtosztódás következménye.
A sejtmag nélküli sejtek nem képesek osztódni. Legismertebb példájuk az emlősök vörösvértestje (eritrocitája), amelyből az egyéb organellumok (pl. mitokondriumok) is hiányoznak, és gyakorlatilag csak az oxigénszállító hemoglobin tartályául szolgálnak. Az eritrociták a vörös csontvelőben való érésük során veszítik el magjukat, riboszómáikat és organellumaikat. A sejtmag akkor tűnik el, amikor a kiindulási eritroblasztból retikulocita (az érett vörösvértest prekurzora) lesz.[50] Egyes mutagének hatására éretlen, mikronukleusszal rendelkező eritrociták is kikerülhetnek a véráramba.[51][52] Mag nélküli sejtek hibás sejtosztódás során is keletkezhetnek, amikor az egyik leánysejtnek két sejtmagja lesz, a másiknak pedig egy sem.
Ezzel szemben a sugárállatkák Acantharea csoportjának tagjai[53] és egyes mikorrhizás gombák több sejtmaggal is rendelkeznek sejtenként,[54] de a Giardia egysejtű bélparazitának is két magja van.[55] Az emberben a harántcsíkolt izom sejtjei, a miociták összeolvadnak és szincíciumot alkotnak; eközben az eredeti sejtmagok megmaradnak.[5] Betegségek és kórképek kísérőjelensége is lehet a többmagvú sejtek megjelenése, például a gyulladás esetén időnként monociták és makrofágok összeolvadásából származó óriás többmagvú sejtek figyelhetők meg,[56] és tumorképződésben is előfordulhatnak.[57] Bizonyos vírusok, például a HIV burokfehérjéi a sejtmembránba épülve a sejtek összeolvadását és sokmagvú szincíciumok kialakulását okozzák.[58]
A dinoflagelláták egyes fajainak két sejtmagva van,[59] azonban ezek genetikai anyaga különbözik: az egyik az eredeti sejtmag, a másik pedig a vele szimbiózisban élő kovamoszaté.
Evolúciója
[szerkesztés]A sejtmag megléte az egyik alapvető különbség a prokarióták és eukarióták között, és kialakulására négy jelentősebb elmélet is született, de általános elfogadottsággal egyik sem bír.[60]
Az első, az ún. szintropikus modell szerint egy archea (ősbaktérium) és egy baktérium szimbiózisából jött létre a magot tartalmazó eukarióta sejt.[61] Az elképzelés szerint a mai metánbaktériumokhoz hasonló archea belsejében parazita vagy szimbionta prokarióta (pl. myxobaktérium) élt, és idővel teljes mértékben alkalmazkodtak egymáshoz. Ez az elmélet analóg a mitokondriumok és kloroplasztiszok bakteriális eredetét hirdető, a szakértők által általánosan elismert endoszimbionta elmélettel.[62] Az ősbaktériumi eredetet támogatja, hogy az eukarióták és archeák génjei (pl. a hisztongének) sok tekintetben hasonlóak. Hasonlóan, a myxobaktériumok is sok olyan tulajdonsággal rendelkeznek (soksejtű komplexumokat alkotnak, mozgékonyak, G-proteinjeik és kinázaik az eukariótákéra hasonlítanak), ami az eukariótákra jellemző.[63]
A második elmélet endoszimbionta állapot nélküli evolúciós átmenetet tételez fel. Például a Planctomycetes baktériumcsoportot hozzák fel, amely membránnal és primitív pórusokkal ellátott sejtmagszerű organellummal rendelkezik.[64] Egy hasonló elképzelés szerint a proto-eukarióta sejt fagocitózissal kebelezte be az archeákat vagy baktériumokat és szerezte meg génjeiket.[65]
A leginkább vitatott modell, a virális eukariogenezis szerint az eukarióták megkülönböztető sajátosságai, közte a membránnal elkülönített sejtmag, vírusfertőzés következményei. Az elmélet alapja az eukarióták és egyes vírusok közötti hasonlóságok: a lineáris kromoszómák, az mRNS 5’cap-ja (sapkája) és a DNS csomagolását végző hisztonok analógiája a vírusok nukleoproteinjeivel. Az elmélet egyik verziója szerint a sejtmag a fagocitózissal párhuzamosan fejlődött ki, és a korai eukarióták egysejtű ragadozók voltak.[66] Egy másik változat szerint az eukarióták himlővírussal fertőzött ősbaktériumtól erednek, aminek az adja az alapját, hogy ezeknek a vírusoknak a DNS-polimeráza hasonlít az eukariótákéra.[67][68] A víruseredettel megpróbálták az ivaros szaporodás tisztázatlan kezdeteit is magyarázni.[69]
A legújabb elmélet, az exomembrán-hipotézis szerint az eukarióták őse egy olyan archea volt, amely egy második membránt fejlesztett az eredeti köré, amelyikből a maghártya lett, és addigi membránkötött struktúráiból kialakultak a pórusok, hogy lehetővé tegyék a belül szintetizált termékei – pl. riboszómák – külső transzportját.[70]
Jegyzetek
[szerkesztés]- ↑ Leeuwenhoek, A. van: Opera Omnia, seu Arcana Naturae ope exactissimorum Microscopiorum detecta, experimentis variis comprobata, Epistolis ad varios illustres viros. J. Arnold et Delphis, A. Beman, Lugdinum Batavorum 1719–1730. Cited after: Dieter Gerlach, Geschichte der Mikroskopie. Verlag Harry Deutsch, Frankfurt am Main, Germany, 2009. ISBN 978-3-8171-1781-9.
- ↑ Harris, H. The Birth of the Cell. New Haven: Yale University Press (1999). ISBN 0-300-07384-4
- ↑ Brown, Robert (1866). „On the Organs and Mode of Fecundation of Orchidex and Asclepiadea”. Miscellaneous Botanical Works I, 511–514. o.
- ↑ a b Cremer, Thomas. Von der Zellenlehre zur Chromosomentheorie. Berlin, Heidelberg, New York, Tokyo: Springer Verlag (1985). ISBN 3-540-13987-7 Online Version here
- ↑ a b c d e f g h Lodish, H. Molecular Cell Biology, 5th, New York: WH Freeman (2004). ISBN 0-7167-2672-6
- ↑ a b c d szerk.: Bruce Alberts, Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, Peter Walter: Molecular Biology of the Cell, Chapter 4, pages 191–234, 4th, Garland Science (2002)
- ↑ Clegg JS (1984. február 1.). „Properties and metabolism of the aqueous cytoplasm and its boundaries”. Am. J. Physiol. 246 (2 Pt 2), R133–51. o. PMID 6364846.
- ↑ a b Paine P, Moore L, Horowitz S (1975). „Nuclear envelope permeability”. Nature 254 (5496), 109–114. o. DOI:10.1038/254109a0. PMID 1117994.
- ↑ szerk.: Rodney Rhoades, Richard Pflanzer: Ch3, Human Physiology, 3rd, Saunders College Publishing (1996)
- ↑ Shulga N, Mosammaparast N, Wozniak R, Goldfarb D (2000). „Yeast nucleoporins involved in passive nuclear envelope permeability”. J Cell Biol 149 (5), 1027–1038. o. DOI:10.1083/jcb.149.5.1027. PMID 10831607. PMC 2174828.
- ↑ a b Pemberton L, Paschal B (2005). „Mechanisms of receptor-mediated nuclear import and nuclear export”. Traffic 6 (3), 187–198. o. DOI:10.1111/j.1600-0854.2005.00270.x. PMID 15702987.
- ↑ Stuurman N, Heins S, Aebi U (1998). „Nuclear lamins: their structure, assembly, and interactions”. J Struct Biol 122 (1–2), 42–66. o. DOI:10.1006/jsbi.1998.3987. PMID 9724605.
- ↑ Goldman A, Moir R, Montag-Lowy M, Stewart M, Goldman R (1992). „Pathway of incorporation of microinjected lamin A into the nuclear envelope”. J Cell Biol 119 (4), 725–735. o. DOI:10.1083/jcb.119.4.725. PMID 1429833. PMC 2289687.
- ↑ a b c d e Goldman R, Gruenbaum Y, Moir R, Shumaker D, Spann T (2002). „Nuclear lamins: building blocks of nuclear architecture”. Genes Dev 16 (5), 533–547. o. DOI:10.1101/gad.960502. PMID 11877373.
- ↑ Moir RD, Yoona M, Khuona S, Goldman RD. (2000). „Nuclear Lamins A and B1: Different Pathways of Assembly during Nuclear Envelope Formation in Living Cells”. Journal of Cell Biology 151 (6), 1155–1168. o. DOI:10.1083/jcb.151.6.1155. PMID 11121432. PMC 2190592.
- ↑ (2002) „Alteration of nuclear lamin organization inhibits RNA polymerase II–dependent transcription”. Journal of Cell Biology 156 (4), 603–608. o. DOI:10.1083/jcb.200112047. PMID 11854306. PMC 2174089.
- ↑ Mounkes LC, Stewart CL (2004). „Aging and nuclear organization: lamins and progeria”. Current Opinion in Cell Biology 16 (3), 322–327. o. DOI:10.1016/j.ceb.2004.03.009. PMID 15145358.
- ↑ Ehrenhofer-Murray A (2004). „Chromatin dynamics at DNA replication, transcription and repair”. Eur J Biochem 271 (12), 2335–2349. o. DOI:10.1111/j.1432-1033.2004.04162.x. PMID 15182349.
- ↑ Grigoryev S, Bulynko Y, Popova E (2006). „The end adjusts the means: heterochromatin remodelling during terminal cell differentiation”. Chromosome Res 14 (1), 53–69. o. DOI:10.1007/s10577-005-1021-6. PMID 16506096.
- ↑ Schardin, Margit (1985. december 1.). „Specific staining of human chromosomes in Chinese hamster x man hybrid cell lines demonstrates interphase chromosome territories”. Human Genetics 71 (4), 281–287. o, Kiadó: Springer Berlin / Heidelberg. [2019. szeptember 13-i dátummal az eredetiből archiválva]. DOI:10.1007/BF00388452. PMID 2416668. (Hozzáférés: 2015. május 29.)
- ↑ Lamond, Angus I. (1998. április 24.). „Structure and Function in the Nucleus”. Science 280 (5363), 547–553. o. DOI:10.1126/science.280.5363.547. PMID 9554838.
- ↑ Kurz, A (1996). „Active and inactive genes localize preferentially in the periphery of chromosome territories”. The Journal of Cell Biology 135 (5), 1195–1205. o, Kiadó: The Rockefeller University Press. [2007. szeptember 29-i dátummal az eredetiből archiválva]. DOI:10.1083/jcb.135.5.1195. PMID 8947544. PMC 2121085. (Hozzáférés: 2015. május 29.)
- ↑ NF Rothfield, BD Stollar (1967). „The Relation of Immunoglobulin Class, Pattern of Antinuclear Antibody, and Complement-Fixing Antibodies to DNA in Sera from Patients with Systemic Lupus Erythematosus”. J Clin Invest 46 (11), 1785–1794. o. DOI:10.1172/JCI105669. PMID 4168731. PMC 292929.
- ↑ S Barned, AD Goodman, DH Mattson (1995). „Frequency of anti-nuclear antibodies in multiple sclerosis”. Neurology 45 (2), 384–385. o. DOI:10.1212/WNL.45.2.384. PMID 7854544.
- ↑ Böhm I. IgG deposits can be detected in cell nuclei of patients with both lupus erythematosus and malignancy. Clin Rheumatol 2007;26(11) 1877-1882
- ↑ Hernandez-Verdun, Daniele (2006). „Nucleolus: from structure to dynamics”. Histochem. Cell. Biol 125 (1–2), 127–137. o. DOI:10.1007/s00418-005-0046-4. PMID 16328431.
- ↑ a b Lamond, Angus I. (2003. október 1.). „Nuclear substructure and dynamics”. current biology 13 (21), R825–828. o. DOI:10.1016/j.cub.2003.10.012. PMID 14588256.
- ↑ a b c Cioce M, Lamond A (2005). „Cajal bodies: a long history of discovery”. Annu Rev Cell Dev Biol 21, 105–131. o. DOI:10.1146/annurev.cellbio.20.010403.103738. PMID 16212489.
- ↑ a b c Pollard, Thomas D.. Cell Biology. Philadelphia: Saunders (2004). ISBN 0-7216-3360-9
- ↑ a b c Dundr, Miroslav (2001). „Functional architecture in the cell nucleus”. Biochem. J. 356 (Pt 2), 297–310. o. DOI:10.1042/0264-6021:3560297. PMID 11368755. PMC 1221839.
- ↑ Fox, Archa. R. Sundby interjúja.. Paraspeckle Size., E-mail Correspondence. 2007. március 7.
- ↑ Goebel, H.H. (1997. január 1.). „Nemaline myopathy with intranuclear rods—intranuclear rod myopathy”. Neuromuscular Disorders 7 (1), 13–19. o. DOI:10.1016/S0960-8966(96)00404-X. PMID 9132135.
- ↑ a b c Matera AG, Frey MA. (1998). „Coiled Bodies and Gems: Janus or Gemini?”. American Journal of Human Genetics 63 (2), 317–321. o. DOI:10.1086/301992. PMID 9683623. PMC 1377332.
- ↑ Matera, A. Gregory (1998). „Of Coiled Bodies, Gems, and Salmon”. Journal of Cellular Biochemistry 70 (2), 181–192. o. DOI:<181::aid-jcb4>3.0.co;2-k 10.1002/(sici)1097-4644(19980801)70:2<181::aid-jcb4>3.0.co;2-k. PMID 9671224.
- ↑ Saunders WS, Cooke CA, Earnshaw WC (1991). „Compartmentalization within the nucleus: discovery of a novel subnuclear region.”. Journal of Cellular Biology 115 (4), 919–931. o. DOI:10.1083/jcb.115.4.919. PMID 1955462
- ↑ Pombo A, Cuello P, Schul W, Yoon J, Roeder R, Cook P, Murphy S (1998). „Regional and temporal specialization in the nucleus: a transcriptionally active nuclear domain rich in PTF, Oct1 and PIKA antigens associates with specific chromosomes early in the cell cycle”. The EMBO Journal 17 (6), 1768–1778. o. DOI:10.1093/emboj/17.6.1768. PMID 9501098. PMC 1170524.
- ↑ Cellular component Nucleus speckle. UniProt: UniProtKB. (Hozzáférés: 2013. augusztus 30.)
- ↑ Handwerger, Korie E. (2006. január 1.). „Subnuclear organelles: new insights into form and function”. TRENDS in Cell Biology 16 (1), 19–26. o. DOI:10.1016/j.tcb.2005.11.005. PMID 16325406.
- ↑ Fox, Archa (2002). „Paraspeckles:A Novel Nuclear Domain”. Current Biology 12 (1), 13–25. o. [2012. december 8-i dátummal az eredetiből archiválva]. DOI:10.1016/S0960-9822(01)00632-7. PMID 11790299. (Hozzáférés: 2015. május 29.)
- ↑ a b Fox, A. (2005). „P54nrb Forms a Heterodimer with PSP1 That Localizes to Paraspeckles in an RNA-dependent Manner”. Molecular Biology of the Cell 16 (11), 5304–5315. o. DOI:10.1091/mbc.E05-06-0587. PMID 16148043. PMC 1266428.
- ↑ Fox, Archa: Nuclear Compartments: Paraspeckles. Nuclear Protein Database, 2004. [2006. május 2-i dátummal az eredetiből archiválva]. (Hozzáférés: 2007. március 6.)
- ↑ Nierhaus, Knud H.. Protein Synthesis and Ribosome Structure: Translating the Genome. Wiley-VCH (2004). ISBN 3-527-30638-2
- ↑ Nicolini, Claudio A.. Genome Structure and Function: From Chromosomes Characterization to Genes Technology. Springer (1997). ISBN 0-7923-4565-7
- ↑ Lehninger, Albert L.. Lehninger principles of biochemistry, 3rd, New York: Worth Publishers (2000). ISBN 1-57259-931-6
- ↑ Moreno F, Ahuatzi D, Riera A, Palomino CA, Herrero P. (2005). „Glucose sensing through the Hxk2-dependent signalling pathway.”. Biochem Soc Trans 33 (1), 265–268. o. DOI:10.1042/BST0330265. PMID 15667322. PMID 15667322
- ↑ Görlich, Dirk (1999). „Transport between the cell nucleus and the cytoplasm”. Ann. Rev. Cell Dev. Biol. 15 (1), 607–660. o. DOI:10.1146/annurev.cellbio.15.1.607. PMID 10611974.
- ↑ Watson, JD. Ch9–10, Molecular Biology of the Gene, 5th, Peason Benjamin Cummings; CSHL Press. (2004). ISBN 0-8053-9603-9
- ↑ Lippincott-Schwartz, Jennifer (2002. március 7.). „Cell biology: Ripping up the nuclear envelope”. Nature 416 (6876), 31–32. o. DOI:10.1038/416031a. PMID 11882878.
- ↑ a b Boulikas T (1995). „Phosphorylation of transcription factors and control of the cell cycle”. Crit Rev Eukaryot Gene Expr 5 (1), 1–77. o. PMID 7549180.
- ↑ Skutelsky, E. (1970. június 1.). „Comparative study of nuclear expulsion from the late erythroblast and cytokinesis”. J Cell Biol 60 (60(3)), 625–635. o. DOI:10.1016/0014-4827(70)90536-7. PMID 5422968.
- ↑ Torous, DK (2000). „Enumeration of micronucleated reticulocytes in rat peripheral blood: a flow cytometric study”. Mutat Res 465 (465(1–2)), 91–99. o. DOI:10.1016/S1383-5718(99)00216-8. PMID 10708974.
- ↑ Hutter, KJ (1982). „Rapid detection of mutagen induced micronucleated erythrocytes by flow cytometry”. Histochemistry 75 (3), 353–362. o. DOI:10.1007/bf00496738. PMID 7141888.
- ↑ Zettler, LA (1997). „Phylogenetic relationships between the Acantharea and the Polycystinea: A molecular perspective on Haeckel's Radiolaria”. Proc Natl Acad Sci USA 94 (21), 11411–11416. o. DOI:10.1073/pnas.94.21.11411. PMID 9326623. PMC 23483.
- ↑ Horton, TR (2006). „The number of nuclei in basidiospores of 63 species of ectomycorrhizal Homobasidiomycetes”. Mycologia 98 (2), 233–238. o. DOI:10.3852/mycologia.98.2.233. PMID 16894968.
- ↑ Adam RD (1991. december 1.). „The biology of Giardia spp”. Microbiol. Rev. 55 (4), 706–32. o. PMID 1779932. PMC 372844.
- ↑ McInnes, A (1988). „Interleukin 4 induces cultured monocytes/macrophages to form giant multinucleated cells”. J Exp Med 167 (2), 598–611. o. DOI:10.1084/jem.167.2.598. PMID 3258008. PMC 2188835.
- ↑ Goldring, SR (1987). „Human giant cell tumors of bone identification and characterization of cell types”. J Clin Invest 79 (2), 483–491. o. DOI:10.1172/JCI112838. PMID 3027126. PMC 424109.
- ↑ Watkins, Brynmor A, Alberta E. „Syncytium formation induced by human immunodeficiency virus type 1 isolates correlates with affinity for CD4.”. Journal of General Virology 78 (10), 2513–2522. o. DOI:10.1099/0022-1317-78-10-2513. ISSN 1465-2099.
- ↑ (2012) „Tertiary endosymbiosis in two dinotoms has generated little change in the mitochondrial genomes of their dinoflagellate hosts and diatom endosymbionts”. PLOS ONE 7 (8), e43763. o. DOI:10.1371/journal.pone.0043763.
- ↑ Pennisi E. (2004). „Evolutionary biology. The birth of the nucleus”. Science 305 (5685), 766–768. o. DOI:10.1126/science.305.5685.766. PMID 15297641.
- ↑ C.Michael Hogan. 2010. Archaea. eds. E.Monosson & C.Cleveland, Encyclopedia of Earth. National Council for Science and the Environment, Washington DC.
- ↑ Margulis, Lynn. Symbiosis in Cell Evolution. San Francisco: W. H. Freeman and Company, 206–227. o. (1981). ISBN 0-7167-1256-3
- ↑ Lopez-Garcia P, Moreira D. (2006). „Selective forces for the origin of the eukaryotic nucleus”. BioEssays 28 (5), 525–533. o. DOI:10.1002/bies.20413. PMID 16615090.
- ↑ Fuerst JA. (2005). „Intracellular compartmentation in planctomycetes”. Annu Rev Microbiol. 59, 299–328. o. DOI:10.1146/annurev.micro.59.030804.121258. PMID 15910279.
- ↑ Hartman H, Fedorov A. (2002). „The origin of the eukaryotic cell: a genomic investigation”. Proc Natl Acad Sci U S A. 99 (3), 1420–1425. o. DOI:10.1073/pnas.032658599. PMID 11805300. PMC 122206.
- ↑ Bell PJ (2001. szeptember 1.). „Viral eukaryogenesis: was the ancestor of the nucleus a complex DNA virus?”. J. Mol. Evol. 53 (3), 251–6. o. DOI:10.1007/s002390010215. PMID 11523012.
- ↑ Takemura M (2001). „Poxviruses and the origin of the eukaryotic nucleus”. J Mol Evol 52 (5), 419–425. o. DOI:10.1007/s002390010171. PMID 11443345.
- ↑ Villarreal L, DeFilippis V (2000). „A hypothesis for DNA viruses as the origin of eukaryotic replication proteins”. J Virol 74 (15), 7079–7084. o. DOI:10.1128/JVI.74.15.7079-7084.2000. PMID 10888648. PMC 112226.
- ↑ Bell PJ (2006. november 1.). „Sex and the eukaryotic cell cycle is consistent with a viral ancestry for the eukaryotic nucleus”. J. Theor. Biol. 243 (1), 54–63. o. DOI:10.1016/j.jtbi.2006.05.015. PMID 16846615.
- ↑ de Roos AD (2006). „The origin of the eukaryotic cell based on conservation of existing interfaces”. Artif Life 12 (4), 513–523.. o. DOI:10.1162/artl.2006.12.4.513. PMID 16953783.
Fordítás
[szerkesztés]- Ez a szócikk részben vagy egészben a Cell nucleus című angol Wikipédia-szócikk ezen változatának fordításán alapul. Az eredeti cikk szerkesztőit annak laptörténete sorolja fel. Ez a jelzés csupán a megfogalmazás eredetét és a szerzői jogokat jelzi, nem szolgál a cikkben szereplő információk forrásmegjelöléseként.
További információk
[szerkesztés]- Goldman, Robert D. (2002). „Nuclear lamins: building blocks of nuclear architecture”. Genes & Dev. 16 (5), 533–547. o. DOI:10.1101/gad.960502. PMID 11877373.
- Görlich, Dirk (1999). „Transport between the cell nucleus and the cytoplasm”. Ann. Rev. Cell Dev. Biol. 15, 607–660. o. DOI:10.1146/annurev.cellbio.15.1.607. PMID 10611974.
- Lamond, Angus I. (1998. április 24.). „Structure and Function in the Nucleus”. Science 280 (5363), 547–553. o. DOI:10.1126/science.280.5363.547. PMID 9554838.
- Pennisi E. (2004). „Evolutionary biology. The birth of the nucleus”. Science 305 (5685), 766–768. o. DOI:10.1126/science.305.5685.766. PMID 15297641.
- Pollard, Thomas D.. Cell Biology. Philadelphia: Saunders (2004). ISBN 0-7216-3360-9
- sejtmag (magyar nyelven). Sulinet - Sebőkné Orosz Katalin. [2013. szeptember 14-i dátummal az eredetiből archiválva]. (Hozzáférés: 2015. június 21.)