Astrocito
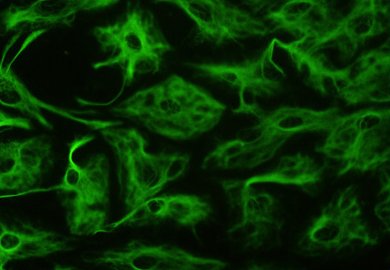




Os astrocitos, tamén chamados colectivamente astroglía, son células de glía cunha característica forma estrelada (de onde lles vén o nome), que se encontran no encéfalo e medula espiñal. Son as célula máis abundantes do cerebro humano. Realizan moitas funcións, como colaboraren bioquimicamente coas células endoteliais da barreira hematoencefálica, a provisión de nutrientes ao tecido nervioso, o mantemento do equilibrio iónico extracelular, e interveñen na reparación e proceso de cicatrización dos tecidos danados do cerebro e medula espiñal.
Diversas investigacións realizadas desde mediados da década de 1990 atoparon que os astrocitos propagan ondas de Ca2+ intercelulares a longas distancias en resposta á estimulación, e, igual que as neuronas, poden liberar certos transmisores (chamados gliotransmisores) dun modo dependente do Ca2+. Os datos suxiren que os astrocitos tamén envían sinais ás neuronas por medio da liberación de glutamato dependente de Ca2+.[1] Estes descubrimentos fixeron que os astrocitos sexan unha importante área de investigación en neurociencia.
Descrición
[editar | editar a fonte]Os astrocitos son un subtipo de células gliais no sistema nervioso central. Teñen forma estrelada con moitos procesos celulares que envolven as sinapses que realizan as neuronas. Os astrocitos identificáronse clasicamente por análise histolóxica; moitas destas células expresan o filamento intermedio proteína acídica fibrilar glial (GFAP). Existen varias formas de astrocitos no sistema nervioso central, entre os que están os fibrosos, protoplásmicos, e radiais. A glía fibrosa está xeralmente localizada na substancia branca, ten relativamente poucos orgánulos, e presenta procesos celulares longos e non ramificados. Este tipo con frecuencia presenta "pés vasculares" que conectan fisicamente as células da parte externa da parede do capilar cando están moi próximas entre elas. A glía protoplásmica é a máis común e atópase na substancia gris, posúe unha gran cantidade de orgánulos, e mostra procesos celulares terciarios curtos e moi ramificados. A glia radial está disposta nun plano perpendicular ao eixe dos ventrículos cerebrais. Un dos seus procesos chega á pía máter, e o outro introdúcese profundamente na substancia gris. A glía radial está principalmente presente durante o desenvolvemento, e xoga un papel na migración neuronal. As células de Mueller da retina e as células de glía de Bergmann do córtex do cerebelo son unha excepción, porque están presentes na vida adulta. Cando están próximos á pía máter, as tres formas de astrocitos envían procesos que forman a membrana pia-glial.
Funcións
[editar | editar a fonte]Inicialmente, en medicina considerábase que no tecido nervioso a rede neuronal era o único importante, e os astrocitos (e a glía en xeral) eran vistos como células que "enchían os espazos" e unían o tecido. Máis recentemente, a función dos atrocitos foi reconsiderada,[2] e agora cóidase que desempeñan varias funcións activas no cerebro, como a secreción e absorción de transmisores neurais e o mantemento da barreira hematoencefálica.[3] Propúxose tamén o concepto de "sinapse tripartita", que se refire á estreita relación que hai nas sinapses entre o elemento presináptico, o elemento postsináptico e o elemento glial.[4] As funcións dos astrocitos son as seguintes:
- Estrutural: Están implicados na estrutura física do cerebro. Son as células gliais máis abundantes do cerebro que están en estreita asociación coas sinapses neuronais. Regulan a transmisión de impulsos eléctricos no cerebro.
- Apoio metabólico: Fornecen ás neuronas nutrientes como o lactato.
- Barreira hematoencefálica: Os pés terminais dos astrocitos que rodean as células endoteliais pénsase que axudan ao mantemento da barreira hematoencefálica, pero algúns estudos recentes indican que non xogan un papel substancial; senón que son as unións herméticas e a lámina basal das células do endotelio cerebral as que desempeñan o papel fundamental no mentemento da barreira.[5] Porén, viuse recentemente que a actividade dos astrocitos está ligada co fluxo sanguíneo do cerebro.[6]
- Captación e liberación de transmisores: Os astrocitos expresan na membrana plasmática transportadores como os transportadores de glutamato para varios neurotransmisores, como o glutamato, ATP, e GABA. Máis recentemente, viuse que os astrocitos liberan glutamato ou ATP en vesículas dun modo dependente do Ca2+.[7] (Pero isto foi discutido para os astrocitos do hipocampo.)[8]
- Regulación da concentración iónica extracelular: Os astrocitos expresan unha gran densidade de canles de potasio. Cando as neuronas están activas, liberan ión potasio, o que incrementa a súa concentración extracelular. Como os astrocitos son moi permeables ao potasio, rapidamente o absorben e evitan a súa acumulación en exceso no espazo extracelular. Se algo interfire esta función, a concentración extracelular de potasio aumenta, orixinando a despolarización neuronal.
- Modulación da transmisión sináptica: No núcleo supraóptico do hipotálamo, observouse que cambios rápidos na morfoloxía dos astrocitos afectan á transmisión heterosináptica entre neuronas.[9] No hipocampo, os astrocitos suprimen a transmisión sináptica ao liberaren ATP, o cal é hidrolizado por ectonucleotidases rendendo adenosina. A adenosina actúa sobre o receptor de adenosina neuronal inhibindo a transmisión sináptica, o que incrementa o rango dinámico dispoñible para a potenciación a longo prazo.[10]
- Vasomodulación: Os astrocitos poden servir como intermediarios na regulación neuronal do fluxo sanguíneo.[11]
- Promoción da actividade mielinizante dos oligodendrocitos: A actividade eléctrica nas neuronas causa que estas liberen ATP, o cal serve como un importante estímulo para que se forme a mielina. Porén, o ATP non actúa directamente sobre os oligodendrocitos, senón que fai que os astrocitos secreten a citocina factor inhibidor da leucemia (LIF), unha proteína regulatoria que promove a actividade mielinizante dos oligodendrocitos. Isto suxire que os astrocitos teñen un papel coordinador executivo no cerebro.[12]
- Reparación do sistema nervioso: Nas lesións do tecido nervioso no sistema nervioso central os astrocitos enchen o espazo e forman unha cicatriz glial, reparando a área e substituíndo as células do sistema nervioso central que non se poden rexenerar.[13]
- Potenciación a longo prazo: Discútese se os astrocitos integran ou non a aprendizaxe e a memoria no hipocampo. Sábese que as células gliais están incluídas nas sinapses neuronais, pero moitos dos estudos sobre potenciación a longo prazo realízanse en cortes do cerebro, e non hai un acordo sobre se os astrocitos teñen ou non un papel directo na modulación da plasticidade sináptica.
Ondas de calcio
[editar | editar a fonte]Os astrocitos están unidos por unións comunicantes, que crean un sincitio funcional conectado electricamente.[14]
Un incremento nas concentracións de calcio intracelular pode propagarse cara ao exterior a través deste sincitio funcional. Os mecanismos de propagación da onda de calcio inclúen a difusión dos ións calcio e IP3 a través das unións comunicantes e unha sinalización extracelular de ATP.[15] As elevacións dos niveis de calcio son o principal factor coñecido na activación dos astrocitos, e son necesarios e suficientes para algúns tipos de liberación de glutamato astrocítica.[16]
Desenvolvemento
[editar | editar a fonte]Os astrocitos son células de macroglía derivadas de poboacións heteroxéneas de células proxenitoras no neuroepitelio do sistema nervioso central en desenvolvemento. Traballos recentes [17] indican que hai unha gran similitude entre os mecanismos xenéticos que especifican a liñaxe de diversas subtipos de neuronas e os das células da macroglía. Ademais da especificación celular neuronal, factores de sinalización clásicos como o SHH (sonic hedgehog), factor de crecemento dos fibroblastos (FGFs), WNTs e proteína morfoxenética ósea (BMP), proporcionan información posicional ás células da macroglía en desenvolvemento por medio de gradientes morfóxenos ao longo dos eixes dorsoventral, anteroposterior e mediolateral. Os patróns resultantes ao longo do neuroeixe levan á segmentación do neuroepitelio en dominios proxenitores (p0, p1 p2, p3 e pMN) para distintos tipos de neuronas na medula espiñal en desenvolvemento. Baseándose en varios estudos, hoxe pénsase que este modelo tamén se aplica á especificación das células macrogliais. Algúns estudos demostraron que se orixinan tres poboacións distintas de astrocitos a partir dos dominios p1, p2 e p3 [18]. Estes subtipos de astrocitos poden ser identificados pola expresión de diferentes factores de transcrición (PAX6, NKX6.1) e marcadores da superficie celular (reelina e SLIT1). As tres poboacións de astrocitos que foron identificadas son: 1) astrocitos VA1 localizados dorsalmente, derivados do dominio p1, que expresan PAX6 e reelina; 2) astrocitos VA3 localizados ventralmente, derivados de p3, que expresan NKX6.1 e SLIT1, e 3) astrocitos VA2 intermedios localizados na substancia branca, derivados do dominio p2, que expresan PAX6, NKX6.1, reelina e SLIT1[19]. Unha vez que ten lugar a especificación do astrocito no sistema nervioso central en desenvolvemento, crese que os precursores dos astrocitos migran ás súas posicións finais no sistema nervioso central antes de que ocorra o proceso terminal de diferenciación celular.
Clasificación
[editar | editar a fonte]Hai diferentes modos de clasificar aos astrocitos.
Liñaxe e fenotipo antixénico
[editar | editar a fonte]Esta clasificación foi establecida polos trraballos clásicos de Raff et al. a comezos da década de 1980 nos nervios ópticos da rata.
- Tipo 1: Antixenicamente é Ran2+, GFAP+, FGFR3+, A2B5-. Poden orixinarse a partir de células preursoras gliais tripotenciais (GRP), pero non a partir de células O2A/OPC bipotenciais (oligodendrocito, precursor do astrocito tipo 2, tamén chamadas células precursoras do oligodendrocito).
- Tipo 2: Antixenicamente é A2B5+, GFAP+, FGFR3-, Ran 2-. Poden desenvolverse in vitro a partir tanto de GRP tripotencial (probablemente no estado O2A) ou de células bipotenciais O2A (ás cales á súa vez poderían ter derivado de GRP) ou in vivo cando estas células proxenitoras son transplantadas ao sitio dunha lesión (pero probablemente non no desenvolvemento normal, polo menos non no nervio óptico da rata). Os astrocitos tipo 2 son o principal compoñente astrocítico nos cultivos de nervio óptico postnatal que son xerados por células O2A que creceron en presenza de soro de tenreira fetal pero non se cre que existan in vivo. (Fulton et al., 1992).
Clasificación anatómica
[editar | editar a fonte]- Protoplásmicos. Situados na substancia gris. Presentan moitos procesos ramificados cuxos pés terminais envolven as sinapses. Algúns astrocitos protoplásmicos xéranse a partir de células proxenitoras da zona subventricular multipotentes.[20][21]
- Astrocitos Gömöri positivos. Son un subconxunto de astrocitos protoplásmicos que conteñen numerosas inclusións citoplasmáticas ou gránulos, que se tinguen positivamente coa tinguidura de hematoxilina cromo-alum de Gömöri. Estes gránulos están formados dos restos de mitocondrias dexeneradas englobados en lisosomas,[22] Certos tipos de estrés oxidativo parecen ser responsables dos danos nas mitocondrias neste tipo de astrocitos especializados. Os astrocitos Gömöri positivos son moito máis abundantes no núcleo arqueado do hipotálamo e no hipocampo ca noutras rexións. Poden ter un papel na regulación da resposta do hipotálamo á glicosa.[23][24]
- Fibrosos. Situados na substancia branca. Teñen procesos longos e finos non ramificados cuxos pés terminais envolven os nódulos de Ranvier. Algúns astrocitos fibrosos xéranse pola glía radial.[25][26][27][28][29]
Clasificación polo transportador/receptor
[editar | editar a fonte]Distínguense dous tipos:
- Tipo GluT. Expresa os transportadores de glutamato (EAAT1 e EAAT2).
- Tipo GluR. Expresa receptores de glutamato (principalmente o mGluR e de tipo AMPA).
Patoloxía
[editar | editar a fonte]Os astrocitomas son tumores primariamente intracraniais derivados de astrocitos. Tamén é posible que as células proxenitoras gliais ou as células nai neurais dean lugar a astrocitomas. Os astrocitomas poden aparecer en calquera parte do encéfalo e ás veces noutras partes do sistema nervioso central, como na medula espiñal.[30][31]
Notas
[editar | editar a fonte]- ↑ Fiacco TA, Agulhon C, McCarthy KD (2008). "Sorting out Astrocyte Physiology from Pharmacology". Annu. Rev. Pharmacol. Toxicol. 49 (1): 151–74. PMID 18834310. doi:10.1146/annurev.pharmtox.011008.145602.
- ↑ "The Brain From Top To Bottom". Thebrain.mcgill.ca. Consultado o 2008-11-29.
- ↑ Kolb & Whishaw: Fundamentals of Human Neuropsychology, 2008
- ↑ Araque A, Parpura V, Sanzgiri RP, Haydon PG (1999). "Tripartite synapses: glia, the unacknowledged partner.". Trends in Neuroscience 22 (5): 208–215. PMID 10322493. doi:10.1016/S0166-2236(98)01349-6.
- ↑ Kimelberg HK, Jalonen T, Walz W. (1993). "Regulation of the brain microenvironment: transmitters and ions.". En Murphy, S. Astrocytes: pharmacology and function. San Diego, CA: Academic Press. pp. 193–222.
- ↑ Swaminathan N (2008). "Brain-scan mystery solved". Scientific American Mind. Oct-Nov: 7.
- ↑ Santello M, Volterra A (2008). "Synaptic modulation by astrocytes via Ca(2+)-dependent glutamate release.". Neuroscience. Mar 158 (1): 253–9. PMID 18455880. doi:10.1016/j.neuroscience.2008.03.039.
- ↑ Agulhon C, Fiacco T, McCarthy K (2010). "Hippocampal short- and long-term plasticity are not modulated by astrocyte Ca2+ signaling.". Science 327 (5970): 1250–1257. PMID 20203048. doi:10.1126/science.1184821.
- ↑ Piet R, Vargová L, Syková E, Poulain D, Oliet S (2004). "Physiological contribution of the astrocytic environment of neurons to intersynaptic crosstalk". Proc Natl Acad Sci USA 101 (7): 2151–5. PMC 357067. PMID 14766975. doi:10.1073/pnas.0308408100.
- ↑ Pascual O, Casper KB, Kubera C, Zhang J, Revilla-Sanchez R, Sul JY, Takano H, Moss SJ, McCarthy K, Haydon PG (2005). "Astrocytic purinergic signaling coordinates synaptic networks". Science 310 (5745): 113–6. PMID 16210541. doi:10.1126/science.1116916.
- ↑ Parri R, Crunelli V (2003). "An astrocyte bridge from synapse to blood flow". Nat Neurosci 6 (1): 5–6. PMID 12494240. doi:10.1038/nn0103-5.
- ↑ Ishibashi T, Dakin K, Stevens B, Lee P, Kozlov S, Stewart C, Fields R (2006). "Astrocytes Promote Myelination in Response to Electrical Impulses". Neuron 49 (6): 823–32. PMC 1474838. PMID 16543131. doi:10.1016/j.neuron.2006.02.006.
- ↑ Fawcett JW, Asher RA. The glial scar and central nervous system repair. Brain Res Bull. 1999 Aug;49(6):377-91. [1] PMID 10483914
- ↑ Bennett M, Contreras J, Bukauskas F, Sáez J (2003). "New roles for astrocytes: gap junction hemichannels have something to communicate". Trends Neurosci 26 (11): 610–7. PMID 14585601. doi:10.1016/j.tins.2003.09.008.
- ↑ Newman, EA.(2001)"Propagation of intercellular calcium waves in retinal astrocytes and Müller cells."J Neurosci. 21(7):2215-23
- ↑ Parpura V, Haydon P (2000). "Physiological astrocytic calcium levels stimulate glutamate release to modulate adjacent neurons". Proc Natl Acad Sci USA 97 (15): 8629–34. PMC 26999. PMID 10900020. doi:10.1073/pnas.97.15.8629.
- ↑ Rowitch, David H.; Kriegstein, Arnold R. (2010). "Developmental genetics of vertebrate glial–cell specification". Nature 468 (7321): 214–22. PMID 21068830. doi:10.1038/nature09611.
- ↑ Muroyama, Y; Fujiwara, Y; Orkin, SH; Rowitch, DH (2005). "Specification of astrocytes by bHLH protein SCL in a restricted region of the neural tube". Nature 438 (7066): 360–363. PMID 16292311. doi:10.1038/nature04139.
- ↑ Hochstim, C; Deneen, B; Lukaszewicz, A; Zhou, Q; Anderson, DJ (2008). "Identification of positionally distinct astrocyte subtypes whose identities are specified by a homeodomain code". Cell 133 (3): 510–522. PMC 2394859. PMID 18455991. doi:10.1016/j.cell.2008.02.046.
- ↑ Levison SW, Goldman JE (1993). "Both oligodendrocytes and astrocytes develop from progenitors in the subventricular zone of postnatal rat forebrain". Neuron 10 (2): 201–12. PMID 8439409. doi:10.1016/0896-6273(93)90311-E.
- ↑ Zerlin M, Levison SW, Goldman JE (1995). "Early patterns of migration, morphogenesis, and intermediate filament expression of subventricular zone cells in the postnatal rat forebrain". J. Neurosci. 15 (11): 7238–49. PMID 7472478.
- ↑ Brawer JR; Stein, Robert; Small, Lorne; Ciss, Soriba; Schipper, Hyman M. (1994). "Composition of Gomori-positive inclusions in astrocytes of the hypothalamic arcuate nucleus". Anatomical Record 240 (3): 407–415. PMID 7825737. doi:10.1002/ar.1092400313.
- ↑ Young JK, McKenzie JC (2004) "GLUT2 immunoreactivity in Gömöri-positive astrocytes of the hypothalamus Arquivado 20 de agosto de 2008 en Wayback Machine.."J. Histochemistry & Cytochemistry 52: 1519-1524 PMID
- ↑ Marty N (2005). "Regulation of glucagon secretion by glucose transporter type 2 (glut2) and astrocyte-dependent glucose sensors". J. Clinical Investigation 115: 3545.
- ↑ Choi BH, Lapham LW (1978). "Radial glia in the human fetal cerebrum: a combined Golgi, immunofluorescent and electron microscopic study". Brain Res. 148 (2): 295–311. PMID 77708. doi:10.1016/0006-8993(78)90721-7.
- ↑ Schmechel DE, Rakic P (1979). "A Golgi study of radial glial cells in developing monkey telencephalon: morphogenesis and transformation into astrocytes". Anat. Embryol. 156 (2): 115–52. PMID 111580. doi:10.1007/BF00300010.
- ↑ Misson JP, Edwards MA, Yamamoto M, Caviness VS (1988). "Identification of radial glial cells within the developing murine central nervous system: studies based upon a new immunohistochemical marker". Brain Res. Dev. Brain Res. 44 (1): 95–108. PMID 3069243. doi:10.1016/0165-3806(88)90121-6.
- ↑ Voigt T (1989). "Development of glial cells in the cerebral wall of ferrets: direct tracing of their transformation from radial glia into astrocytes". J. Comp. Neurol. 289 (1): 74–88. PMID 2808761. doi:10.1002/cne.902890106.
- ↑ Goldman SA, Zukhar A, Barami K, Mikawa T, Niedzwiecki D (1996). "Ependymal/subependymal zone cells of postnatal and adult songbird brain generate both neurons and nonneuronal siblings in vitro and in vivo". J. Neurobiol. 30 (4): 505–20. PMID 8844514. doi:10.1002/(SICI)1097-4695(199608)30:4<505::AID-NEU6>3.0.CO;2-7.
- ↑ Astrocytomas. (2010). Retrieved 2011, from IRSA: http://www.irsa.org/astrocytoma.html Arquivado 05 de abril de 2012 en Wayback Machine.
- ↑ Astrocytoma Tumors (2005, August). Retrieved 2011, from American Association of Neurological Surgeons: http://www.aans.org/Patient%20Information/Conditions%20and%20Treatments/Astrocytoma%20Tumors.aspx Arquivado 21 de xullo de 2011 en Wayback Machine..
Véxase tamén
[editar | editar a fonte]Bibliografía
[editar | editar a fonte]- Halassa, M.M., Fellin, T., and Haydon, P.G. (2006). "The tripartite synapse: roles for gliotransmission in health and disease". Trends in Mol. Sci 13 (2): 54–63. PMID 17207662. doi:10.1016/j.molmed.2006.12.005.
- White, F.A., Jung, H. and Miller, R.J. (2007). "Chemokines and the pathophysiology of neuropathic pain". Proc. Natl Acad. Sci. USA 104 (51): 20151–20158. PMC 2154400. PMID 18083844. doi:10.1073/pnas.0709250104.
- Milligan, E.D. and Watson, L.R. (2009). "Pathological and protective roles of glia in chronic pain". Neuron-Glia Interactions 10: 23–36.
- Watkins, L.R., Milligan, E.D. and Maier, S.F. (2001). "Glial activation: a driving force for pathological pain". Trends in Neurosci. 24 (8): 450–455. doi:10.1016/S0166-2236(00)01854-3.
- Volterra, A. and Meldolesi, J. (2005). "Astrocytes, from brain glue to communication elements: the revolution continues". Nat. Rev Neurosci. 6 (8): 626–640. PMID 16025096. doi:10.1038/nrn1722.
- Freeman, M. R. (2010). "Specification and Morphogenesis of Astrocytes". Science 330 (6005): 774–8. PMID 21051628. doi:10.1126/science.1190928.
Ligazóns externas
[editar | editar a fonte]- Cell Centered Database - Astrocyte
- UIUCHistologySubject - 57 [2]
- "Astrocytes" Arquivado 21 de novembro de 2012 en Wayback Machine. at Society for Neuroscience
- The Department of Neuroscience at Wikiversity
- NIF Search - Astrocyte via the Neuroscience Information Framework
| Control de autoridades |
|
|---|