Lõigustumine
Lõigustumine on sügoodi jagunemine mitoosi teel ilma rakkude kasvamiseta. Lõigustumise käigus taastatakse organismi hulkraksus ning tuuma-tsütoplasma suhe. Lõigustumise tulemusena moodustub kõigepealt kobarloode ehk moorula, millest hiljem areneb blastula.[1]
Embrüoloogias tähendab lõigustumine varajase embrüo rakkude jagunemist. Paljude liikide sügoodid teevad läbi rakujagunemise ilma kasvamiseta, mille käigus tekib rakkude kogum, mis on sama suurusega kui algne sügoot. Lõigustumise tulemusena tekivad rakud, mida nimetatakse blastomeerideks. Blastomeerid moodustavad tiheda rakumassi ehk moorula. Lõigustumine lõppeb blastula moodustamisega.
Lõigustumine erineb teistest rakujagunemise vormidest sellepoolest, et lõigustumise käigus suureneb rakkude arv ilma rakumassi suurenemiseta. See tähendab seda, et iga õnnestunud jagunemise käigus tuuma suhe tsütoplasma hulgaga võrreldes kasvab.
Kiire rakutsükkel tagatakse kõrge proteiinide taseme säilitamisega, mille ülesanne on kontrollida rakutsükli kulgu. Sellisteks valkudeks on tsükliinid ja nendega seotud tsükliinisõltuvad kinaasid (cdk). CyclinB/cdc2 kompleks on tuntud ka kui MPF, mis soodustab mitoosi sisenemist. Lõigustumise tagamiseks töötavad karüokineesi (mitoos) ja tsütokineesi protsessid koos. Mitoos saab toimuda tsütokineesi puudusel. Tsütokinees aga ilma mitoosita toimuda ei saa. Lõigustumise lõpp langeb kokku sügoodi transkriptsiooni algusega. Seda punkti nimetatakse midblastula transitsiooniks ja see on kontrollitud tuuma-tsütoplasma suhtega (1/6).[2]
Muna koosneb munarakust ja seda ümbritsevatest kestadest. Küpsele munale on iseloomulik animaalne ja vegetatiivne poolus. Muna poolust, milles on palju rebu, nimetatakse vegetatiivseks pooluseks. Vastaspoolus, milles on vähem rebu on animaalne poolus. Sõltuvalt rebu hulgast, mida muna sisaldab, võib lõigustumise jagada holoblastiliseks (täielik lõigustumine) ja meroblastiliseks (osaline lõigustumine).
Lõigustumine võib olla sünkroonne või asünkroonne sõltuvalt sellest, kas blastomeerid jagunevad üheaegselt või erineval ajal. Lõigustumine võib olla täielik või osaline, sõltuvalt ootsüüdis sisalduvast rebuhulgast. Kui rebuhulk on suur, siis toimub osaline lõigustumine. Vähese või keskmise rebuhulga korral toimub totaalne jagunemine. Holoblastiliseks nimetatakse munarakke, mis lõigustuvad täielikult. Osaliselt lõigustuvad munarakud on meroplastilised.[1]
Asünkroonne ja sünkroonne lõigustumine
[muuda | muuda lähteteksti]Lõigustumine võib olla sünkroonne või asünkroonne. Selline jaotus sõltub sellest, kas blastomeerid jagunevad üheaegselt või erineval ajal. Enamikul loomarühmadel on lõigustumise algus sünkroonne. Hiljem muutub lõigustumine aga asünkroonseks. Imetajatel on esimene lõigustumine asünkroonne. See tähendab seda, et kaks esimest blastomeeri alustavad kohe alguses asünkroonset lõigustumist. Sünkroonne lõigustumine toimub kiiremini kui asünkroonne lõigustumine. Embrüoloogias on sünkroonse lõigustumise pikkus oluline eri liikide arengustaadiumide ajalisel võrdlemisel. Sünkroonse lõigustumise pikkus ei sõltu keskkonnast.[1]
Blastulatsioon
[muuda | muuda lähteteksti]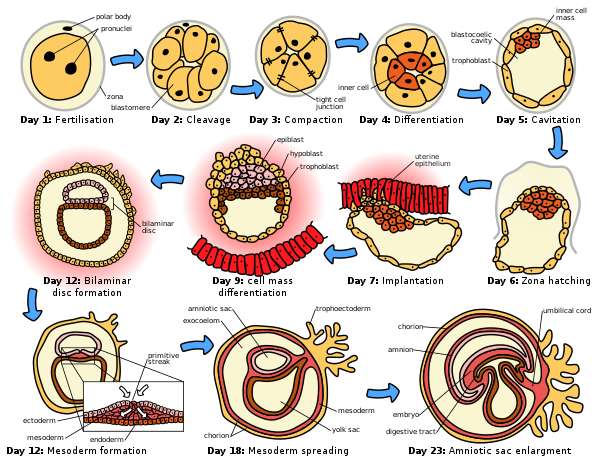
Eri liikidel muutub DNA replikatsioon teatud rakutsüklite järel asünkroonseks, sest rakujagunemised ei toimu enam ühel ajal. Blastulatsiooniks nimetatakse hetke, kui embrüo läheb asünkroonsesse lõigustumise faasi. Blastulatsiooni käigus tekib embrüo sisse blastotsööl. Lõigustumist saab jaotada ka blastomeeride suuruse järgi. Kui täieliku lõigustumise korral on blastomeerid ühesuurused, siis on tegu totaal-ekvaalse lõigustumisega. Totaal-adekvaalne lõigustumine esineb siis, kui animaalse pooluse rakud on veidi väiksemad ja blastomeerid on natuke erineva suurusega. Kui täielikul lõigustumisel on blastomeerid erineva suurusega, siis on tegu totaal-inekvaalse lõigustumisega. Sellise lõigustumise puhul nimetatakse muna animaalse pooluse rakke mikromeerideks ja vegetatiivse pooluse rakke makromeerideks.[1]
Holoblastiline lõigustumine
[muuda | muuda lähteteksti]Holoblastiline lõigustumine esineb täielikult lõigustuva munaraku puhul. Holoblastiline lõigustumine jaguneb omakorda veel radiaalseks, spiraalseks, bilateraalseks ja rotatsiooniliseks lõigustumiseks.
Radiaalne lõigustumine
[muuda | muuda lähteteksti]Radiaalne lõigustumine on iseloomulik okasnahksetele. Lõigustumise tulemusena tekib tsöloblastula. Esimesed kaks lõigustumist toimuvad vertikaaltelje suunas ning kolmas lõigustumine toimub horisontaaltasapinnas. Lõigustumistasapinnad on üksteise suhtes risti. Kahe esimese lõigustumise tulemusena on tekkinud neli ühesuurust blastomeeri. Kolmanda lõigustumise tulemusena tekib kaheksa enam-vähem ühesugust blastomeeri. Kahepaiksete radiaalne lõigustumine sarnaneb okasnahksete lõigustumisega, kuid kuna kahepaiksete munarakk on reburohke, siis jääb vegetatiivse pooluse lõigustumine ajaliselt maha animaalse pooluse lõigustumisest. Selle tulemusena moodustuvad animaalsele poolusele mikromeerid ja vegetatiivsele poolusele makromeerid.
Spiraalne lõigustumine
[muuda | muuda lähteteksti]Selline lõigustumise tüüp on iseloomulik paljudele selgrootutele. Spiraalse lõigustumise tulemusena tekib väikese blastotsööliga stereoblastula. Nimetus spiraalne lõigustumine tuleb sellest, et lõigustumise käigus pöörduvad mikromeeride käävid makromeeri käävide suhtes 45 kraadi võrra. Käävi pöörde suund (päripäeva või vastupäeva) määrab ära selle, millise lõigustumise suunaga on loom. Kahe esimese lõigustumise järel on blastomeerid enam-vähem ühesuguse suurusega. Peale kolmandat jagunemist saab eristada mikromeere animaalsel poolusel ja makromeere vegetatiivsel poolusel.
Bilateraalne lõigustumine
[muuda | muuda lähteteksti]Bilateraalne lõigustumine on iseloomulik tunikaatidele. Bilateraalse lõigustumise käigus toimub mõlemal poolusel (animaalsel/vegetatiivsel) lõigustumine peegelpildi järgi.
Rotatsiooniline lõigustumine
[muuda | muuda lähteteksti]Rotatsioonilise lõigustumise tüüp on iseloomulik imetajatele. Rotatsiooniline lõigustumine tähendab seda, et esimesed kaks blastomeeri, mis lõigustumise käigus tekivad, moodustavad edasises lõigustumises lõigustumistasapinnad eri suundades. Ühel blastomeeril on vertikaalsuunaline lõigustumine ja teisel horisontaalne. Kuni 8-raku staadiumini on lõigustumise käigus tekkinud blastomeerid teineteisega nõrgalt seotud, peale seda toimub kompakteerumine ehk tihendamine, mille käigus blastomeerid kleepuvad teineteisega tihedalt kokku, moodustades kompaktse struktuuri. Selles staadiumis olevat sügooti nimetatakse moorulaks. Moorula – kobarloode, millele on iseloomulik õõnsuse puudumine. Moorula staadiumile järgneb blastotsüsti staadium, mille lõpuks on välja arenenud põisloote ehk blastotsüsti sisemine õõnsus ehk blastotsööl. Kuni moorula staadiumini on kõik rakud totipotentsed. Kompakteerumise järgselt tekib kaks erinevat rakkude populatsiooni – trofoblast ehk trofektoderm ja embrüoblast ehk sisemine rakkude mass (ICM). Sisemine rakkude mass areneb edasi embrüoks. Trofoblasti rakud sekreteerivad vedelikku, mis põhjustab blastotsööli teket. Lisaks sellele on trofoblast oluline embrüo implantatsiooniks emakasse ning sellest arenevad lootevälised organid ja koed. Kogu lõigustumise protsess leiab aset munajuhas ning lõigustumise ajal liigub sügoot munajuhas olevate ripsrakkude ripsmete võnkelise liikumise mõjul emakavalendiku poole, kus toimub implantatsioon. Enne implanteerumist toimub blastotsüsti vabanemine glükoproteiinidest koosnevast zona pellucida’st ehk rebukestast. Kinnitumist emaka endomeetriumisse nimetatakse implantatsiooniks ning seda võimaldavad trofektodermi poolt toodetud proteaasid.[1]
Meroblastiline munarakkude lõigustumine
[muuda | muuda lähteteksti]Meroblastilise lõigustumise korral on tegu osalise lõigustumisega. Lõigustub rebuvaene osa, reburohke osa ei lõigustu. Osaline lõigustumine jaguneb diskoidaalseks ehk kettaliseks ja superfitsiaalseks ehk pindmiseks.
Partsiaalne diskoidne lõigustumine
[muuda | muuda lähteteksti]Partsiaalse diskoidaalse lõigustumise korral lõigustub ainult sügoodi rebuvaene osa. Rebuvaese osa lõigustumise järgselt kujuneb iduketas ehk blastodisk. Rakuvälisest rebust on blastodiski keskosa eraldatud subgerminaalse õõnega. Kölleri sirp – epiblasti rakud, mis paiknevad iduketta tagumises servas. Tulevasele iduketta bilateraalsümmeetriale pannakse alus juba munajuhas. Munajuhas toimub muna pöörlev liikumine. Pöörlemise tõttu liigub tsütoplasma iduketta ülaserva. Kölleri sirp moodustub lõigustumise käigus iduketta ülaservas paiknevast tsütoplasmast. Kölleri sirp määrab tulevase organismi posterioorse suuna.
Partsiaalne superfitsiaalne lõigustumine
[muuda | muuda lähteteksti]Partsiaalse superfitsiaalse lõigustumise puhul jaguneb algul ainult sügoodi tuum. Esimesena toimuvad tuuma mitootilised jagunemised. Sellele järgneb tuumade liikumine raku perifeeriasse, kus moodustatakse ilma rakkude vaheseinteta süntsütsiaalne blastoderm. Energiidid – tuumad (millel on ümber oma makrotorukeste ja mikrofilamentide rikas tsütoplasma) ja nendega seotud tsütoplasmasaared. Tuuma/tsütoplasma suhtest sõltub asünkroonsele lõigustumisele üleminek ja blastomeeride moodustumine. Kõikidel putukatel ei esine superfitsiaalset lõigustumist. On ka selliseid putukaid, kellel esineb totaalne lõigustumine. Sellisteks on näiteks parasiitsed putukad.[1]
Viited
[muuda | muuda lähteteksti]- ↑ 1,0 1,1 1,2 1,3 1,4 1,5 Jüri Kärner. Sissejuhatus arengubioloogiasse, Tartu: Tartu Ülikooli Kirjastus, 1997.
- ↑ Forgács, G. & Newman, Stuart A (2005). "Biological physics of the developing embryo". Cambridge University Press. Vaadatud 04.11.2011.
{{netiviide}}: CS1 hooldus: mitu nime: autorite loend (link)